- Bacterie
-
Bacteria
Bactérie
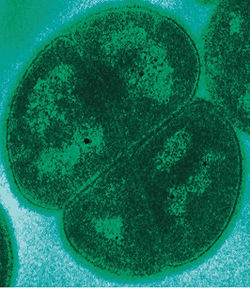
Une bactérie Deinococcus radiodurans. Domaine Prokaryota Règne Bacteria
— auteur incomplet —, date à préciserDivisions de rang inférieur Selon NCBI(en) : - Acidobacteria
- Actinobacteria
- Aquificae
- Bacterioidetes
- Chlamydiae
- Chlorobi
- Chloroflexi
- Chrysiogenetes
- Cyanobacteria
- Deferribacteres
- Deinococcus-Thermus
- Dictyoglomi
- Fibrobacteres
- Firmicutes
- Fusobacteria
- Gemmatimonadetes
- Nitrospira
- Planctomycetes
- Proteobacteria
- Spirochaetes
- Thermodesulfobacteria
- Thermomicrobia
- Thermotogae
- Verrucomicrobia
Bacteria Incertae sedis
Classification phylogénétique Position :
 Retrouvez ce taxon sur Wikispecies
Retrouvez ce taxon sur Wikispecies
D'autres documents multimédia
sont disponibles sur CommonsParcourez la microbiologie sur Wikipédia : 





Les bactéries (Bacteria) sont des organismes vivants unicellulaires procaryotes (caractérisées par une absence de noyau et d'organites). La plupart des bactéries possèdent une paroi cellulaire glucidique, le peptidoglycane. Les bactéries mesurent quelques micromètres de long et peuvent présenter différentes formes : des formes sphériques (coques), des formes allongées ou en bâtonnets (bacilles), des formes plus ou moins spiralées. L’étude des bactéries est la bactériologie, une branche de la microbiologie.
Les bactéries sont ubiquitaires et sont présentes dans tous les types de biotopes rencontrés sur Terre. Elles peuvent être isolées du sol, des eaux douces, marines ou saumâtres, de l’air, des profondeurs océaniques, des déchets radioactifs[1], de la croûte terrestre, sur la peau et dans l’intestin des animaux. Il y a environ 40 millions de cellules bactériennes dans un gramme de sol et 1 million de cellules bactériennes dans un millilitre d’eau douce. On estime qu'il y aurait (à un instant donné) quatre à six quintillions (4 ×1030 à 6×1030), soit entre 400 et 600 milliards de milliards de milliards de bactéries dans le monde[2], représentant une grande partie de la biomasse du monde[2]. Cependant, un grand nombre de ces bactéries ne sont pas encore caractérisées car non cultivables en laboratoire[3]. Les bactéries ont une importance considérable dans les cycles biogéochimiques comme le cycle du carbone et la fixation de l’azote de l’atmosphère.
Chez l'Homme, il a été calculé que 1012 bactéries colonisent la peau, 1010 bactéries colonisent la bouche et 1014 bactéries habitent dans l'intestin, ce qui fait qu'il y a dix fois plus de cellules bactériennes que de cellules humaines dans le corps humain[4]. La plupart de ces bactéries sont inoffensives ou bénéfiques pour l’organisme. Il existe cependant de nombreuses espèces pathogènes à l'origine de beaucoup de maladies infectieuses comme le choléra, la syphilis, la peste, l’anthrax, la tuberculose. Le plus souvent, les maladies bactériennes mortelles sont les infections respiratoires, la tuberculose à elle seule tue environ 2 millions de personnes par an, principalement en Afrique subsaharienne[5]. Des bactéries peuvent entraîner des troubles respiratoires ou intestinaux alors que d’autres peuvent être responsables de l’infection d'une blessure. Les infections bactériennes peuvent être traitées grâce aux antibiotiques, qui le plus souvent inhibent une de leurs fonctions vitales (par exemple, la pénicilline bloque la synthèse de la paroi cellulaire).
Les bactéries peuvent être très utiles à l’Homme lors des processus de traitement des eaux usées, dans l’agroalimentaire lors de la fabrication des yaourts ou du fromage et dans la production industrielle de nombreux composés chimiques[6].
Histoire
Les bactéries étant microscopiques, elles ne sont donc visibles qu'avec un microscope. Antoine van Leeuwenhoek fut le premier à observer des bactéries, grâce à un microscope de sa fabrication, en 1668[7]. Il les appela « animalcules » et publia ses observations dans une série de lettres qu'il envoya à la Royal Society[8],[9],[10].
Le mot « bactérie » apparaît pour la première fois avec le microbiologiste allemand Christian Gottfried Ehrenberg en 1828[11]. Ce mot dérive du grec βακτηριον, qui signifie « bâtonnet ».
Au XIXe siècle, les travaux de Louis Pasteur ont révolutionné la bactériologie. Il démontra en 1859 que les processus de fermentation sont causés par des micro-organismes et que cette croissance n’était pas due à la génération spontanée. Il démontra aussi le rôle des micro-organismes comme agents infectieux[12]. Pasteur conçut également des milieux de culture, des procédés de destruction des micro-organismes comme l’autoclave et la pasteurisation.
Le médecin allemand Robert Koch et ses collaborateurs mirent au point les techniques de culture des bactéries sur milieu solide. Robert Koch est un des pionniers de la microbiologie médicale, il a travaillé sur le choléra, la maladie du charbon (anthrax) et la tuberculose. Il démontra de façon claire qu’une bactérie pouvait être l’agent responsable d’une maladie infectieuse et il proposa une série de postulats (les postulats de Koch, toujours utilisés aujourd'hui[13]) confirmant le rôle étiologique d’un micro-organisme dans une maladie. Il obtint le prix Nobel de médecine et de physiologie en 1905[14].
Bien qu'on savait au dix-neuvième siècle que les bactéries sont la cause de nombreuses maladies, il n'y avait pas d'antiseptique disponibles[15]. En 1910, Paul Ehrlich développa le premier antibiotique, par l'évolution des colorants sélectifs teintés Treponema pallidum—le spirochaete qui cause la syphilis—en composés qui tuent l'agent pathogène de façon sélective[16]. Ehrlich a reçu en 1908 le prix Nobel pour ces travaux sur l'immunologie, et pionnier de l'usage de colorant pour détecter et identifier les bactéries, son travail étant la base de la coloration de Gram et de la coloration de Ziehl-Neelsen[17].
Les microbiologistes Martinus Beijerinck et Sergei Winogradsky initièrent les premiers travaux de microbiologie de l’environnement et d’écologie microbienne en étudiant les communautés microbiennes du sol et de l’eau et les relations entre ces micro-organismes.
Si les bactéries étaient connues au XIXe siècle, il n’existait pas encore de traitement antibactérien. En 1909, Paul Ehrlich mit au point un traitement contre la syphilis avant l’utilisation de la pénicilline en thérapeutique suggérée par Ernest Duchesne en 1897 et étudiée par Alexander Fleming en 1929.
En 1977, Carl Woese grâce à ses travaux de phylogénie moléculaire divisa les procaryotes en deux groupes : les Bacteria et les Archaea[18].
Structure cellulaire
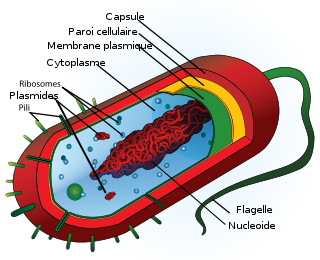 Schéma de la structure cellulaire d’une cellule bactérienne typique.
Schéma de la structure cellulaire d’une cellule bactérienne typique.
En tant que procaryote (organisme sans noyau), les bactéries sont des cellules relativement simples, caractérisées par une absence de noyau et d’organites comme les mitochondries et les chloroplastes, elles n'ont pas non plus de réticulum endoplasmique ou d'appareil de golgi[19].
Une caractéristique importante des bactéries est la paroi cellulaire. Les bactéries peuvent être divisées en deux groupes (Gram négatif et Gram positif) basé sur la différence de la structure et de la composition chimique de la paroi cellulaire mise en évidence grâce à la coloration de Gram. Les bactéries à coloration de Gram positif possèdent une paroi cellulaire contenant un peptidoglycane (ou muréine) épais et des acides teichoïques alors que bactéries à coloration de Gram négatif présentent un peptidoglycane fin localisé dans le périplasme entre la membrane cytoplasmique et une membrane cellulaire externe. La paroi donne à la bactérie sa forme et la protège contre l’éclatement sous l’effet de la très forte pression osmotique du cytoplasme. Le peptidoglycane assure la rigidité de la paroi. Il existe toutefois des bactéries sans paroi : ce sont les mycoplasmes.
Intracellulaire
Les bactéries étaient vues comme de simples sacs de cytoplasme, mais de nombreux niveaux de complexité structurelle ont été découverts depuis, comme la découverte du cytosquelette procaryote[20],[21], et la localisation spécifique de protéines dans le cytoplasme bactérien[22]. Ces compartiments subcellulaires ont été nommés « hyperstructure bactériennes » (« bacterial hyperstructures » en anglais)[23].
Les bactéries possèdent un chromosome sous forme de filament d’ADN, support de l’hérédité. Le chromosome bactérien est en général circulaire. En plus de cet ADN génomique, les cellules bactériennes contiennent souvent des molécules d’ADN circulaire extra-chromosomiques appelées plasmides. Les cellules contiennent aussi de nombreux ribosomes permettant la synthèse protéique grâce au mécanisme de la traduction. Le cytoplasme des procaryotes contient souvent des substances intracellulaires de réserve qui sont des stocks de nutriments sous forme de glycogène, amidon ou poly-b-hydroxybutyrate (PBH). Certaines espèces de bactéries aquatiques possèdent des vésicules à gaz qui assurent la flottabilité des cellules. D’autres espèces, les bactéries magnétotactiques, ont la particularité de présenter un magnétosome.
Extracellulaire
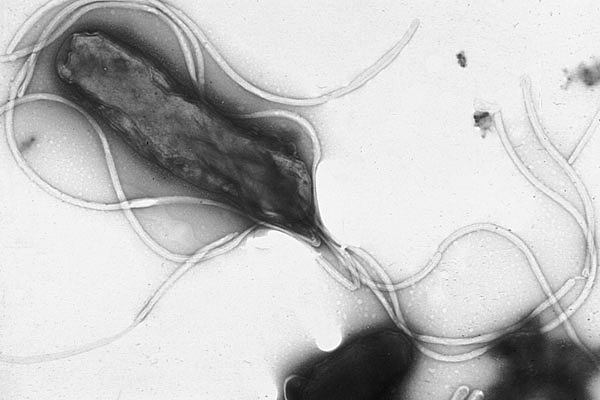
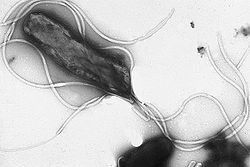 Helicobacter pylori micragraphie électronique montrant de nombreux flagelles à la surface de la cellule.
Helicobacter pylori micragraphie électronique montrant de nombreux flagelles à la surface de la cellule.Beaucoup de bactéries possèdent des structures extra-cellulaires comme des flagelles utilisés pour la mobilité des cellules, et des fimbriae permettant l’attachement ou le phénomène de conjugaison. Les bactéries hétérotrophes peuvent utiliser leurs flagelles pour se diriger vers des zones riches en substances organiques (nutriments) grâce au phénomène appelé chimiotactisme.
Quelques bactéries peuvent fabriquer de fines couches externes à la paroi cellulaire, généralement constituées de polysaccharides (des sucres). Quand la couche est compacte, on parle de capsule. Les capsules constituent par exemple une barrière de protection de la cellule contre l’environnement externe et aussi contre la phagocytose. Elle facilite aussi l’attachement aux surfaces et la formation de biofilms. Klebsiella, Bacillus anthracis, Streptococcus pneumoniae sont des exemples de bactéries capsulées. Quand la couche est diffuse, on parle de couche mucoïde. Quand la couche est plus épaisse, on parle de glycocalyx. Le glycocalyx permet aux bactéries d’adhérer à un support.
Certaines bactéries qualifiées de bactéries engainées produisent une couche externe dense et rigide : la gaine. Ce phénomène est courant chez les bactéries de l’eau qui forment des chaînes filamenteuses (Sphaerotilus natans par exemple). La gaine protège les cellules contre les turbulences de l’eau. Les bactéries du groupe Cytophaga – Flavobacterium produisent une couche muqueuse qui leur permet de rester en contact étroit avec un milieu solide. D'autres bactéries comme les Spirillum peuvent s’envelopper d’une couche protéique appelée la couche S.Endospores
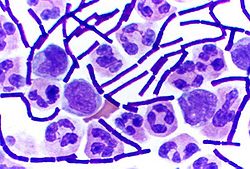
Quelques bactéries, Gram positif, comme Bacillus, Clostridium, Sporohalobacter, Anaerobacter et Heliobacterium peuvent fabriquer des endospores leur permettant de résister à certaines conditions de stress environnemental ou chimique[24]. La formation d'un endospore n'est pas un processus de reproduction. Les Anaerobacter peuvent faire jusqu'à sept endospores en une seule cellule[25]. Les bactéries à endospores ont une zone centrale de cytoplasme contenant l'ADN et ribosomes entouré par une couche du cortex et protégé par un manteau imperméable et rigide.
Les bactéries à endospores peuvent survivre dans des conditions physiques et chimiques extrèmes, tels que des niveaux élevés de rayonnement UV, les rayons gamma, les détergents, les désinfectants, une forte chaleur ou pression et à la dessiccation[26]. Ces organismes pourraient rester viable durant des millions d'années[27],[28]. Les endospores peuvent même permettre aux bactéries de survivre à l'exposition au vide et au rayonnement dans l'espace[29].
Les bactéries à endospore peuvent également causer des maladies: par exemple, la maladie du charbon peut être contractée par l'inhalation d'endospores Bacillus anthracis, et la contamination des plaies profondes avec la perforation par Clostridium tetani responsable du tétanos[30].
Croissance et reproduction
La division cellulaire
Deux cellules identiques sont produites à partir d’une cellule mère. La croissance cellulaire se manifeste par un accroissement du volume cellulaire, suivi de la synthèse d’un septum transversal au milieu de la cellule, aboutissant à la séparation des deux cellules filles. La division bactérienne est précédée par la duplication du chromosome bactérien grâce à la réplication de l’ADN.
Quelques bactéries présentent des structures reproductives plus complexes mais toujours de manière asexuée, facilitant la dispersion : Myxococcus élabore des fructifications, tandis que Streptomyces forme des hyphes aériens.
Quand elles se trouvent dans un milieu propice les bactéries peuvent se multiplier à une allure vertigineuse. Une population de bactérie peut doubler toutes les 20 minutes en fonction de : la disponibilité en nutriments, la présence de bactéries concurrentes, la présence de prédateurs (par exemple des paramécies), la présence de bactériophages, la présence d’antibiotiques (inhibant par exemple la synthèse de la paroi bactérienne, entraînant donc leur mort) produits par des champignons ou des actinomycètes (bactéries filamenteuses).
Croissance et culture des bactéries
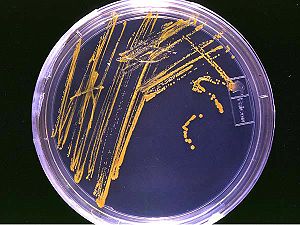 Colonies bactériennes sur milieu solide gélosé en boîte de Pétri.
Colonies bactériennes sur milieu solide gélosé en boîte de Pétri. Dans la nature, depuis des milliards d'années, les biofilms et concrétions bactériennes contribuent au cycle de nombreux éléments, à la formation de « filons » riches en métaux (par bioconcentration), ainsi qu'à la formation et dégradation des roches.
Dans la nature, depuis des milliards d'années, les biofilms et concrétions bactériennes contribuent au cycle de nombreux éléments, à la formation de « filons » riches en métaux (par bioconcentration), ainsi qu'à la formation et dégradation des roches.Au laboratoire, les bactéries peuvent être cultivées en milieu de culture liquide ou en milieu solide. Le milieu de culture doit apporter les éléments nutritifs ou nutriments élémentaires à la bactérie. Les milieux de culture gélosés solides sont utilisés pour isoler des cultures pures de cellules bactériennes. Dans le cas des bactéries se divisant rapidement, une cellule bactérienne dispersée sur un milieu gélosé va se multiplier et, au bout de 24 à 48 heures, devenir un amas de bactéries, appelé une colonie bactérienne, visible à l’œil nu.
Le temps de génération est le temps nécessaire à une bactérie pour se diviser. Le temps de génération correspond donc au temps nécessaire pour qu’une population de cellules double en nombre. Ce temps est très variable selon les espèces de bactéries et les conditions environnementales. Au laboratoire, dans des conditions idéales, il est par exemple de 20 minutes pour Escherichia coli, 100 minutes pour Lactobacillus acidophilus, 1 000 minutes pour Mycobacterium tuberculosis.
La croissance d’une population bactérienne dans un milieu de culture liquide non renouvelé, peut être observée dans le temps. Les cellules se divisent, et leur nombre augmente avec le temps. Si on relève le nombre de bactéries à différents intervalles au cours de la croissance, on obtient une courbe de croissance. Elle présente quatre phases principales :
- La phase de latence correspond à une période d’adaptation de la bactérie au milieu
- Au cours de la phase de croissance exponentielle, les bactéries se développent de façon maximale avec un taux de croissance maximal et constant
- Après une phase transitoire de ralentissement, le nombre de bactéries n’évolue plus : c’est la phase stationnaire. Les divisions bactériennes qui se font encore sont compensées par la mort de bactéries
- La dernière phase est la phase de mortalité ou de déclin. Les bactéries ne se divisent plus, elles meurent et peuvent être lysées. Le milieu de culture n’apporte plus les conditions nécessaires au développement des bactéries. On observe une courbe de décroissance exponentielle progressive.
Paramètres influant sur la croissance microbienne
Certaines conditions environnementales (paramètres physico-chimiques) influencent la croissance des micro-organismes. Parmi celles-ci figurent le pH (acidité et alcalinité), la température, la présence d’O2, de CO2, la disponibilité de l’eau.
La plupart des micro-organismes tolèrent une gamme de pH permettant la croissance. Le pH optimal de croissance de beaucoup de bactéries est proche de la neutralité (pH 7). Les micro-organismes acidophiles se développent à des pH acides, alors que les micro-organismes alcalinophiles se développent à des pH basiques.
De même, les bactéries peuvent être distinguées selon leur aptitude à croître en fonction de la température. Les mésophiles se développent généralement à des températures comprises entre 20 et 45 °C. Les psychrophiles possèdent des températures optimales de croissance inférieures à 15 °C, alors que les bactéries thermophiles croissent de façon optimale à des températures comprises entre 45 et 70 °C. Les micro-organismes ayant des températures optimales de croissance supérieures à 70 °C sont qualifiés d’hyperthermophiles.Génétique
Matériel génétique
La plupart des bactéries possèdent un unique chromosome circulaire. Il existe toutefois de rares exemples de bactéries, comme Rhodobacter sphaeroides possédant deux chromosomes. Les bactéries du genre Borrelia ont la particularité d'avoir un génome linéaire et segmenté, ce qui est exceptionnel chez les procaryotes. La taille du génome peut être très variable selon les espèces de bactéries étudiées. Le génome de la souche de Escherichia coli séquencé en 1997 est constitué de 4,6 Mpb (4 600 000 paires de bases), il code 4 200 protéines. Le génome d’une autre souche de E. coli séquencé en 2001 comprend 5,5 Mpb codant 5 400 protéines.
Certaines bactéries présentent un tout petit génome, comme la bactérie parasite Mycoplasma genitalium avec un génome de 580 000 paires de bases et la bactérie endosymbiotique d’insecte, Candidatus Carsonella ruddii avec un génome de seulement 160 000 paires de bases[31]. Au contraire, la bactérie du sol Sorangium cellulosum possède un génome constitué de 12 200 000 paires de bases[32]. Chose peu commune, les Spirochètes ainsi que des Streptomyces présentent la particularité d’avoir un chromosome linéaire[33].Les bactéries contiennent également souvent un ou plusieurs plasmides, qui sont des molécules d’ADN extra-chromosomique. Ces plasmides peuvent conférer certains avantages aux bactéries, comme la résistance à des antibiotiques ou des facteurs de virulence. Les plasmides sont généralement des ADN double brin circulaire. Ils se répliquent indépendamment du chromosome. Le chromosome bactérien peut d’autre part intégrer de l’ADN de virus bactérien (bactériophage). Ces bactériophages peuvent contribuer au phénotype de l’hôte[34]. Par exemple, les bactéries Clostridium botulinum et Escherichia coli O157:H7 synthétisent une toxine codée par un gène qui provient d’un phage qui s’est intégré au génome de ces bactéries au cours de l’évolution[35].
Variation génétique
Les bactéries sont des organismes asexués, après la division bactérienne, les cellules filles héritent d’une copie identique du génome de leur parent. Cependant, toutes les bactéries sont capables d’évoluer par modification de leur matériel génétique causé par des recombinaisons génétiques ou des mutations. Les mutations (changement ponctuel aléatoire de l'information génétique d'une cellule) proviennent d’erreur durant la réplication de l’ADN ou de l’exposition à des agents mutagènes. le taux de mutation varie grandement selon les espèces ou les souches bactériennes[36].
Quelques bactéries peuvent également transférer du matériel génétique entre les cellules. Il existe 3 mécanismes de transfert de gènes entre les cellules : la transformation, la transduction, et la conjugaison.
Au cours de la transformation, c’est un plasmide qui est transféré dans la cellule bactérienne, alors qu’au cours de la transduction, le transfert d’ADN a lieu par l’intermédiaire d’un bactériophage. Au cours de la conjugaison, deux bactéries peuvent se rapprocher, grâce à des structures spéciales, les pili, et il y a alors un transfert d’ADN d’une bactérie à une autre. L’ADN étranger peut être intégré dans le génome et être transmis aux générations suivantes. Cette acquisition de gènes, provenant d’une bactérie ou de l’environnement, est appelé transfert horizontal de gènes (HGT pour horizontal gene transfer)[37]. Le transfert de gènes est particulièrement important dans les mécanismes de résistance aux antibiotiques[38].Morphologie et association des bactéries
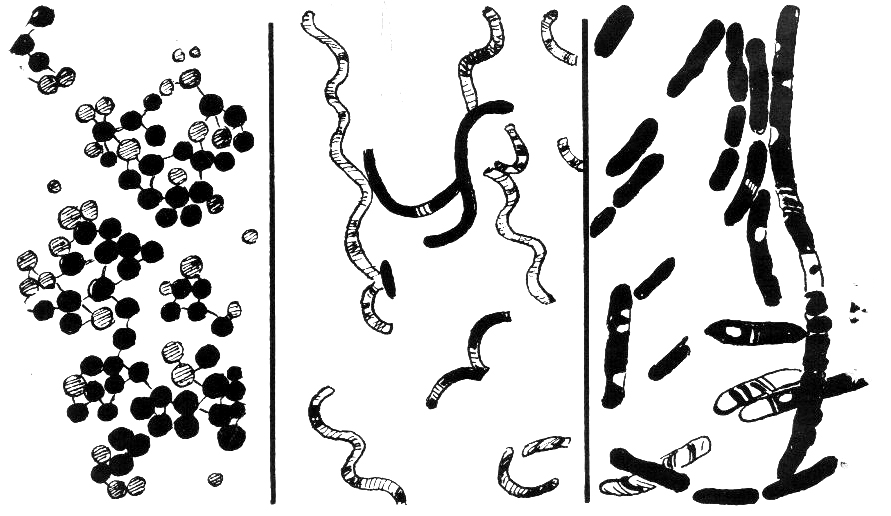
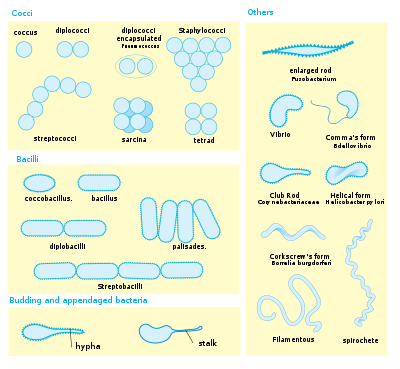 Les bactéries présentent une grande diversité de morphologies et d'arrangements cellulaires.
Les bactéries présentent une grande diversité de morphologies et d'arrangements cellulaires.Les bactéries présentent une grande diversité de tailles et de formes. Les cellules bactériennes typiques ont une taille comprise entre 0,5 et 5 µm de longueur, cependant, quelques espèces comme Thiomargarita namibiensis et Epulopiscium fishelsoni peuvent mesurer jusqu’à 500 µm (0,5 mm) de long et être visibles à l’œil nu[39]. Parmi les plus petites bactéries, les Mycoplasmes mesurent 0,3 µm, soit une taille comparable à certains gros virus[40].
La plupart des bactéries sont soit sphériques, appelées coques (pl. cocci, du grecque kókkos, grain), ou soit en forme de bâtonnets, appelés bacilles (pl. baccili, du Latin baculus, bâton). Il existe aussi des formes intermédiaires : les cocobacilles. Quelques bactéries en forme de bâtonnets sont légèrement incurvées comme les Vibrio. D’autres bactéries sont hélicoïdales. Ce sont des spirilles si la forme est invariable et rigide, des spirochètes si l’organisme est flexible et peut changer de forme. La grande diversité de formes est déterminée par la paroi cellulaire et le cytosquelette. Les différentes formes de bactéries peuvent influencer leur capacité d’acquérir des nutriments, de s’attacher aux surfaces, de nager dans un liquide et d’échapper à la prédation.
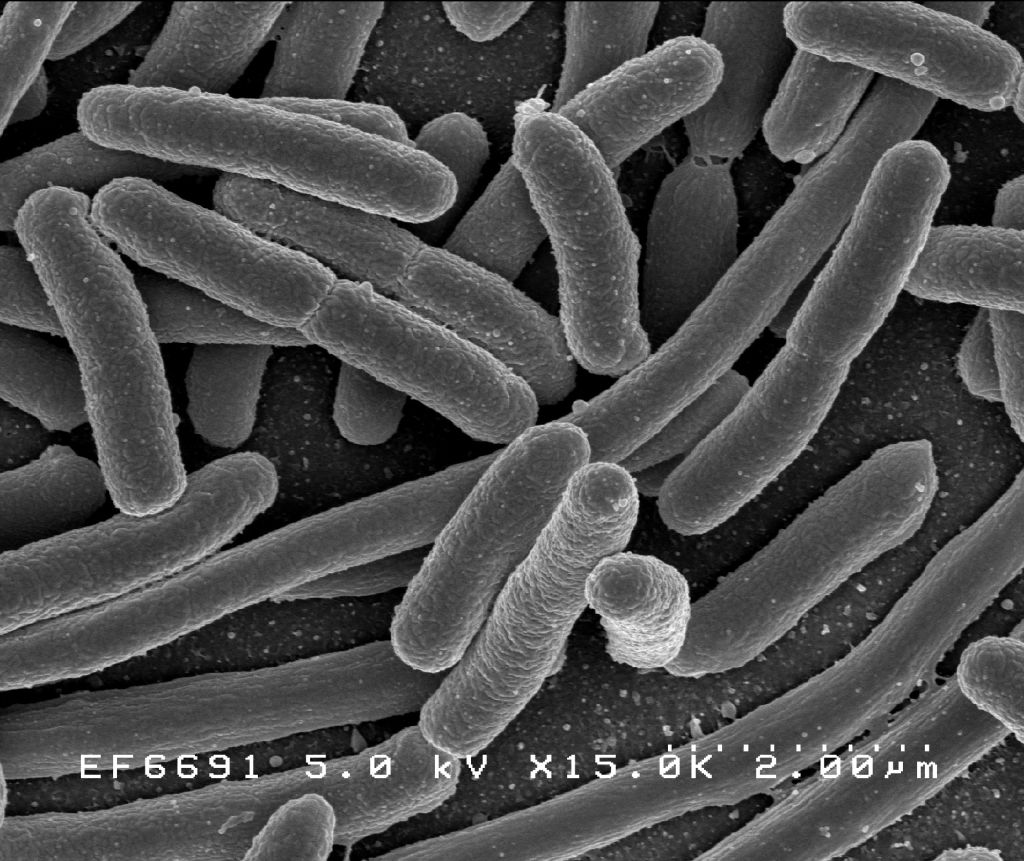
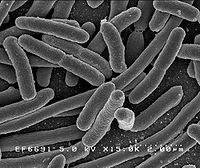 Escherichia Coli observée au microscope électronique.
Escherichia Coli observée au microscope électronique.Beaucoup d’espèces bactériennes peuvent être observées sous forme unicellulaire isolée alors que d’autres espèces sont associées en paires (diploïdes) comme les Neisseria ou en en chaînette, caractéristique des Streptocoques. Dans ces cas, les coques se divisent selon un axe unique et les cellules restent liées après la division. Certains coques se divisent selon un axe perpendiculaire et s’agencent de façon régulière pour former des feuillets. D’autres se divisent de façon désordonnée et forment des amas comme les membres du genre Staphylococcus qui présentent un regroupement caractéristique en grappe de raisins. D’autres bactéries peuvent s’élonger et former des filaments composés de plusieurs cellules comme les Actinomycetes. D’autres organismes comme les cyanobactéries forment des chaînes appelées trichomes. Dans ce cas, les cellules sont en relation étroite et les échanges physiologiques sont favorisés.
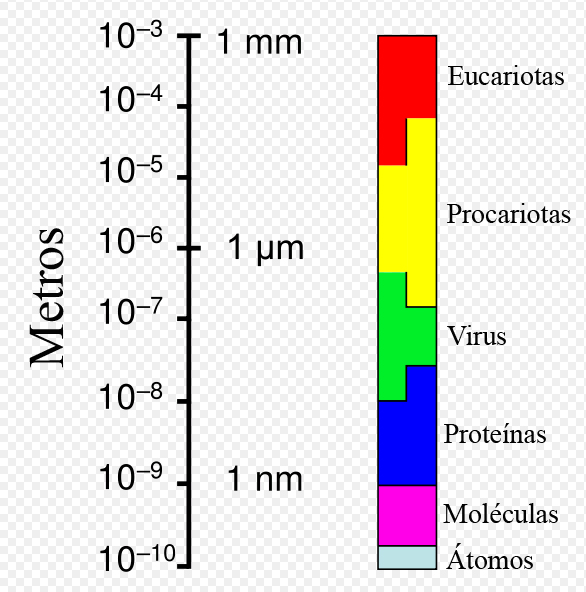
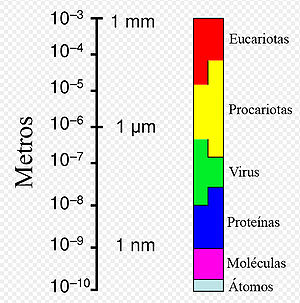 Gamme de tailles montrant les cellules procaryotes en relation avec d'autres organismes et biomolécules.
Gamme de tailles montrant les cellules procaryotes en relation avec d'autres organismes et biomolécules.En dépit de leur apparente simplicité, les bactéries peuvent aussi former des associations complexes. Elles peuvent s’attacher aux surfaces et former des agrégations appelées biofilms. Les bactéries présentes dans le biofilm peuvent présenter un arrangement complexe de cellules et de composants extra-cellulaires, formant des structures secondaires comme des microcolonies, dans lesquelles se forme un réseau de canal facilitant la diffusion des nutriments.
Au sein des biofilms des relations s'établissent entre bactéries, conduisant à une réponse cellulaire intégrée. Les molécules de la communication cellulaire ou « lang » sont soit des Homosérine lactones pour les bactéries à Gram négatif, soit des peptides courts pour les bactéries à Gram positif. De plus au sein de biofilms établis, les caractéristiques physico-chimiques (pH, oxygénation, métabolites) sont néfastes au bon développement bactérien et constituent des conditions stressantes. Les bactéries mettent en place des réponses de stress qui sont autant d'adaptation à ces conditions défavorables. En général les réponses de stress rendent les bactéries plus résistantes à toute forme de destruction par des agents mécaniques ou des molécules biocides.
Mobilité des bactéries
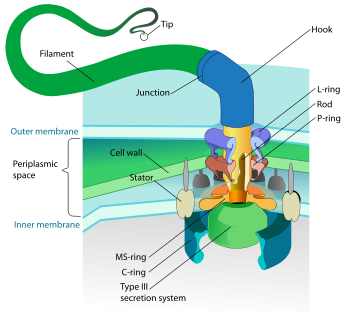 Flagelle d'une bactérie Gram-. La base entraine une rotation du crochet et des filaments.
Flagelle d'une bactérie Gram-. La base entraine une rotation du crochet et des filaments.Certaines bactéries sont mobiles et peuvent se déplacer grâce à un ou plusieurs flagelles, d’autres bactéries peuvent se déplacer par glissement.
Les flagelles des bactéries sont de longs appendices protéiques flexibles. Leur nombre et leur position peuvent différer selon les espèces de bactéries. La flagellation (ou ciliature) polaire monotriche correspond à la présence d’un seul flagelle à un pôle de la bactérie (exemple des Vibrio). La flagellation polaire lophotriche correspond à la présence de plusieurs flagelles au pôle de la bactérie (Pseudomonas par exemple). D’autres bactéries comme Escherichia coli produisent des flagelles sur toute la surface cellulaire et possèdent donc une flagellation péritriche.
Beaucoup de bactéries (comme Escherichia coli) ont deux modes distincts de circulation : mouvement vers l'avant (natation) et mouvement de rotation ou mouvement de « roulis » également et appelé « lang » en anglais. Le tumbling leur permet de se réorienter et leur fait faire un mouvement en trois dimensions, évoluant vers un mouvement de marche au hasard[41]. Les flagelles d'un groupe de bactéries, les spirochaetes, se trouvent entre deux membranes dans l'espace périplasmique[42].
Le filament du flagelle est constitué d’une protéine, la flagelline. Le type de rotation du flagelle peut déterminer le type de mouvement de la bactérie.
Les bactéries mobiles peuvent réagir à des stimuli, être attirées par des substances nutritives comme les sucres, les acides aminés, l’oxygène, ou être repoussées par des substances nuisibles. il s'agit de la chimiotaxie, phototaxis et magnetotaxis[43],[44]. Ce comportement est appelé le chimiotactisme. Des chimiorécepteurs de nature protéique sont présents au niveau de la membrane plasmique et du périplasme des bactéries et peuvent détecter différentes substances attractives ou nocives.
Les ions de cuivre bloquent la rotation des flagelles. Pour le faire repartir, on recourt à l’acide éthylènediaminetétraacétique, capable de capturer les ions et donc d'en libérer le flagelle.
Métabolisme
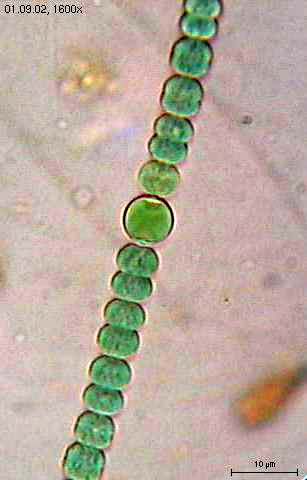
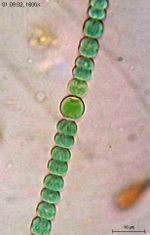 Une cyanobactérie : Anabaena sperica.
Une cyanobactérie : Anabaena sperica.Le métabolisme d’une cellule est l’ensemble des réactions chimiques qui se produisent au niveau de cette cellule. Pour réaliser ce processus, les bactéries, comme toutes les autres cellules, ont besoin d’énergie. L’ATP est la source d’énergie biochimique universelle. L’ATP est commune à toutes les formes de vies, mais les réactions d’oxydo-réduction impliquées dans sa synthèse sont très variées selon les organismes et notamment chez les bactéries. Les bactéries vivent dans pratiquement toutes les niches environnementales de la biosphère. Elles peuvent ainsi utiliser une très large variété de source de carbone et/ou d’énergie.
Les bactéries peuvent être classées selon leur type de métabolisme, en fonction des sources de carbone et d’énergie utilisés pour la croissance, les donneurs d’électrons et les accepteurs d’électrons.
L’énergie cellulaire des chimiotrophes est d’origine chimique alors que celle des phototrophes est d’origine lumineuse. La source de carbone des autotrophes est le CO2, tandis que des substrats organiques sont la source de carbone des hétérotrophes. Il est aussi possible de distinguer deux sources possibles de protons (H+) et d'électrons (e-) : les bactéries réduisant des composés minéraux sont des lithotrophes alors que celles réduisant des substances organiques sont des organotrophes.Les bactéries peuvent être divisées en quatre grands types nutritionnels en fonction de leurs sources de carbone et d’énergie :
- Les photoautotrophes utilisent la lumière comme source d’énergie et le CO2 comme source de carbone.
- Les photohétérotrophes se développent par photosynthèse. Ils assimilent le CO2 en présence d’un donneur d’électrons.
- Les chimioautotrophes utilisent des substrats inorganiques réduits pour l’assimilation réductrice du CO2 et comme source d’énergie.
- Les chimiohétérotrophes utilisent des substrats organiques comme source de carbone et d’énergie.
Chez les chimiohétérotrophes, les substrats sont dégradés en plus petites molécules pour donner des métabolites intermédiaires (pyruvate, acétylCoA…) qui sont eux-mêmes dégradés avec production de CO2, H2O et d’énergie. Ces réactions productrices d’énergie sont des réactions d’oxydation d’un substrat hydrogéné, avec libération de protons et d’électrons grâce à des déshydrogénases. Le transfert de protons et d’électrons à un accepteur final est réalisé par toute une série d’enzymes qui forment une chaîne de transport électronique. L’énergie ainsi produite est libérée par petites étapes dans le but d’être transférée dans des liaisons chimiques riches en énergie (ATP, NADH, NADPH). Suivant la nature de l’accepteur final d’électrons, on distingue les processus de la respiration et de la fermentation. La respiration peut être aérobie quand O2 est l’accepteur final de protons et d’électrons, ou anaérobie (respiration nitrate, et respiration fumarate par exemple). Dans tous les cas, l’accepteur final d’électrons doit être une molécule oxydée (O2, NO3−, SO2−).
Chez les organismes aérobies, l’oxygène est utilisé comme accepteur d’électrons. Chez les organismes anaérobies, d’autres composés inorganiques comme le nitrate, le sulfate ou le dioxyde de carbone sont utilisés comme accepteurs d’électrons. Ces organismes participent à des processus écologiques très importants lors de la dénitrification, la réduction des sulfates et l’acétogénèse. Ces processus sont aussi importants lors de réponses biologiques à la pollution, par exemple, les bactéries réduisant les sulfates sont responsables de la production de composés hautement toxiques à partir du mercure (méthyl et diméthylmercure) présent dans l’environnement. Les anaérobies (non respiratoires) utilisent la fermentation pour fournir de l’énergie à la croissance des bactéries. Au cours de la fermentation, un composé organique (le substrat ou la source d’énergie) est le donneur d’électrons tandis qu’un autre composé organique est l’accepteur d’électrons. Les principaux substrats utilisés lors de la fermentation sont des glucides, des acides aminés, des purines et des pyrimidines. Divers composés peuvent être relargués par les bactéries lors des fermentations. Par exemple, la fermentation alcoolique conduit à la formation d’éthanol et de CO2. Les bactéries anaérobies facultatives sont capables de modifier leur métabolisme entre la fermentation et différents accepteurs terminaux d’électrons, selon les conditions du milieu où elles se trouvent.Selon leur mode de vie, les bactéries peuvent être classées en différents groupes :
- Les aérobies strictes peuvent vivre uniquement en présence de dioxygène ou oxygène moléculaire (O2).
- Les aéro-anaérobies facultatives peuvent vivre en présence ou en absence de dioxygène.
- Les anaérobies ne peuvent vivre qu'en absence de dioxygène. Les aérotolérants sont des organismes anaérobies qui peuvent tout de même survivre en présence d’oxygène.
- les microaérophiles requièrent de l’oxygène pour survivre mais à une concentration faible.
Les bactéries lithotrophes peuvent utiliser des composés inorganiques comme source d’énergie. L’hydrogène, le monoxyde de carbone, l’ammoniac (NH3), les ions ferreux ainsi que d’autres ions métalliques réduits et quelques composés du soufre réduit. Le méthane peut être utilisé par les méthanotrophes comme source de carbone et d’électrons. Chez les phototrophes aérobie et les chimiolithotrophe, l’oxygène est utilisé comme accepteur terminal d’électrons, alors qu’en condition anaérobie, ce sont des composés inorganiques qui sont utilisés.
En plus de la fixation du CO2 lors de la photosynthèse, quelques bactéries peuvent fixer l’azote N2 (fixation de l’azote en utilisant une enzyme : la nitrogénase. Des bactéries aérobies, anaérobies et photosynthétiques sont capables de fixer l’azote. Les cyanobactéries qui fixent l’azote, possèdent des cellules spécialisées (les hétérocystes).
Bactéries et écosystème
Les bactéries, avec les autres micro-organismes participent pour une très large part à l’équilibre biologique existant à la surface de la Terre. Ils colonisent en effet tous les écosystèmes et sont à l’origine de transformations chimiques fondamentales lors des processus biogéochimiques responsables du cycle des éléments sur la planète.
Écosystème aquatique
Les eaux naturelles comme les eaux marines (océans) ou les eaux douces (lacs, mares, étangs, rivières…) sont des habitats microbiens très importants. Les matières organiques en solution et les minéraux dissous permettent le développement des bactéries. Les bactéries participent dans ces milieux à l’autoépuration des eaux. Elles sont aussi la proie des protozoaires. Les bactéries composant le plancton des milieux aquatiques sont appelées le bactérioplancton.
Bactérie du sol
Le sol est composé de matière minérale provenant de l’érosion des roches et de matière organique (l’humus) provenant de la décomposition partielle des végétaux. La flore microbienne y est très variée. Elle comprend des bactéries, des champignons, des protozoaires, des algues, des virus, mais les bactéries sont les représentants les plus importants quantitativement. On peut y retrouver tous les types de bactéries, des autotrophes, des hétérotrophes, des aérobies, des anaérobies, des mésophiles, des psychrophiles, des thermophiles. Tout comme les champignons, certaines bactéries sont capables de dégrader des substances insolubles d’origine végétale comme la cellulose, la lignine, de réduire les sulfates, d’oxyder le soufre, de fixer l’azote atmosphérique et de produire des nitrates. Les bactéries jouent un rôle dans le cycle des nutriments des sols, et sont notamment capables de fixer l’azote. Elles ont donc un rôle dans la fertilité des sols pour l’agriculture. Les bactéries abondent au niveau des racines des végétaux avec lesquels elles vivent en mutualisme.
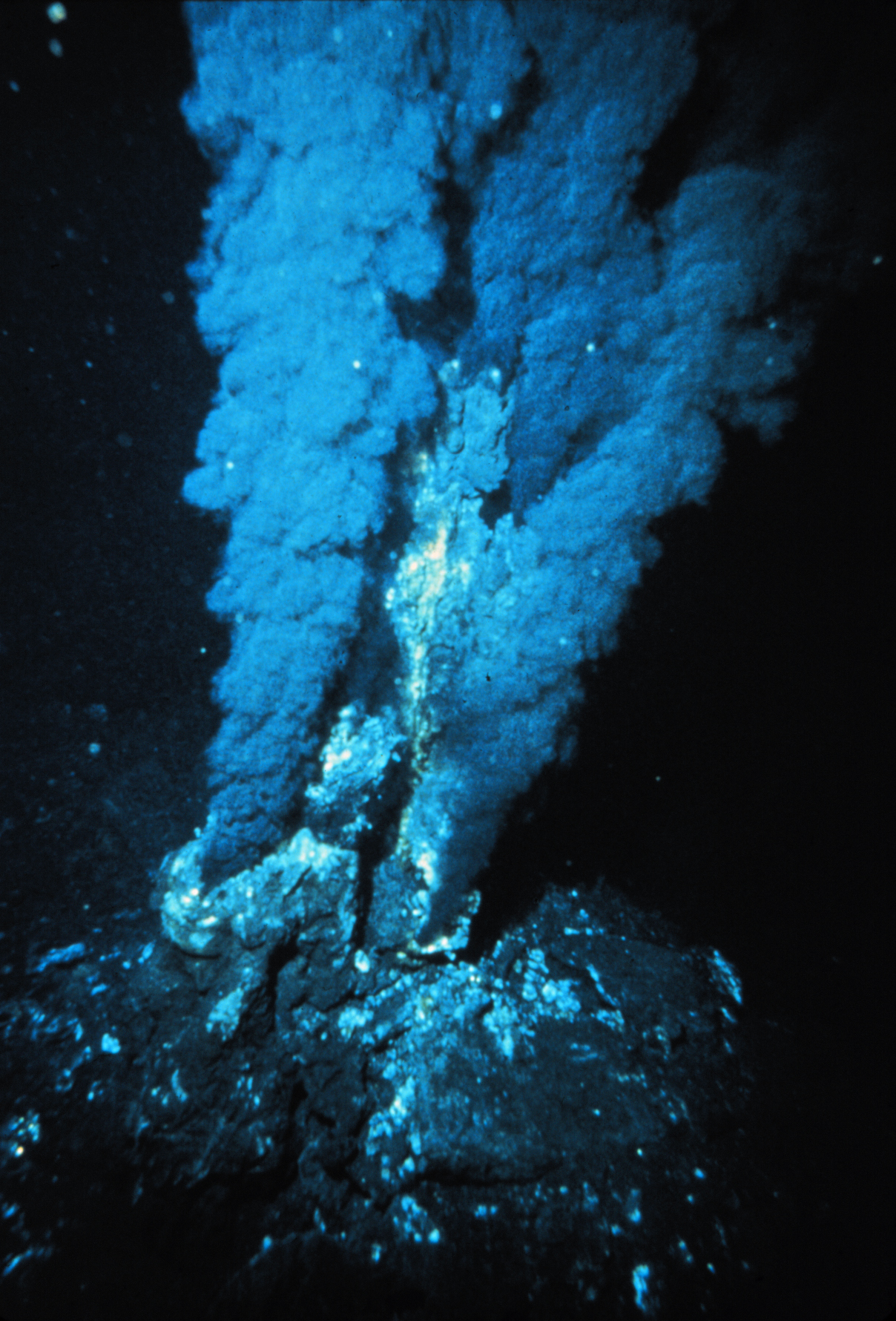
 Une cheminée hydrothermale
Une cheminée hydrothermaleÀ la différence des milieux aquatiques, l’eau n’est pas toujours disponible dans les sols. Les bactéries ont mis en place des stratégies pour s’adapter aux périodes sèches. Les Azotobacter produisent des cystes, les Clostridium et les Bacillus des endospores ou d’autres types de spores chez les Actinomycètes.
Environnements extrêmes
Les bactéries peuvent aussi être rencontrées dans des environnements plus extrêmes. Elles sont qualifiées d’extrémophiles. Des bactéries halophiles sont rencontrées dans des lacs salés, des bactéries psychrophiles sont isolées d’environnements froids comme des océans Arctique et Antarctique, des banquises. Des bactéries thermophiles sont isolées des sources chaudes ou des cheminées hydrothermales.
Interactions avec d’autres organismes
En dépit de leur apparente simplicité, les bactéries peuvent entretenir des associations complexes avec d’autres organismes. Ces associations peuvent être répertoriées en parasitisme, mutualisme et commensalisme. En raison de leurs petites tailles, les bactéries commensales sont ubiquitaires et sont rencontrées à la surface et à l’intérieur des plantes et des animaux.
Mutualistes
Dans le sol, les bactéries de la rhizosphère (couche de sol fixée aux racines des plantes) fixent l’azote et produisent des composés azotés utilisés par les plantes (exemple de la bactérie Azotobacter ou Frankia). En échange, la plante excrète au niveau des racines des sucres, des acides aminés et des vitamines qui stimulent la croissance des bactéries. D’autres bactéries comme Rhizobium sont associées aux plantes légumineuses au niveau de nodosités sur les racines.
Il existe de nombreuses relations symbiotiques ou mutualistes de bactéries avec des invertébrés. Par exemple, les animaux qui se développent à proximité des cheminées hydrothermales des fonds océaniques comme les vers tubicoles Riftia pachyptila, les moules Bathymodiolus ou la crevette Rimicaris exoculata vivent en symbiose avec des bactéries chimiolitho-autotrophes.
Buchnera est une bactérie endosymbiote des aphides (puceron). Elle vit à l'intérieur des cellules de l'insecte et lui fournit des acides aminés essentiels. La bactérie Wolbachia est hébergée dans les testicules ou les ovaires de certains insectes. Cette bactérie peut contrôler les capacités de reproduction de son hôte.
Des bactéries sont associées aux termites et lui apportent des sources d'azote et de carbone.Des bactéries colonisant la panse des herbivores permettent la digestion de la cellulose par ces animaux. La présence de bactéries dans l’intestin de l’Homme contribue à la digestion des aliments mais les bactéries fabriquent également des vitamines comme l’acide folique, la vitamine K et la biotine[45].
Des bactéries colonisent le jabot d'un oiseau folivore (consommateur de feuilles), le Hoazin (Opisthocomus hoazin). Ces bactéries permettent la digestion de la cellulose des feuilles, de la même manière que dans le rumen des ruminants.Des bactéries bioluminescentes comme Photobacterium sont souvent associées à des poissons ou des invertébrés marins. Ces bactéries sont hébérgées dans des organes spécifiques chez leurs hôtes et émettent une luminescence grâce à une protéine particulière : la luciférase. Cette luminescence est utilisée par l'animal lors de divers comportements comme la reproduction, l'attraction de proies ou la dissuasion de prédateurs.
Pathogènes
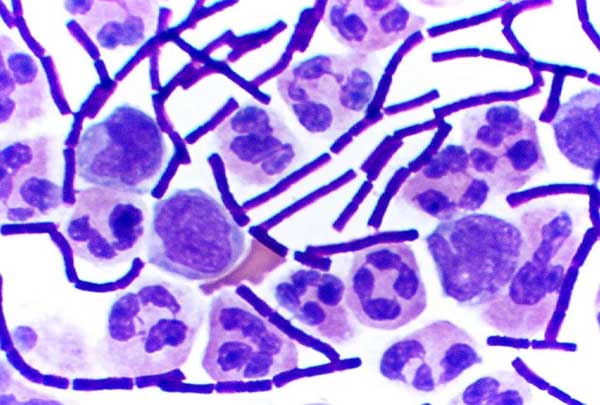
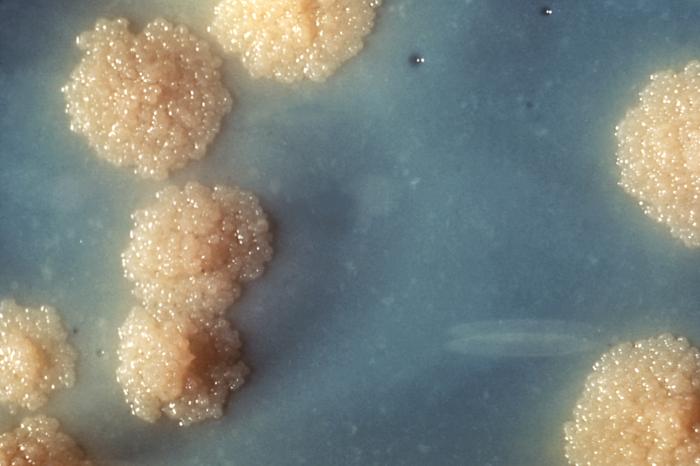
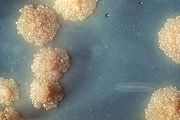 Culture de Mycobacterium tuberculosis.
Culture de Mycobacterium tuberculosis.Les bactéries pathogènes sont responsables de maladies humaines et causent des infections. Les organismes infectieux peuvent être distingués en trois types : les pathogènes obligatoires, accidentels ou opportunistes.
Un pathogène obligatoire ne peut survivre en dehors de son hôte. Parmi les bactéries pathogènes obligatoires, Corynebacterium diphtheriae entraîne la diphtérie, Treponema pallidum est l’agent de la syphilis, Mycobacterium tuberculosis provoque la tuberculose, Mycobacterium leprae la lèpre, Neisseria gonorrhoeae la gonorrhée. Les Rickettsia à l’origine du typhus sont des bactéries parasites intracellulaires.
Un pathogène accidentel présent dans la nature peut infecter l’Homme dans certaines conditions. Par exemple, Clostridium tetani provoque le tétanos en pénétrant dans une plaie. Vibrio cholerae entraîne le choléra suite à la consommation d’une eau contaminée.
Un pathogène opportuniste infecte des individus affaiblis ou atteints par une autre maladie. Des bactéries comme Pseudomonas aeruginosa, des espèces de la flore normale, comme des Staphylococcus de la flore cutanée, peuvent devenir des pathogènes opportunistes dans certaines conditions. On rencontre ce type d’infection surtout en milieu hospitalier.La capacité d’une bactérie à provoquer une maladie est son pouvoir pathogène. L’intensité du pouvoir pathogène est la virulence. L’aboutissement de la relation bactérie-hôte et l’évolution de la maladie dépendent du nombre de bactéries pathogènes présentes dans l’hôte, de la virulence de cette bactérie, des défenses de l’hôte et de son degré de résistance.
Pour déclencher une maladie, les bactéries infectieuses doivent d’abord pénétrer dans l’organisme et adhérer à un tissu. Des facteurs d’adhésion permettent la fixation des bactéries à une cellule. Le pouvoir invasif est la capacité de la bactérie à se répandre et à se multiplier dans les tissus de l’hôte. Les bactéries peuvent produire des substances lytiques lui permettant de se disséminer dans les tissus. Certaines bactéries présentent aussi un pouvoir toxinogène qui est la capacité de produire des toxines, substances chimiques portant préjudice à l’hôte. On peut distinguer les exotoxines libérées lors de la multiplication des bactéries et les endotoxines fixées dans la membrane des bactéries.Les bactéries pathogènes tentant d’envahir un hôte rencontrent toutefois de nombreux mécanismes de défense assurant à l’organisme une protection aux infections. Une bonne alimentation et une hygiène de vie correcte constituent une première protection. La peau, les muqueuses forment une première ligne de défense contre la pénétration d’organismes pathogènes. Les bactéries de la flore normale constituent aussi une barrière de protection. Lorsqu’un micro-organisme a pénétré ces premières lignes de défense, il rencontre des cellules spécialisées qui se mobilisent contre l’envahissement : ce sont les phagocytes. L’inflammation est une réaction défensive non spécifique. Un second système de défense très efficace est le système immunitaire spécifique, capable de reconnaître des antigènes portés ou sécrétés par les bactéries, et d’élaborer des anticorps et des cellules immunitaires spécifiques de ces antigènes.
Importance des bactéries dans l’industrie et les technologies
L’origine de la microbiologie industrielle date de l’époque préhistorique. Les premières civilisations ont utilisé sans le savoir des micro-organismes pour produire des boissons alcoolisées, du pain et du fromage.
Les bactéries comme Lactobacillus, Lactococcus ou Streptococcus, combinées aux levures et moisissures interviennent dans l’élaboration d’aliments fermentés comme les fromages, les yaourts, la bière, le vin, la sauce de soja, le vinaigre, la choucroute.
Les bactéries acétiques (Acetobacter, Gluconobacter) peuvent produire de l'acide acétique à partir de l'éthanol. Elles sont rencontrées dans les jus alcoolisés et sont utilisées dans la production du vinaigre. Elles sont également exploitées pour la production d'acide ascorbique (vitamine C) à partir du sorbitol transformée en sorbose.
La capacité des bactéries hétérotrophes à dégrader une large variété de composés organiques est exploitée dans des processus de traitement des déchets comme la bioremédiation ou le traitement des eaux usées. Des bactéries sont également utilisées dans les fosses septiques pour en assurer l'épuration. Des bactéries, capables de dégrader des hydrocarbures du pétrole, peuvent être utilisées lors du nettoyage d'une marée noire. Le processus de nettoyage de milieux pollués par des micro-organismes est la bioremédiation.
Des bactéries peuvent être utilisées pour récupérer des métaux d'intérêts économiques à partir de minerais. C'est la biolixiviation. L'activité de bactéries est ainsi exploitée pour la récupération du cuivre.
Des bactéries peuvent être utilisées à la place de pesticides en lutte biologique pour combattre des parasites des plantes. Par exemple, Bacillus thuringiensis produit une protéine Bt qui est toxique pour certains insectes. Cette toxine est utilisée en agriculture pour combattre des insectes qui se nourrissent de plantes.
En raison de leur capacité à se multiplier rapidement et de leur relative facilité à être manipulées, certaines bactéries comme Escherichia coli sont des outils très utilisés en biologie moléculaire, génétique et biochimie. Les scientifiques peuvent déterminer la fonction de gènes, d’enzymes ou identifier des voies métaboliques nécessaires à la compréhension fondamentale du vivant et permettant également de mettre en œuvre de nouvelles applications en biotechnologie.
De nombreuses enzymes utilisées dans divers processus industriels ont été isolées de micro-organismes. Les enzymes des détergents sont des protéases de certaines souches de Bacillus. Des amylases capables d’hydrolyser l’amidon sont très utilisées dans l’industrie alimentaire. La Taq polymérase utilisée dans les réactions de polymérisation en chaîne (PCR) pour l’amplification de l’ADN provient d’une bactérie thermophile Thermus aquaticus.
Les bactéries génétiquement modifiées sont très utilisées pour la production de produits pharmaceutiques. C’est le cas par exemple de l’insuline, l’hormone de croissance, certains vaccins, des interférons… Certaines bactéries comme Streptomyces sont très employées pour la production d’antibiotiques.
Certaines bactéries peuvent provoquer une dégradation d'installation (biocorrosion), en particulier les bactéries sulfato-réductrices
Classification et Identification
La taxonomie permet de classer de façon rationnelle les organismes vivants. Chez les bactéries, les taxons dans l’ordre hiérarchique sont les suivant : phylums (ou divisions), classes, sous-classes, ordres, sous-ordres, familles, sous-familles, tribus, sous-tribus, genres, sous-genres, espèces et sous-espèces. Différentes approches permettent la classification des bactéries.
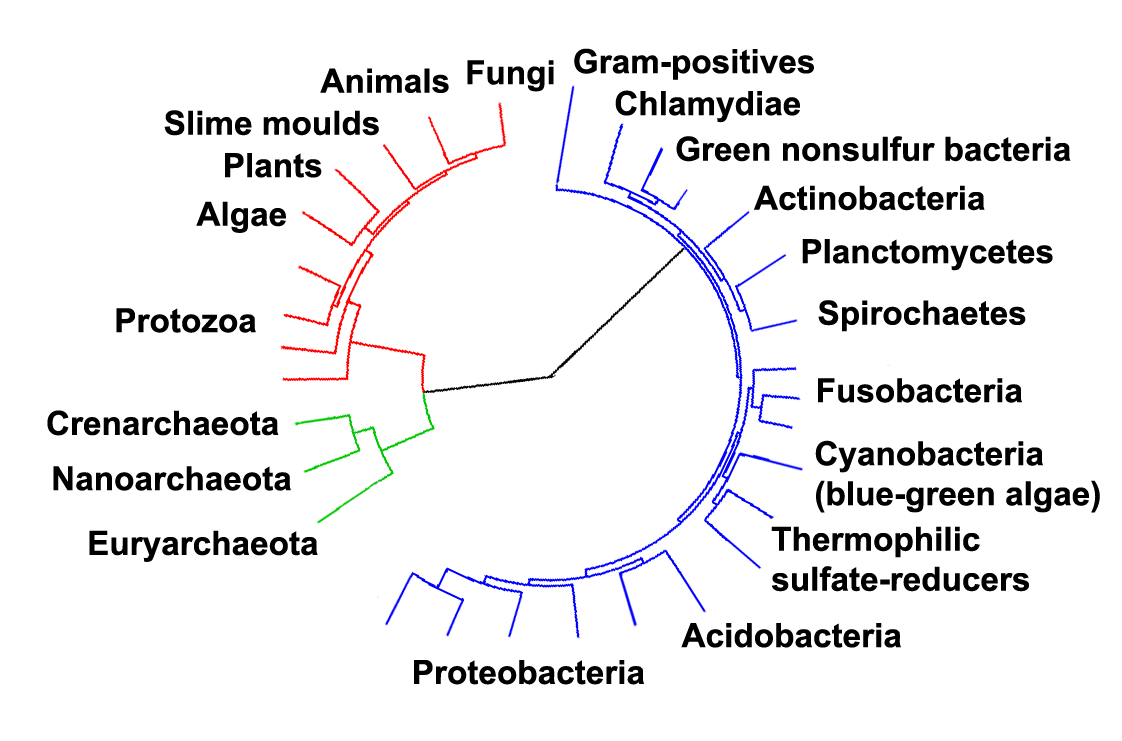
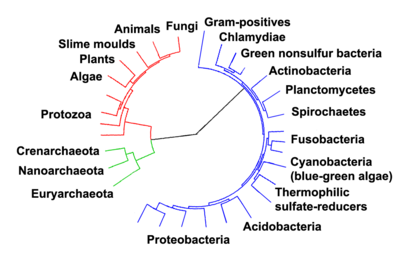 Arbre phylogénétique montrant la diversité des bactéries, comparés aux autres organismes[46]. Les eucaryotes sont colorés en rouges, les archaea en vert et les bactéries en bleu.
Arbre phylogénétique montrant la diversité des bactéries, comparés aux autres organismes[46]. Les eucaryotes sont colorés en rouges, les archaea en vert et les bactéries en bleu.Classification phénotypique
- Critères morphologiques (forme et groupement des bactéries, présence ou absence de flagelle, nature de la paroi, type de mobilité, présence d’endospore).
- Critères physiologiques (type métabolique, source d’énergie, de carbone, d’azote, type de substrat utilisé, capacité à produire certaines molécules, produits de fermentation, métabolites secondaires…).
- Critères de pathogénicité.
- Critères de sérogroupage.
Chimiotaxonomie
Il s’agit de l’analyse chimique de constituants cellulaires (structure et composition de la paroi, des membranes plasmiques, du peptidoglycane).
Classification moléculaire
- Composition en bases de l’ADN. Le pourcentage de guanine + cytosine varie d’un organisme à un autre, mais est relativement constant au sein d’une même espèce. Le pourcentage de G + C varie de 25 à 70 % chez les procaryotes.
- Hybridation ADN – ADN. Cette technique permet de comparer la totalité du génome bactérien et d'estimer le degré d'homologie entre deux bactéries. Cette caractéristique est importante dans la définition d’une espèce bactérienne.
- Séquençage des ARN ribosomiques ou séquençage des gènes codant les ARNr. Chez les procaryotes, la comparaison de la séquence en nucléotides de l'ARN 16S permet d'évaluer le degré de parenté entre ces organismes. Ces gènes sont qualifiés d’horloge moléculaire et permettent la classification phylogénétique.
Identification des espèces bactériennes
La détermination génétique des espèces se base sur l’étude des gènes des ARN ribosomiques. Le choix des gènes des ARNr 16S se justifie pour les raisons suivantes :
- les ARNr 16S sont des molécules ubiquistes ;
- leur structure est bien conservée car toute modification pourrait nuire à la synthèse protéique. Il en résulte une évolution très lente de ces gènes.
Le choix des gènes des ARNr plutôt que les ARNr eux-mêmes se base sur le choix de la technique de l’amplification par PCR. Cette technique permet, à partir d’une colonie de bactéries, d’obtenir des fragments d’ADN correspondants au gène ou à une partie du gène. Les analyses génétiques concernent également la région intergénique 16S-23S des opérons des ARN ribosomiques. Cette dernière est une région de longueur variable selon les organismes. Elle donne une indication immédiate sur le fait que deux souches données appartiennent ou non à la même espèce.
Tous les micro-organismes possèdent au moins une copie des gènes codant les ARN ribosomiques. Ces molécules sont indispensables à la synthèse des protéines, raison pour laquelle cette séquence d’ADN est très conservée au sein des espèces (plus de 99 %). Cette conservation de séquence permet d’utiliser cette région pour la détermination des espèces. En effet, Le degré de similarité des séquences d’ARNr entre deux organismes indique leur parenté relative. La procédure utilisant l’ARNr 16S comme facteur d'identification implique l'extraction de l’ADN des bactéries d’une colonie. Puis des amorces reconnaissant des zones très conservées du gène permettent d'amplifier par PCR une grande partie du gène ARNr 16S, qui par la suite est séquencé. Les données sur la séquence nucléotidique sont comparées avec des bases de données de séquences déjà connues. Les séquences du gène codant l’ARNr 16S sont connues pour plus de 4 000 souches bactériennes. Ces séquences peuvent être consultées par interrogation de banques de données telles que EMBL et GenBank par les logiciels Fasta et Blast. Le Ribosomal Data Project II (RDP) est également intéressant dans la mesure où sa base de données est spécifique de l’ARN 16S. Ces logiciels sont accessibles en ligne sur l’internet. Selon les différents auteurs, le degré d’homologie entre deux bactéries pour qu’elles appartiennent à la même espèce doit être supérieur à 97 %, voire 99 %.
Comme les gènes de la région intergénique 16S-23S sont moins conservés, ils diffèrent d’une souche à l’autre aussi bien au niveau de la séquence que de la longueur. Ceci résulte de ce que de nombreuses bactéries ont des copies multiples par génome de l’opéron de l’ARNr, il en résulte lors de l’amplification un motif caractéristique. Comme pour le gène de l’ARNr 16S, l’étude systématique de la région intergénique 16S-23S requiert l’amplification de cette région par PCR. L’utilité de la région intergénique 16S-23S est qu’elle permet de distinguer des espèces différentes et parfois différentes souches au sein de la même espèce. En effet la région intergénique étant moins conservée, des variabilités au niveau des séquences peuvent se présenter pour des souches de la même espèce mais appartenant à des biovars différents.
Les séquences de la région intergénique 16S-23S sont comparées par interrogation des bases de données IWoCS qui est spécifique de la région intergénique 16S-23S. La base de données GenBank est également très bien fournie. Le degré d’homologies devrait idéalement être proche de 100 % pour des souches identiques.
Anecdotes
Les plus anciennes bactéries en vie
Le 4 septembre 2007, un forage dans le pergélisol du nord-ouest Canadien a permis à des scientifiques de l'université de Californie dirigée par le professeur Eske Willerslev (Université de Copenhague) de mettre au jour une bactérie vieille d'environ 500 000 ans et toujours vivante.
On a retrouvé une bactérie endormie à l'intérieur d'une abeille qui était dans de l'ambre (résine fossile - provenant de conifères de l'époque oligocène, qui poussaient sur l'emplacement de l'actuelle mer Baltique - se présentant sous forme de morceaux durs et cassants, plus ou moins transparents, jaunes ou rougeâtres) depuis 25 à 40 millions d'années.
De même, une bactérie demeurée endormie depuis 250 millions d'années a été découverte dans un cristal de sel. Elle a été découverte par Russell Vreeland de l'université de West Chester en Pennsylvanie dans un lit de sel à environ 600 mètres sous terre, près de Carlsbad au Nouveau-Mexique.
Dans l'espace, les bactéries deviendraient presque trois fois plus virulentes. C'est du moins le cas de Salmonella typhimurim, une bactérie responsable d'intoxication alimentaire. Celles-ci ont fait un voyage à bord de la navette Atlantis en 2006. À leur retour, les bactéries qui avaient été conservées dans un récipient étanche, ont été transmise à des souris. Il n'a fallu que le tiers de la dose habituelle pour tuer la moitié du groupe de souris qui avait été infecté[47],[48].
Recherche de bactéries extraterrestres
On cherche actuellement à savoir s'il a existé une vie bactérienne sur la planète Mars. Certains éléments d'analyse du sol martien semblent s'orienter en ce sens, et la présence abondante d'eau sur Mars jadis a peut-être pu constituer un terrain extrêmement favorable au développement de la vie bactérienne, si elle est apparue. Si la chose venait à être confirmée, ce serait un élément important en faveur de l'hypothèse de panspermie.
Une chose semble certaine aujourd'hui (2006) : les différents appareils américains et européens envoyés sur Mars dans un but d'exploration de la planète y ont laissé une grande quantité de bactéries extrêmophiles d'origine terrestre.[49] Si ces bactéries terrestres arrivent à survivre en trouvant l'eau martienne profonde (qui parfois rejaillit en surface ou s'accumule en glace autour des pôles) et à s'adapter au milieu physicochimique et climatique de cette planète (notamment des extrémophiles capables d'utiliser les oxydes de fer et de carbone), elles pourraient contaminer et coloniser rapidement des sols qu'on cherche à explorer aujourd'hui, et même produire assez vite de nouvelles espèces spécifiquement martiennes, produisant à terme de profonds changements dans la chimie des sols, voire sur l'atmosphère très ténue de Mars.[réf. nécessaire]
D'autres recherches s'intéressent aussi aux glaces de la lune jovienne Europe qui abritent de l'eau liquide sous leur surface.
Références
- ↑ (en)Fredrickson J, Zachara J, Balkwill D, et al, « Geomicrobiology of high-level nuclear waste-contaminated vadose sediments at the Hanford site, Washington state », dans Appl Environ Microbiol, vol. 70, no 7, 2004, p. 4230–41 [texte intégral lien PMID lien DOI]
- ↑ a et b (en)(en) W. Whitman, D. Coleman et W. Wiebe, « Prokaryotes: the unseen majority », dans Proc Natl Acad Sci U S A, vol. 95, no 12, 1998, p. 6578–83 [texte intégral lien PMID lien DOI].
- ↑ (en)Rappé M, Giovannoni S. (2003) "The uncultured microbial majority". Annu Rev Microbiol 57: 369 – 94.
- ↑ (en) Textbook of Bacteriology : Flore bactérienne de l’Homme
- ↑ (en)2002 WHO mortality data
- ↑ (en)Ishige T, Honda K, Shimizu S, « Whole organism biocatalysis », dans Curr Opin Chem Biol, vol. 9, no 2, 2005, p. 174–80 [lien PMID lien DOI]
- ↑ (en)Porter JR, « Antony van Leeuwenhoek: Tercentenary of his discovery of bacteria », dans Bacteriological reviews, vol. 40, no 2, 1976, p. 260–269 [texte intégral lien PMID]
- ↑ (en)van Leeuwenhoek A, « An abstract of a letter from Mr. Anthony Leevvenhoek at Delft, dated Sep. 17, 1683, Containing Some Microscopical Observations, about Animals in the Scurf of the Teeth, the Substance Call'd Worms in the Nose, the Cuticula Consisting of Scales », dans Philosophical Transactions (1683–1775), vol. 14, 1684, p. 568–574 [texte intégral (page consultée le 2007-08-19)]
- ↑ (en)van Leeuwenhoek A, « Part of a Letter from Mr Antony van Leeuwenhoek, concerning the Worms in Sheeps Livers, Gnats, and Animalcula in the Excrements of Frogs », dans Philosophical Transactions (1683–1775), vol. 22, 1700, p. 509–518 [texte intégral (page consultée le 2007-08-19)]
- ↑ (en)van Leeuwenhoek A, « Part of a Letter from Mr Antony van Leeuwenhoek, F. R. S. concerning Green Weeds Growing in Water, and Some Animalcula Found about Them », dans Philosophical Transactions (1683-1775), vol. 23, 1702, p. 1304–11 [texte intégral lien DOI (pages consultées le 2007-08-19)]
- ↑ Etymology of the word "bacteria"
- ↑ (en)Pasteur's Papers on the Germ Theory
- ↑ (en)O'Brien S, Goedert J, « HIV causes AIDS: Koch's postulates fulfilled », dans Curr Opin Immunol, vol. 8, no 5, 1996, p. 613–618 [lien PMID lien DOI]
- ↑ (en)The Nobel Prize in Physiology or Medicine 1905
- ↑ (en)Thurston A, « Of blood, inflammation and gunshot wounds: the history of the control of sepsis », dans Aust N Z J Surg, vol. 70, no 12, 2000, p. 855–61 [lien PMID lien DOI]
- ↑ (en)Schwartz R, « Paul Ehrlich's magic bullets », dans N Engl J Med, vol. 350, no 11, 2004, p. 1079–80 [lien PMID lien DOI]
- ↑ (en)Biography of Paul Ehrlich
- ↑ Woese C, Kandler O, Wheelis M (1990). « Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya ». Proc Natl Acad Sci U S A 87
- ↑ (en)(en) Berg JM, Tymoczko JL Stryer L, Molecular Cell Biology, WH Freeman, 2002 (ISBN 0-7167-4955-6)
- ↑ (en)Gitai Z, « The new bacterial cell biology: moving parts and subcellular architecture », dans Cell, vol. 120, no 5, 2005, p. 577–86 [lien PMID lien DOI]
- ↑ (en)Shih YL, Rothfield L, « The bacterial cytoskeleton », dans Microbiol. Mol. Biol. Rev., vol. 70, no 3, 2006, p. 729–54 [texte intégral lien PMID lien DOI]
- ↑ (en)Gitai Z, « The new bacterial cell biology: moving parts and subcellular architecture », dans Cell, vol. 120, no 5, March 2005, p. 577–86 [lien PMID lien DOI]
- ↑ (en)Norris V, den Blaauwen T, Cabin-Flaman A, et al, « Functional taxonomy of bacterial hyperstructures », dans Microbiol. Mol. Biol. Rev., vol. 71, no 1, March 2007, p. 230–53 [texte intégral lien PMID lien DOI]
- ↑ (en)Nicholson W, Munakata N, Horneck G, Melosh H, Setlow P, « Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments », dans Microbiol Mol Biol Rev, vol. 64, no 3, 2000, p. 548–72 [texte intégral lien PMID lien DOI]
- ↑ (en)Siunov A, Nikitin D, Suzina N, Dmitriev V, Kuzmin N, Duda V, « Phylogenetic status of Anaerobacter polyendosporus, an anaerobic, polysporogenic bacterium », dans Int J Syst Bacteriol, vol. 49 Pt 3, p. 1119–24 [texte intégral lien PMID]
- ↑ (en)Nicholson W, Fajardo-Cavazos P, Rebeil R, Slieman T, Riesenman P, Law J, Xue Y, « Bacterial endospores and their significance in stress resistance », dans Antonie Van Leeuwenhoek, vol. 81, no 1–4, 2002, p. 27–32 [lien PMID lien DOI]
- ↑ Vreeland R, Rosenzweig W, Powers D, « Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal », dans Nature, vol. 407, no 6806, 2000, p. 897–900 [lien PMID lien DOI]
- ↑ (en)Cano R, Borucki M, « Revival and identification of bacterial spores in 25- to 40-million-year-old Dominican amber », dans Science, vol. 268, no 5213, 1995, p. 1060–4 [lien PMID lien DOI]
- ↑ (en)Nicholson W, Schuerger A, Setlow P, « The solar UV environment and bacterial spore UV resistance: considerations for Earth-to-Mars transport by natural processes and human spaceflight », dans Mutat Res, vol. 571, no 1–2, 2005, p. 249–64 [lien PMID]
- ↑ (en)Hatheway C, « Toxigenic clostridia », dans Clin Microbiol Rev, vol. 3, no 1, 1990, p. 66–98 [texte intégral lien PMID]
- ↑ Nakabachi A, Yamashita A, Toh H, Ishikawa H, Dunbar H, Moran N, Hattori M (2006). The 160-kilobase genome of the bacterial endosymbiont Carsonella. Science 314 (5797): 267
- ↑ Pradella S, Hans A, Spröer C, Reichenbach H, Gerth K, Beyer S (2002). « lang » Sorangium cellulosum So ce56. Arch Microbiol 178 (6): 484-92.
- ↑ Hinnebusch J, Tilly K (1993). « lang ». Mol Microbiol 10 (5): 917-22.
- ↑ Brüssow H, Canchaya C, Hardt W (2004). « lang ». Microbiol Mol Biol Rev 68 (3): 560-602.
- ↑ Perna N, Mayhew G, Pósfai G, Elliott S, Donnenberg M, Kaper J, Blattner F (1998). « lang ». Infect Immun 66 (8): 3810-7.
- ↑ Denamur E, Matic I (2006). « lang ». Mol Microbiol 60 (4): 820 – 7.
- ↑ Davison J (1999). « lang ». Plasmid 42 (2): 73 – 91.
- ↑ Hastings P, Rosenberg S, Slack A (2004). « lang ». Trends Microbiol 12 (9): 401
- ↑ Schulz H, Jorgensen B. (2001). « lang ». Annu Rev Microbiol 55 : 105 – 37
- ↑ Robertson J, Gomersall M, Gill P. (1975). « lang ». J Bacteriol. 124 (2): 1007 – 18.
- ↑ (en)Wu M, Roberts J, Kim S, Koch D, DeLisa M, « Collective bacterial dynamics revealed using a three-dimensional population-scale defocused particle tracking technique », dans Appl Environ Microbiol, vol. 72, no 7, 2006, p. 4987–94 [texte intégral lien PMID lien DOI]
- ↑ (en)Bardy S, Ng S, Jarrell K, « Prokaryotic motility structures », dans Microbiology, vol. 149, no Pt 2, 2003, p. 295–304 [texte intégral lien PMID lien DOI]
- ↑ (en)Lux R, Shi W, « Chemotaxis-guided movements in bacteria », dans Crit Rev Oral Biol Med, vol. 15, no 4, 2004, p. 207–20 [lien PMID]
- ↑ (en)Frankel R, Bazylinski D, Johnson M, Taylor B, « Magneto-aerotaxis in marine coccoid bacteria », dans Biophys J, vol. 73, no 2, 1997, p. 994–1000 [lien PMID]
- ↑ (en)O'Hara A, Shanahan F, « The gut flora as a forgotten organ », dans EMBO Rep, vol. 7, no 7, 2006, p. 688–93 [lien PMID lien DOI]
- ↑ (en)Ciccarelli FD, Doerks T, von Mering C, Creevey CJ, Snel B, Bork P, « Toward automatic reconstruction of a highly resolved tree of life », dans Science, vol. 311, no 5765, 2006, p. 1283–7 [lien PMID lien DOI]
- ↑ (en)Lethal Bacteria Turn Deadlier After Space Travel
- ↑ Des bactéries plus virulentes après un séjour dans l’espace, MICROBIOLOGIE, Cécile Dumas, article du 12 juin 2008 du NouvelObs.com reprenant l’article de Sciences et Avenir.com du 25 septembre 2007, consulté le 19 novembre 2008
- ↑ Demidov VV, Goncharov AA, Osipov VB, Trofimov VI., Modern aspects of planetary protection and requirements to sterilization of space hardware, Adv Space Res. 1995 Mar;15(3):251-5.
Voir aussi
Articles connexes
Bibliographie
- Madigan, M. T., Martinko, J. M. Brock Biology of Microorganisms, 11th Ed. Pearson Prentice Hall, Upper Saddle River, NJ, 2005.
- Perry J., Staley J., Lory S. Microbiologie. Éditions Dunod, 2004.
- Prescott, L.M., Harley, J.P. Klein, D.A., Microbiologie, 2e édition DeBoeck eds, 2003.
- Singleton P. Bactériologie : pour la médecine, la biologie et les biotechnologies. Cours, 6e édition, Dunod, 2005.
- François Denis, Marie-Cécile Ploy, Christel Martin, Edouard Bingen. "Bactériologie médicale - Techniques usuelles". Masson, 2007
Liens externes
- Référence Tree of Life Web Project : Bacteria (en)
- Référence Catalogue of Life : Bacteria (en)
- Référence AlgaeBase : genre Bacteria Shadbolt 1854 (en)
- Référence World Register of Marine Species : taxon Bacteria (en) (+liste espèces)
- Référence NCBI : Bacteria (en)
- Morphologie et structure des bactéries (J.P. Euzéby)
- Voir un schéma détaillé d'une cellule bactérienne
- (de) Bacterial Growth and Cell Wall
 Portail de la microbiologie
Portail de la microbiologie Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant
Catégorie : Bactérie




Wikimedia Foundation. 2010.