- Conjugaison (Génétique)
-
Conjugaison (génétique)
 Pour les articles homonymes, voir Conjugaison (homonymie).
Pour les articles homonymes, voir Conjugaison (homonymie).En biologie, la conjugaison est une méthode non sexuée utilisée par les bactéries afin de s'échanger des informations génétiques. Elle consiste en une transmission de plasmides de conjugaison d'une bactérie donneuse à une bactérie receveuse et, potentiellement, son intégration dans le génome de celle-ci.
Sommaire
Utilité de la conjugaison
Chez les procaryotes (bactéries), la multiplication est asexuée : elle est réalisée par de simples divisions cellulaires. Lors de ces divisions, le génome de l'individu est multiplié par deux puis est réparti équitablement entre la cellule mère et la cellule fille. Le brassage génétique, qui est essentiel à la survie d'une espèce puisqu'il permet indirectement aux individus de s'adapter à leur environnement, est donc inexistant.
C'est pourquoi plusieurs autres techniques permettant d'assurer une diversité génétique suffisante se sont développées chez les bactéries.- Les mutations ponctuelles aléatoires
- Mais l'évolution induite par ces mutations est très lente et n'est pas contrôlée par la bactérie.
- Les transferts horizontaux :
- Beaucoup plus rapides, ce sont ces mécanismes qui sont à l'origine d'une grande partie de la diversité génétique des microorganismes. Il existe trois mécanismes : La transduction, la transformation et la conjugaison. C'est cette dernière qui est décrite ici.
Les conséquences des ces transferts pour les bactéries sont énormes. Les gènes de résistance aux antibiotiques, par exemple, peuvent très rapidement se propager dans la population mondiale de bactéries. Progressivement, les bactéries acquièrent une résistance à tous les antibiotiques. Les échanges sont si efficaces que la population mondiale bactérienne peut être modélisée par un organisme unique s'échangeant des informations à la manière de cellules dans un corps humain. Les résistances aux antibiotiques ne sont pas les seules concernées, d'autres résistances peuvent elles aussi être transférées :Ce sont ces mécanismes qui, au fil du temps, ont transformé la bactérie bien connue Escherichia coli en Shigella flexnerie, responsable de dysenterie.
Transfert du plasmide conjugatif
Le transfert entre les organismes donneur et accepteur de plasmide se fait en quatre grandes étapes :
- Reconnaissance entre donneur (F+) et accepteur (F-) grâce au pilus
- Transfert d'un des deux brins du plasmide
- Synthèse du brin complémentaire chez l'accepteur
- Recirculisation du plasmide chez l'accepteur
Reconnaissance et contact
La reconnaissance se fait par l'intermédiaire du pilus. Le pilus est une structure protéique qui sert de lien entre le donneur et l'accepteur. Il est synthétisé par le donneur en utilisant le code génétique présent sur le plasmide. Il est constitué de filaments protéiques (polymère de piline) et a la forme d'un tube creux.
Le pilus est essentiel pour l'initiation de l'agrégation donneur-récepteur. En effet, il y a une interaction entre le pilus et certaines protéines (protéine OMPA) présentes sur l'extérieur de la membrane de la bactérie acceptrice. Il s'agit de la phase de reconnaissance. Pour que le transfert se fasse, l'agrégation doit être stable durant 1 à 2 minutes. Pour cela des protéines spécifiques (de type adhésine) reconnaissent aussi les protéines OMPA sur la bactérie acceptrice et stabilisent l'ensemble. Sans l'aide de ces protéines le complexe ne tiendrait en place que 30 à 40 ms.
Une fois en place, le pilus sert à rapprocher les deux bactéries et à les amener au contact. Ce rapprochement se fait par dépolymérisation du milieu du pilus, raccourcissant ainsi le pont entre les deux bactéries et tirant les deux cellules l'une vers l'autre.
 Figure 1 : Schéma de la rétraction du pilus.
Figure 1 : Schéma de la rétraction du pilus.
F+ Bactérie donneuse F- Bactérie acceptriceUne fois ce contact réalisé, il y a réorganisation du pilus qui se transforme en port de sécrétion. Le reste du pilus sert à perforer la membrane de l'accepteur afin de faire passer le plasmide.
Transfert
Une fois que l'agrégat des deux bactéries est stabilisé, il y a transfert d'un des deux brins du plasmide. Un ensemble de protéines va se former pour aider à ce processus : c'est le relaxosome. Le relaxosome contient les protéines nécessaires à la coupure de l'ADN, au désenroulement du brin (grâce à des hélicases) et au déplacement du brin vers le port de sécrétion. Il y a donc coupure simple brin du plasmide au niveau de son origine de transfert (oriT) puis, le brin va "glisser" vers la bactérie hôte dans le sens 5' vers 3'.
En même temps, du côté de la bactérie donneuse, le brin complémentaire est resynthétisé afin de garder un plasmide double brin.
Synthèse du brin complémentaire
Une fois introduite dans la bactérie acceptrice, l'ADN simple brin, très fragile, est stabilisé par des protéines (SSB). Il y a alors synthèse du brin complémentaire, afin de former un nouveau plasmide double brin. Cette synthèse se fait de façon discontinue. Des protéines primases synthétisent des amorces à partir desquelles les ADN polymérases pourront démarrer leur synthèse.
La recircularisation du nouveau plasmide se fait grâce à une protéine (relaxase) qui va permettre la jonction des deux bouts.
Régulations
Il existe plusieurs niveaux de régulation dans le phénomène de conjugaison. Il paraît par exemple nécessaire de réguler l’expression des gènes permettant la conjugaison. Les plasmides, dans leur bactérie hôte, ont leurs gènes de conjugaison réprimés par défaut puisqu'exprimer ces gènes alors que la bactérie ne peut pas ou n'a pas besoin de faire de conjugaison est un gaspillage d'énergie.
Il y a activation des gènes de conjugaison quand :
- la bactérie acceptrice reçoit un plasmide;
- la bactérie est soumise à un stress (nutritif, physique...);
- la bactérie arrive au terme d'un cycle biologique : le mécanisme de régulation est tel qu'un cycle se met en place avec des périodes de répression des gènes et des périodes d'activations.
L'activation des gènes de conjugaison entraîne un compte à rebours vers leur inactivation. Les bactéries sont donc momentanément capables de conjugaison puis perdent cette capacité avant la prochaine stimulation. Le phénomène de conjugaison dans une population bactérienne se fait donc par poussée invasive violente.
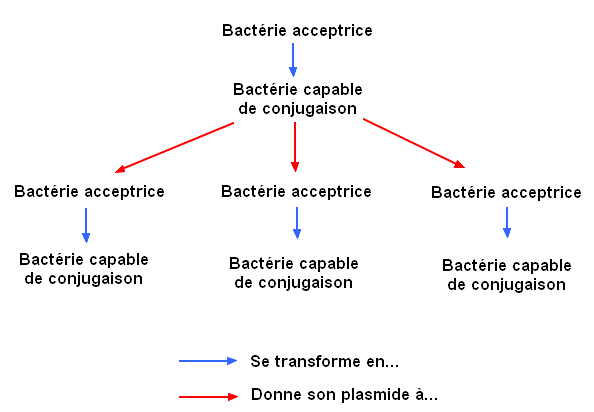 Figure 2 : Schéma de la poussée invasive de conjugaison parmi une population bactérienne.
Figure 2 : Schéma de la poussée invasive de conjugaison parmi une population bactérienne.Exclusion donneur/donneur
La synthèse du pilus et l'initiation de la conjugaison sont des processus très coûteux en énergie pour la bactérie. Si la cellule avec laquelle la bactérie donneuse entre en contact possède déjà un plasmide, cette dépense d'énergie est inutile. C'est pour cela que certains systèmes se sont mis en place afin d'éviter la conjugaison entre deux donneurs.
Le premier système est l'exclusion de surface.
Si un plasmide est présent dans une bactérie, celui-ci permet le codage de lipoprotéines de la membrane externe (TRAT). Ces protéines sont semblables aux protéines OMPA qui servent de message signal et sur lesquelles le pilus peut se fixer afin d'initier l'agrégation. La différence est qu'elles ont une affinité pour le pilus supérieur que les protéines OMPA et qu'une fois le pilus fixé, le processus de conjugaison est arrêté.
Ce système permet à lui seul de diviser par 10 à 20 fois les contacts pilus-OMPA.Le deuxième système est l'exclusion d'entrée. Ici, une protéine de la membrane interne (TRAS) va bloquer le passage des signaux envoyés par le donneur (signal de mobilisation) après l'agrégation. Ces signaux sont nécessaires au bon déroulement de la conjugaison et le blocage des ces signaux arrête la conjugaison.
Ce système permet à lui seul de diviser par 100 à 200 fois les cas de transferts entre deux bactéries donneuses.Les plasmides conjugatifs
Description
Le plasmide F (Fertilité) est un plasmide modèle. Il a été très étudié pour comprendre les mécanismes de régulation qui gouvernent la conjugaison. Il fait environ 100 kpb ce qui fait de lui un gros plasmide. Beaucoup des gènes présents sur ce plasmide ont encore un rôle inconnu, mais les fonctions de transfert (mises en jeu pendant la conjugaison) ont été très étudiées.
Ces fonctions de transferts sont codées par 36 gènes organisés en un seul opéron et répartis sur 34 kpb environ. Les gènes liés au transfert sont appelés tra et trb.En résumé, le plasmide est constitué de trois grandes catégories de gènes :
- Les gènes tra
- Les gènes de résistances
- Les gènes de métabolisme
Et plusieurs fonctions très importantes :
- La fonction ori
- La fonction oriT
- La fonction part
Les gènes tra sont donc impliqués dans les phénomènes de régulation, la synthèse du pilus, la stabilisation de l'agrégation, l'exclusion donneur-donneur et dans le métabolisme de transfert.
La fonction ori est l'origine de réplication. C'est à partir d'elle que commence la réplication des plasmides et c'est elle qui contrôle le nombre de copies d'un même plasmide présent dans une bactérie.
La fonction oriT est l'origine de transfert, c'est à partir de lui que va commencer le désenroulement simple brin du plasmide avant le transfert vers la bactérie acceptrice. C'est dans cette zone aussi que se trouvent les systèmes de stabilisation des plasmides qui ont pour but le maintien de la présence des plasmides dans la population bactérienne.
La fonction part est la fonction de partition. Elle contrôle la répartition des plasmides présents dans la bactérie lors de la division cellulaire.Critère d'incompatibilité
Deux plasmides dans une même bactérie hôte sont dits incompatibles si les deux ne se retrouvent pas ensemble dans les bactéries des générations suivantes.
Cette incompatibilité est due aux fonctions ori et part... Deux plasmides s'excluant mutuellement, c'est-à-dire ne pouvant coexister dans la même bactérie, appartiennent au même groupe d'incompatibilité.Mobilisation de gène bactérien par conjugaison
- À compléter
Références
liens externes :
- (fr)http://www.ac-creteil.fr/biotechnologies/doc_microbio-bacterialconjugation.htm
- (en)http://bcs.whfreeman.com/mga2e/pages/bcs-main.asp?s=003&n=72&i=147&v=category&o=|60|72|&ns=0%20&uid=0&rau=0
Littératures :
 Portail de la microbiologie
Portail de la microbiologie Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire
Catégories : Biologie moléculaire | Génétique | Microbiologie - Les mutations ponctuelles aléatoires
Wikimedia Foundation. 2010.