- Chimiotactisme
-
Chimiotaxie
La chimiotaxie, un des types de la taxie, est le phénomène par lequel les cellules corporelles, les bactéries, ainsi que les autres organismes uni-, ou multicellulaires dirigent leurs mouvements en fonction de certaines espèces chimiques présentes dans l’environnement. Trouver de la nourriture (par exemple du glucose) en se dirigeant vers sa concentration la plus élevée est très important pour les bactéries, aussi bien qu’éviter les agents nuisibles (tel que le phénol par exemple). Chez les êtres vivants pluricellulaires la chimiotaxie joue un rôle insigne dans le développement et dans le fonctionnement physiologique de l’organisme. Or, il a été découvert que les mécanismes permettant la chimiotaxie dans les animaux peuvent être supprimés lors de la formation des métastases cancéreuses.
Sommaire
Histoire de la recherche de la chimiotaxie
 Numéraires de la recherche du chimiotactisme
Numéraires de la recherche du chimiotactisme
Bien que la migration des cellules a été découverte dès le début du développement du microscope (Leeuwenhoek), la première description érudite a été faite par T. W. Engelmann (en 1881) et par W.F. Pfeffer (en 1884) chez les bactéries et par H.S. Jennings (en 1906) chez les organismes ciliés. E. Metchnikov, lauréat du prix Nobel, a également contribué à la recherche de ce domaine en étudiant ce processus comme première étape de la phagocytose. L’importance biologique et pathologique de la chimiotaxie est devenue largement acceptée pendant les années 1930. La définition la plus fondamentale du phénomène a été également formulée à cette époque. Les aspects primordiaux du contrôle de qualité d’un essai chimiotactique ont été décrits par H. Harris dans les années 1950.
Pendant les deux décennies suivantes, la révolution de la biologie cellulaire moderne ainsi que celle de la biochimie ont assuré le développement d'une large gamme de techniques nouvelles pour l’étude des cellules donnant de réponse migratoire et des fractions subcellulaires responsables pour le chimiotactisme. Le travail pionnier de J. Adler a représenté un tournant au niveau de la compréhension du processus entier de la transmission intracellulaire des signaux chez les bactéries. [1] Le 3 novembre, 2006, Dr. Dennis Bray (Université de Cambridge) a été honoré par le prix « Microsoft European Science Award » pour ses recherches concernant la chimiotaxie de l’E. coli. [2],[3]Phylogénie et signalisation chimiotactique
La chimiotaxie est une des réponses physiologiques les plus fondamentales. Le développement des systèmes de récepteurs permettant la détection des substances nuisibles ou favorables de l’environnement était essentiel pour les organismes unicellulaires dès les premières étapes de la phylogénie. L’analyse compréhensive du chimiotactisme du Tetrahymena pyriformis -eucaryote protozoaire- et celle des séquences consensus des acides aminés présents dans la soupe primordiale suggère l’existence d’une forte corrélation entre le caractère chimiotactique de ces composés relativement simples et leur apparition sur la Terre. Ainsi, on suppose que les premières molécules étaient fortement chimioattractantes (par exemple Gly, Glu, Pro), tandis que les acides aminés apparus plus tardivement pouvaient être des chimiorepellants forts (par exemple Tyr, Trp, Phe). [4]
La chimiotaxie bactérienne
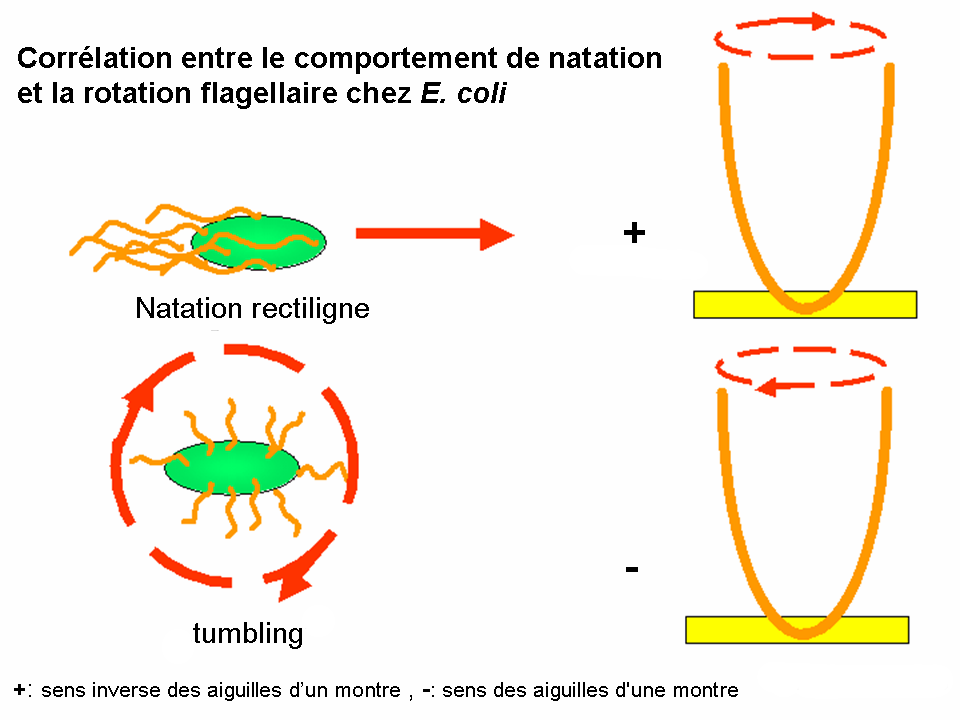
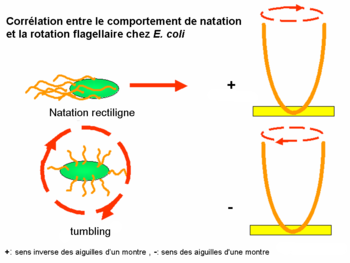 Corrélation entre le comportement de natation et la rotation flagellaire chez E. coli
Corrélation entre le comportement de natation et la rotation flagellaire chez E. coliCertaines bactéries, dont par exemple Escherichia coli, ont quelques flagelles (en général 4-10 par cellule). Ces derniers peuvent tourner dans deux sens :
- La rotation dans le sens inverse des aiguilles d’un montre (sens trigonométrique+) arrange les flagelles dans un seul faisceau synchrone; il en résulte la natation rectiligne de la bactérie. (phase appelée « run » en anglais)
- Lors de leur rotation dans le sens des aiguilles d'une montre (sens trigonométrique-), les flagelles se désolidarisent les uns des autres, ainsi la bactérie bouge de manière erratique et tourne dans tous les sens mais ne peut pas se déplacer. (phase appelée « tumble » en anglais)
Le sens de la rotation est toujours observé en dehors de la cellule, en regardant les flagelles en face de la cellule.
Comportement
Le mouvement résultant de la bactérie est donc dû à l’alternance des deux phases (« run » et « tumble »). Si on observe le mouvement d’une bactérie dans un environnement homogène on trouve une trajectoire aléatoire due aux phases de natations rectilignes relativement courtes, interrompues par des phases de « tumble » aléatoires qui réorientent les bactéries. Les bactéries tel que E. coli sont incapables de choisir la direction dans laquelle elles nagent et ne sont pas capables de nager longtemps suivant une ligne droite à cause de la diffusion rotationnelle. Autrement dit, les bactéries « oublient » la direction dans laquelle elles vont. Tenant compte de ces contraintes, il est remarquable que les bactéries sont capables de diriger leur mouvement afin de trouver des lieux favorables de concentration élevée en attractants (en général en nourriture) ainsi que d’éviter les substances répellantes (en générales les agents toxiques).
À la présence d’un gradient chimique les bactéries vont répondre avec la chimiotaxie, elles vont diriger donc leur mouvement en fonction du gradient chimique. Si la bactérie sent qu’elle bouge dans la bonne direction (c’est à dire elle va vers l’attractant ou bien elle s’éloigne du repellant) elle continue de nager tout droit pour plus longtemps avant la phase « tumbling ». Dans le cas contraire, elle va arrêter de nager plus tôt et essayer une nouvelle direction choisie au hasard. Autrement dit, les bactéries comme E. coli utilisent une perception temporelle pour décider si la vie commence à devenir meilleure ou pire. De cette façon, elles trouvent assez bien les lieux où la concentration des attractants est la plus élevée (en général la source). De plus, même en cas de concentrations très élevées, elles sont capables de distinguer des différences très faibles de concentration. La détection des reppellants fonctionne avec la même efficacité. Il paraît remarquable que cette marche aléatoire soit le résultat d’un choix simple entre deux formes de mouvement aléatoire, notamment entre la natation et le « tumbling » . En effet, les réponses chimiotactiques telles qu’oublier la direction et choisir le type du mouvement ressemblent à la capacité de prendre des décisions des êtres vivants plus évolués possédant un cerveau avec lequel elles sont capable de traiter les informations sensorielles.La structure hélicale du filament flagellaire unique est cruciale pour l'exécution de ces mouvements. Par ailleurs, la protéine constituant le filament flagellaire, la flagelline, est assez similaire chez toutes les bactéries ayant des flagelles. Les vertébrés semblent tirer un bénéfice de ce fait : il possèdent un récepteur immunologique (TLR5) conçu pour la reconnaissance de cette protéine conservée.
Comme c’est souvent le cas dans la biologie, certaines bactéries ne suivent pas la règle. Beaucoup de bactéries, telle que Vibrio, n’ont qu’un seule flagelle polaire (ciliature monotriche). Leur méthode de chimiotaxie est différente. D’autres possèdent un flagelle qui se situe entièrement dans la paroi. Quand ces bactéries bougent, la cellule entière dont la forme rassemble à une tire-bouchon, est mise en rotation. [5]Transmission des signaux
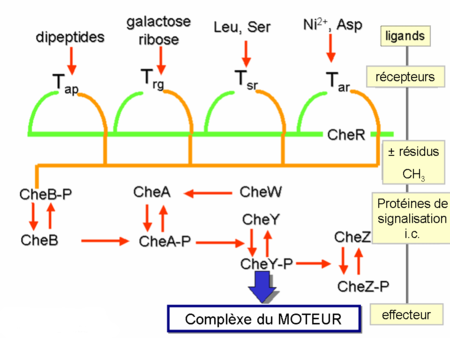
Le gradient chimique est perçu à l’aide de plusieurs récepteurs transmembranaires, appelés « methyl accepting chemotaxis proteins (MCPs) » qui varient par rapport à la molécule détectée. Ces récepteurs fixent des attractants ou des repellants, soit directement soit indirectement, via des interactions avec des protéines du périplasme. Ces signaux sont ensuite transmis des récepteurs -à travers de la membrane plasmique- au cytosol où les protéines Che deviennent activées. Les protéines Che altèrent la fréquence de « tumbling » et les récepteurs.
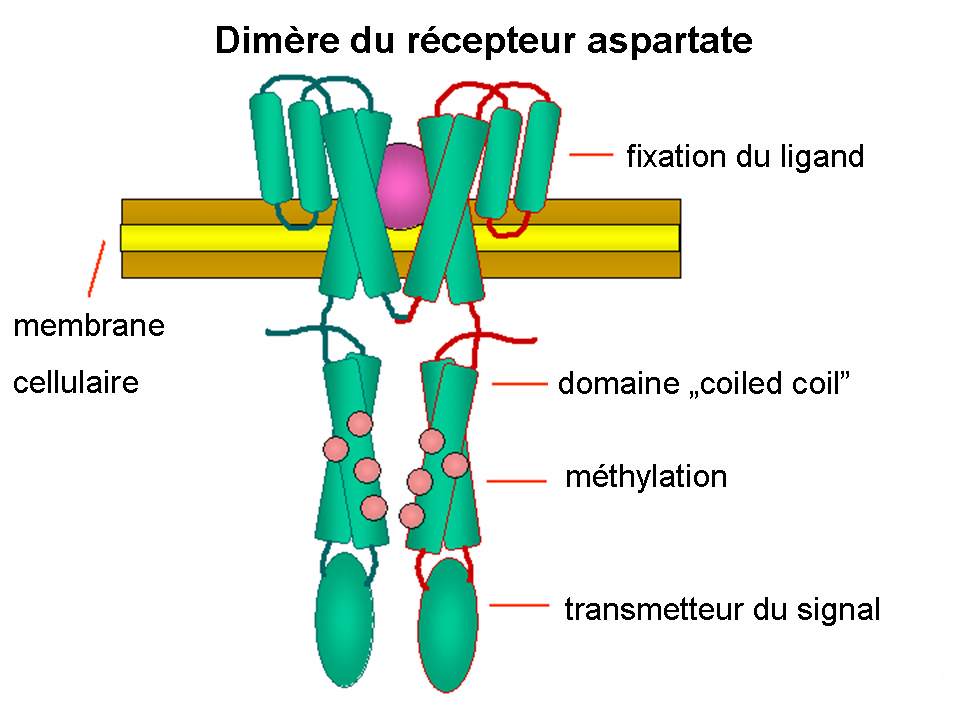
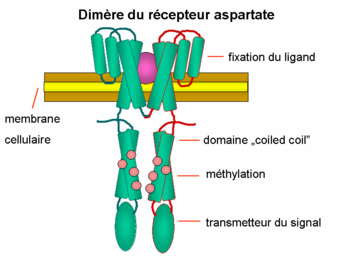 Dimère du récepteur aspartate
Dimère du récepteur aspartateRégulation des flagelles
Les protéines CheA et CheW se fixent au récepteur. L’activation du récepteur par un stimulus externe résulte l’autophosphorylation de l'histidine kinase CheA à un résidu unique de histidine fortement conservé. Puis, CheA transfère un groupe phosphoryl aux résidus aspartates conservés des régulateurs de réponse, CheB et CheY. [remarque : CheA est une histidine kinase et ne transmet pas le groupe phosphoryl d’une façon active. C’est le régulateur de réponse CheB qui le prend à partir de CheA.] Ce mécanisme de transmission de signal s’appelle un « système à deux composants » qui est une forme très répandue de la transmission des signaux chez les bactéries. CheY en interagissant avec la protéine commutateur flagellaire, la protéine FliM, induit le changement du sens de rotation des flagelles du sens inverse des aiguilles d’une montre au sens des aiguilles d'une montre et ainsi induit le «tumbling » . La modification de l’état de rotation d’un seul flagelle peut perturber le faisceau entier et provoquer le « tumbling ».
Régulation de récepteur
Une fois activée par CheA, CheB agit comme une méthyle estérase et enlève des groupes méthyles des résidus glutamates de la partie cytoplasmique du récepteur. Elle a un effet antagoniste par rapport à la méthyle transferase CheR qui méthyle les mêmes résidus de glutamates. Plus de groupes de méthyle sont rajoutés au récepteur, plus ce dernier est sensible. Comme le signal à partir du récepteur entraîne la déméthylation de ce dernier par un mécanisme de boucle de feedback, le système est toujours ajusté au niveau des composés chimiques dans l’environnement. Ainsi, il reste sensible aux changements faibles, mêmes pour les concentrations extrêmes. Cette régulation permet à la bactérie de ‘se souvenir’ des concentrations récentes et les comparer avec les concentrations actuelles, donc de savoir si elle se déplace dans le sens du gradient ou au contraire, dans le sens inverse du gradient. Pourtant, le système de méthylation seul n’explique pas le spectre large de sensibilité des bactéries aux différents gradients chimiques. Des mécanismes régulatoires supplémentaires tels que le « clustering » des récepteurs et les interactions de type récepteur-récepteur modulent également la voie de signalisation.
 La voie de signalisation de l'E.coli
La voie de signalisation de l'E.coliChimiotaxie eucaryotique
Bien que le mécanisme de la chimiotaxie des eucaryotes soit assez différent de celui des bactéries, la perception du gradient chimique reste une étape cruciale du processus. À cause de leur taille, les procaryotes ne sont pas capables de détecter les gradients de concentration effectifs, donc ces cellules "scannent" et évaluent leur environnement en nageant continuellement. (La natation étant la série des étapes consécutives de natation droite et de « tumbling ».) Les dimensions des cellules eucaryotes, en revanche, permettent la détection du gradient. Cela résulte d'une distribution dynamique et polarisée des récepteurs. L’induction de ces derniers par des chimioattractants ou par des chimiorepellants provoque la migration vers la substance de l’activité chimiotactique ou au contraire, dans la direction opposée.
 Différence de la percéption du gradient chez les procaryotes et les eucaryotes
Différence de la percéption du gradient chez les procaryotes et les eucaryotesLes niveaux des récepteurs, les voies de signalisation et les mécansimes effecteurs représentent tous des divers componants de type eucaryotiques. Chez les eucaryotes unicellulaires les effecteurs majeurs sont les pseudopodes, les cils ou les flagelles de type eucaryote (par exemple l’Amoeba ou le Tetrahymena). Quelques cellules eucaryotes d’origine mammifère plus évoluées, telles que celles du système immunitaire sont également capables de se déplacer au lieu nécessaire. A part des cellules immunocompétentes (granulocyte, monocyte, lymphocyte), un groupe nombreux des cellules considérées auparavant étant fixées sur des tissus, est également mobile sous certaines conditions physiologiques (mastocyte, fibroblaste, cellules endothéliales) ou pathologiques (par exemple métastases). La chimiotaxie joue un rôle important lors du début de l’embryogenèse car la formation des couches du germe est guidée par les gradients des molécules de signal.
Motilité
Contrairement à la motilité pendant la chimiotactisme des bactéries, le mécanisme permettant le déplacement physique des cellules eucaryotes est encore peu élucidé. Probablement, il existe un mécanisme par lequel un gradient chimiotactique extracellulaire est perçu et transformé en un gradient de phosphatidylinositol 1,4,5 tris phosphate (PIP3) intracellulaire. Cela résulte un gradient de l’activation de la voie de signalisation qui mène à la polymérisation des filaments d’actine. L’élongation de l’extrémité (+) des filaments d’actine assure une connection avec la surface interne de la membrane cytoplasmique via différents ensembles de peptides. Ainsi, elle résulte la formation des pseudopodes. Les cils des cellules eucaryotes sont également capables de provoquer la chimiotactisme, mais dans ce cas il s’agit plutôt d’une induction Ca2+ dépendant du système microtubulaire du corps basal et du 9×2+2 microtubules (axonème) du cils. Le battement orchestré des milliers de cils est synchronisé par un système submembranaire formé entre les corps basaux. Les détails de la voie de signalisation ne sont pas encore complètement éclairés.
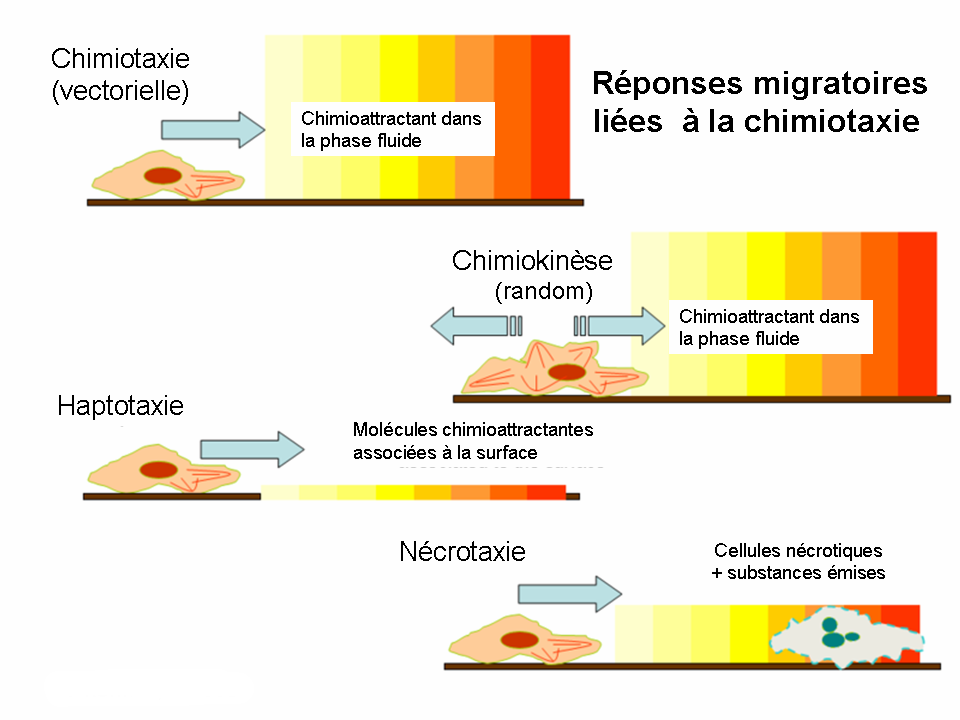
Réponses migratoires liées à la chimiotaxie
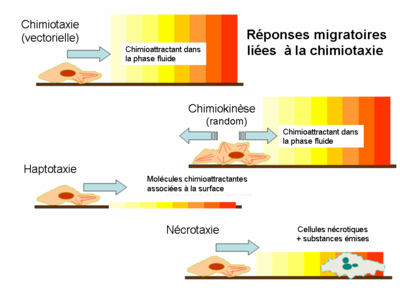
Bien que la chimiotaxie soit la forme de migration la plus fréquemment étudiée, il existe encore quelques autres types de locomotion au niveau cellulaire.
- Le chimiokinèse est également induit par les molécules de la phase liquide de l’environnement. Cependant, la réponse déclenchée n’est pas un mouvement vectoriel mais une taxie aléatoire. Ni l’amplitude, ni la fréquence du déplacement n’ont de caractéristiques ou de direction bien définis car ce comportement assure plutôt le scanning de l’environnement que la migration entre deux points distincts.
- Pendant l’haptotaxie le gradient de chimioattractant se présente sur une surface tandis que pendant la chimiotaxie classique le gradient se développe dans une phase soluble. La surface biologique active du point de vue de l’haptotaxie la plus importante est la matrice extracellulaire (MEC); la présence des ligands fixés est responsable pour l’induction de la migration transendothéliale et celle de l’angiogénèse.
- La nécrotaxie représente un type spécial de la chimiotaxie, lors duquel le chimioattractant est issu d’une cellule nécrotique ou apoptotique. En fonction du caractère chimique de la substance émise, la nécrotaxie peut accumuler ou repousser les cellules d’où l’importance pathophysiologique de ce phénomène.
 Réponses migratoires liées à la chimiotaxie
Réponses migratoires liées à la chimiotaxieRécepteurs
Dans la majorité des cas, les cellules eucaryotes perçoient la présence des stimuli chimiotactiques via des récepteurs de 7 domaines transmembranaires (récepteurs serpentines), couplés aux protéines G qui sont des hétèrotrimères. Ces récepteurs se regroupent dans une famille très nombreuse représentant une proportion significative du génome. Certains membres de cette superfamille de gènes interviennent pendant la vue (rhodopsins) ou bien lors de l’olfaction. Les classes majeures des récepteurs professionnels responsables pour la chimiotaxie sont stimulées par les peptides formylés (formyl peptide receptors (FPR)), par les chimiokines (chemokine receptors (CCR ou CXCR)) et par les leukotriènes (leukotriene receptors (BLT)). Pourtant l’induction d’une large gamme de récepteurs membranaires (par exemple ceux des acides aminés, de l’insuline, des peptides vasoactifs) déclenche également la migration de la cellule.
Sélection chimiotactique
Certains récepteurs chimiotactiques sont exprimés sur la surface membranaire et ont des caractéristiques à long terme car ils sont déterminés génétiquement. Cependant, il en existe d’autres ayant une dynamique à court terme, assemblés d’une façon ad hoc en présence du ligand. La différence des caractéristiques des récepteurs chimiotactiques et de celles des ligands permet la sélection des cellules ayant une réponse chimiotactique par un simple essai chimiotactique (« chemotaxis assay »). Par la sélection chimiotactique il est possible de déterminer si une molécule encore non caractérisée agit via les voies de récepteur long terme ou court terme. Le terme sélection chimiotactique peut également désigner une technique permettant de séparer les cellules eucaryotes ou procaryotes en fonction de leur capacité de répondre au ligand de sélection par la chimotaxie.
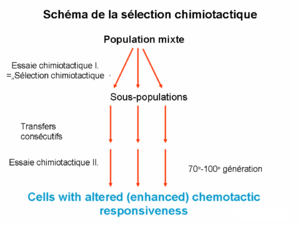 Sélection chimiotactique
Sélection chimiotactiqueLigands chimiotactiques
Le nombre de molécules capables de déclencher une réponse chimiotactique est relativement élevé. De plus, nous pouvons distinguer des molécules chimiotactiques primaires et secondaires. Les groupes majeurs des ligands primaires sont les suivants :
- Les peptides formylés sont des di-, tri-, tetrapeptides d’origine bactérienne (voir groupe formyl à l’extrémité N des peptides). Ils sont émis de la bactérie soit in vivo soit après la décomposition de la cellule. Un membre typique de ce groupe est la N-formylmethionyl-leucyl-phenylalanine (fMLF ou fMLP dans les références). La fMLF issue des bactéries est l’élément-clé de l’inflammation car elle a un effet chimioattractant caractéristique sur les granulocytes neutrophiles ainsi que sur les monocytes.
- Fragment C3a et fragment C5a sont des produits intermédiaires de la cascade protéolitique du système de complément. Leur synthèse est un résultat commun des 3 voies possibles (voie « classique », voie des lectins, voie « alterne ») de l’activation du système et se fait par l’enzyme convertase. Les cellules cibles majeures de ces dérivés sont également les granulocytes neutrophiles ainsi que les monocytes.
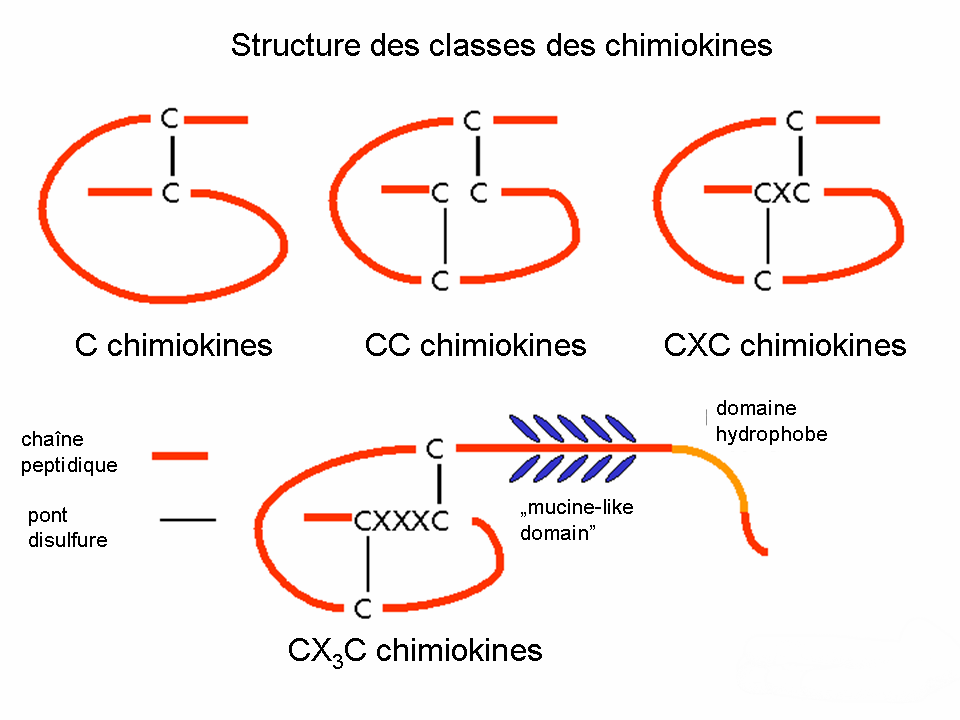
- Les chimiokines appartiennent à une classe spéciale des cytokines. Les diverses familles (C, CC, CXC, CX3C chimiokines) représentent non seulement des molécules de structure similaire par rapport à la position des ponts disulfure, mais la spécificité de cible des groupes est également différente. Les CC chimiokines ont un impact sur les monocytes (p.e. RANTES), les CXC chimiokines sont spécifiques pour les granulocytes neutrophiles (p.e. IL-8).
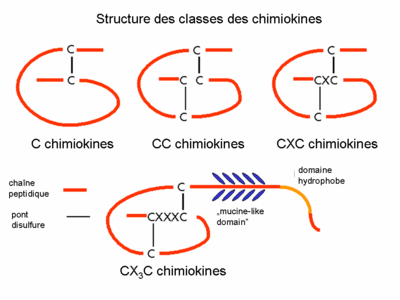 Structure des classes des chimiokines
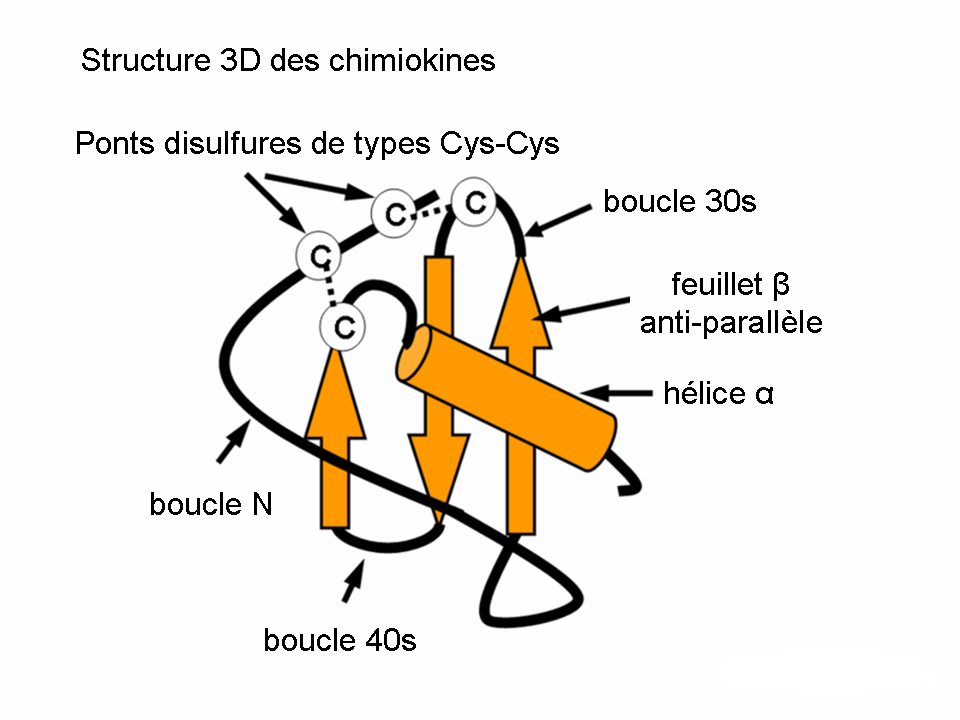
Structure des classes des chimiokinesL’étude de la structure 3D des chimiokines a prouvé qu’une composition caractéristique des motifs feuillet-β et une hélice-α assure l’expression de la séquence nécessaire pour l’interaction avec le récepteur de chimiokine. La formation d’un dimère ainsi que l’activité biologique accrue ont été démontrées par la christallographie d’un certain nombre de chimiokines (p.ex. IL-8)
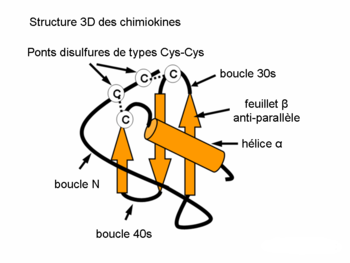 Structure 3D de l'Il-8
Structure 3D de l'Il-8- Les leucotriènes appartiennent au groupe des eicosanoïdes. Ils sont des médiateurs importants de la cascade de l’acide arachidonique converti par la 5-lipoxigènase. Le membre prédominant de la famille est le leukotriène B4 (LTB4) qui induit l’adhérence, la chimiotaxie et l’agrégation des leukocytes. Le pouvoir chimioattractant caractéristique du LTB4 est induit via les récepteurs (spanning leukotriene receptors) de 7 domaines transmembranaires, couplés aux protéines G. L’expression de ces récepteurs est très importante lors de l’inflammation et l’allergie.
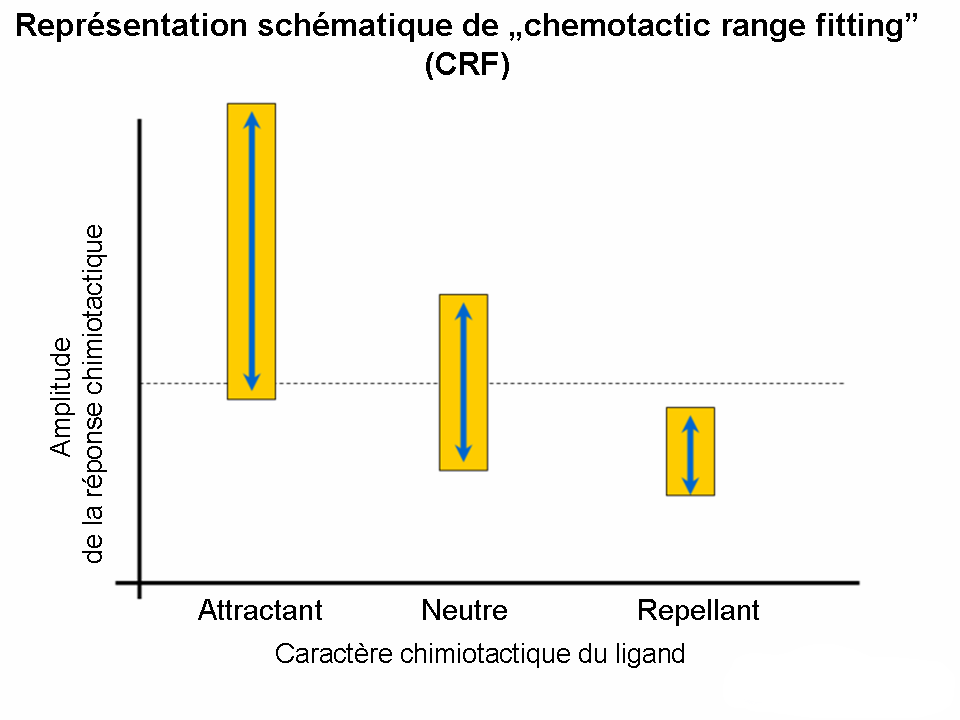
« Chemotactic range fitting » (CRF)
Les réponses chimiotactiques déclenchées par les interactions entre les ligands et les récepteurs sont en général distinguées à la base de la concentration effective optimale du ligand. Or, la corrélation entre l’amplitude de la réponse et la proportion des cellules répondant est également une propriété caractéristique de la signalisation chimiotactique. Les études des différentes familles de ligands (p. ex. Acides aminés, oligopeptides) ont prouvé l’existence d’une correspondance (« fitting of ranges ») entre les gammes (amplitudes, nombre des cellules répondant) et le chimiotactisme. Le pouvoir chimioattractant est accompagné par des gammes larges, tandis que le caractère chimioreppellant par des gammes étroites.
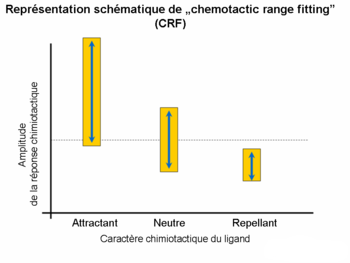 Chemotactic range fitting
Chemotactic range fittingImportance clinique
Le changement éventuel du potentiel migratoire des cellules a une importance relativement élevée dans le développement de certains symptômes et syndromes cliniques. Le chimiotactisme altéré des pathogènes extracellulaires (p. ex. Escherichia coli) et intracellulaires (p. ex. Listeria monocytogenes) représente en soi une cible clinique significative. La modification de la capacité chimiotactique intrinsèque de ces micro-organismes par des agents pharmaceutiques peut diminuer voire inhiber le taux des infections ou la propagation des maladies contagieuses. En plus des infections, il existe d’autres maladies dont l’origine (there are some other diseases where impaired chemotaxis is the primary étiologique factor,) est une chimiotaxie restreinte (endommagée) telles que le syndrome de Chediak-Higashi pendant dans lequel des vésicules intracellulaires géantes inhibent la migration normale des cellules.
Chimiotaxie (Chtx.) lors des maladies Type de la maladie Chtx. accrue Chtx. défectueuse Infections inflammations AIDS, Brucellose Chtx. cause la maladie - Syndrome de Chediak-Higashi , Syndrome de Kartagener Chtx. est touchée athérosclérose, arthrite, parodontite, psoriasis, blessure de reperfusion, metastase sclérose en plaque, Maladie de Hodgkin, stérilité (mâle) Intoxications asbestos, benzopyrène sels de Hg et de Cr, ozone (O3) Les publications scientifiques
La recherche de la migration cellulaire, comme on l’a pu lire dans le chapitre sur son histoire, nécessite l’application simultanée des techniques classiques et modernes qui se complètent. En même temps, cela permet de fournir des données enrichissantes aussi bien sur le plan de la recherche fondamentale que sur celui de la recherche appliquée. C’est grâce à ce fait que durant les 20-25 dernières années, le nombre des publications s’intéressant à la chimiotaxie en tant que phenomène s’est accru. De plus, les chapitres fournissant des informations sur la migration cellulaire ou bien sur la chimiotaxie constituent une partie précieuse de nombreuses publications appartenant au domaine de la génétique, de la biochimie, de la physiologie cellulaire, de la pathologie et au domaine clinique. L’activité de publication sur la recherche de la migration a comme particularité que le nombre des travaux étudiant la chimiotaxie ressort parmi ceux étudiant les diverses formes du déplacement cellulaire (thermotaxie, géotaxie, phototaxie, etc). Ce fait montre l’importance biologique est médicale exceptionnelle de ce type de migration.
Mesure de la chimiotaxie
Une large gamme de techniques est disponible pour évaluer l’activité chimotactique des cellules ainsi que le caractère chimioattractant ou chimioreppelant des ligandes. Les exigences de base des mesurages sont les suivantes :
- que le gradient de concentration puisse se développer relativement vite et puisse subsister pendant longtemps dans le système.
- que les activités chimiotactiques et chimiocinétiques soient distinguables.
- que la migration des cellules soit libre dans les deux directions opposées suivant l’axe du gradient de concentration
- que la réponse détectée soit le résultat d’une migration active des cellules.
Malgré le fait que l’essai chimiotactique idéal ne soit toujours pas disponible, il existe pourtant certains protocoles et pièces d’équipement offrant une correspondance acceptable avec les conditions énumérées ci-dessus. Les plus utilisés sont :
- « Agar plate assays » (migration sous agarose)
p. ex. PP-chamber
- « Two-chamber techniques »
p. ex. Chambre de Boyden, chambre de Zigmond, chambre de Dunn, chambre « Multi-well » Techniques capillaires
- Autres
p. ex. Technique de « T-maze » – technique d’opalescence – essai d’orientation (Vous trouvez une chapitre plus détaillée à « Chemotaxis assay ») Pour être capables de se mouvoir, les cellules ont besoin de plusieurs composants cellulaires (tels que les moteurs cellulaires, diverses enzymes, etc.). De plus, elles doivent être capables de changer leur forme. Dans un sens large le mouvement cellulaire a deux types... Hapoptatique (qui signifie un mouvement en réponse des stimuli physiques ou méchaniques). Chimiotactique (qui est un mouvement en réponse d’un gradient chimique).
Références
- ↑ Julius Adler and Wung-Wai Tso, « Decision-Making in Bacteria: Chemotactic Response of Escherichia Coli to Conflicting Stimuli », dans Science, vol. 184, 1974, p. 1292–4
- ↑ http://research.microsoft.com/displayArticle.aspx?id=1572 consulté le 6 novembre 2006
- ↑ http://news.bbc.co.uk/2/hi/science/nature/6113522.stm consulté le 6 novembre 2006
- ↑ The 'Optimal' Chemotactic Ligand - Amino acids consulté le 3 février 2008
- ↑ Howard C. Berg, « E. coli in motion », dans Springer-Verlag, NY., vol. ISBN 0-387-00888-8, 2003
Liens externes
Catégories : Physiologie cellulaire | Perception



Wikimedia Foundation. 2010.