- Homme de Néandertal
-
 Homme de Néandertal
Homme de Néandertal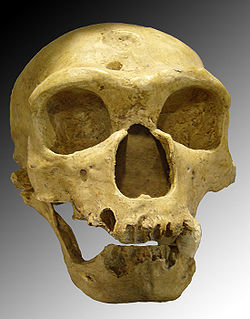
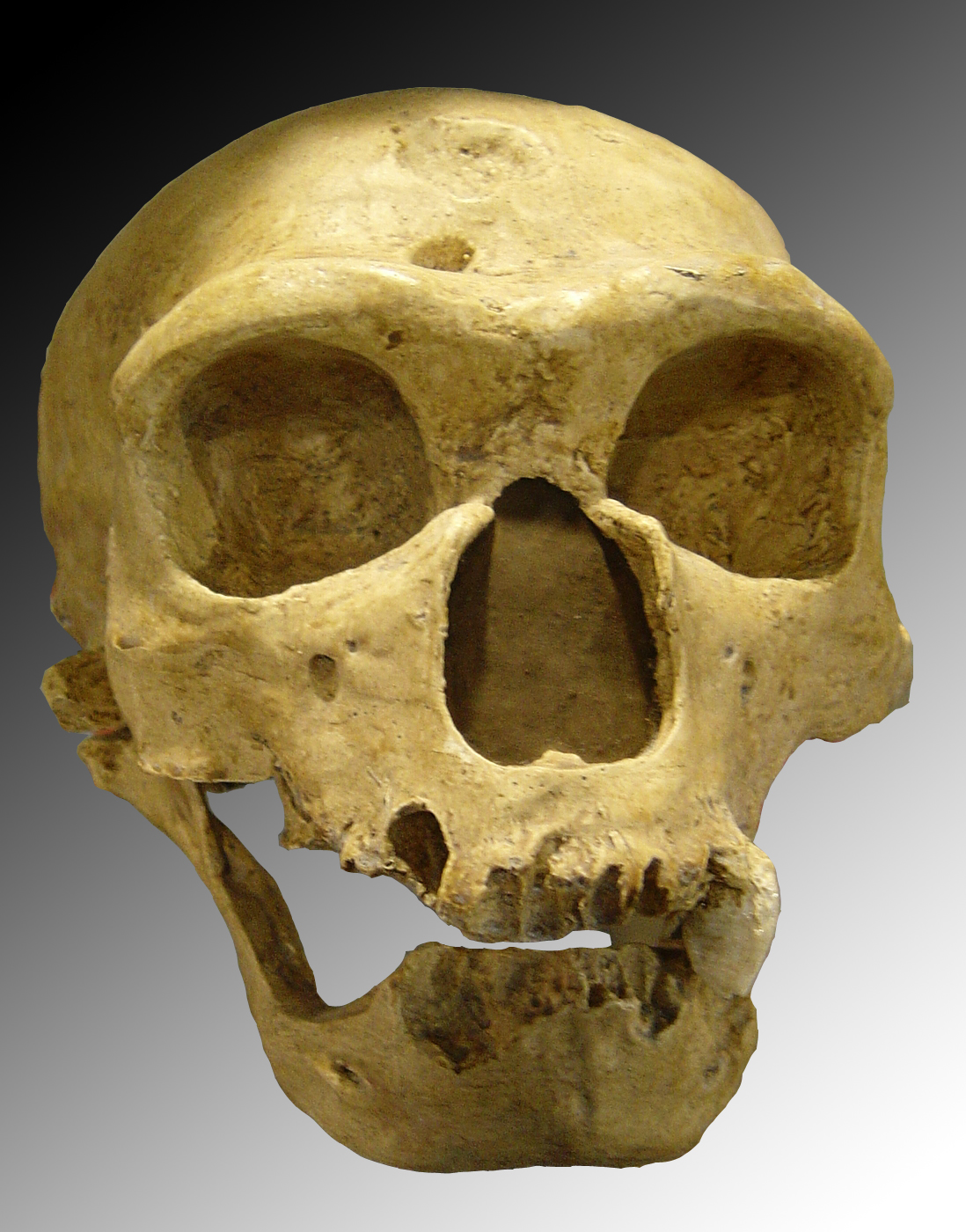
Crâne de Néandertalien :
l'Homme de la Chapelle-aux-Saints.Classification Règne Animalia Sous-règne Metazoa Super-embr. Deuterostomia Embranchement Chordata Sous-embr. Vertebrata Classe Mammalia Sous-classe Theria Infra-classe Eutheria Ordre Primates Sous-ordre Haplorrhini Infra-ordre Simiiformes — non-classé — Catarrhini Super-famille Hominoidea Famille Hominidae Sous-famille Homininae Tribu Hominini Genre Homo Nom binominal Homo neanderthalensis
William King, 1864 Retrouvez ce taxon sur Wikispecies
Retrouvez ce taxon sur Wikispecies
D'autres documents multimédia
sont disponibles sur CommonsUn homme de Néandertal ou Néandertalien est un représentant fossile du genre Homo qui a vécu en Europe et en Asie occidentale au Paléolithique moyen, entre environ 250 000 et 28 000 ans avant le présent. Durant plus d'un siècle à compter de sa découverte, les hypothèses émises à son sujet ont reflété les préjugés du moment : longtemps considéré comme une sous-espèce au sein de l'espèce Homo sapiens, nommée par conséquent Homo sapiens neanderthalensis, il fut ensuite considéré par la majorité des auteurs comme une espèce indépendante nommée Homo neanderthalensis. En 2010, le séquençage partiel de l'ADN nucléaire néandertalien par une équipe coordonnée par Svante Pääbo pourrait démontrer un métissage ancien entre les hommes de Néandertal et les humains anatomiquement modernes en Eurasie[1]. Ces résultats remettent en question l'idée selon laquelle ces deux groupes correspondent à des espèces distinctes[2],[3].
Premier homme fossile reconnu, premier être humain disparu distinct de l'Homme actuel, l'homme de Néandertal est à l'origine d'une riche culture matérielle appelée Moustérien ainsi que des premières préoccupations esthétiques et spirituelles en Europe (sépultures). Après une difficile reconnaissance, l'homme de Néandertal a longtemps pâti d'un jugement négatif par rapport aux Homo sapiens. Il est encore considéré dans l'imagerie populaire comme un être simiesque, fruste, laid et attardé. Il est en fait plus robuste qu'Homo sapiens et son cerveau est légèrement plus volumineux en moyenne. Les progrès de l'archéologie préhistorique et de la paléoanthropologie depuis les années 1960 ont mis au jour un être d'une grande richesse culturelle.
De nombreux points sont encore à élucider notamment concernant le moment de son apparition et les causes de son extinction, après plus de 300 000 ans[4] d'existence, vers 28 000 ans avant le présent.
Historique de la découverte[5],[6]
Les Néandertaliens avant Neandertal
 Le vallon des Grottes Schmerling à Engis en Belgique, où furent découverts les premiers ossements néandertaliens
Le vallon des Grottes Schmerling à Engis en Belgique, où furent découverts les premiers ossements néandertaliens
Deux fossiles de Néandertaliens ont été découverts peu avant celui de l'individu éponyme. En 1830, un crâne d'enfant est mis au jour par P.C. Schmerling à Engis (Belgique). En 1848, un crâne d'adulte est trouvé à Gibraltar dans le site de Forbe's Quarry. Si le premier appartient à un jeune individu sur lequel les traits caractéristiques des Néandertaliens sont moins évidents, le deuxième aurait dû conduire à reconnaître l'existence d'une espèce humaine fossile. Sans doute était-il trop tôt, comme le prouvent d'ailleurs les difficultés pour faire admettre que les os recueillis à Neandertal correspondent bien à un homme fossile.
1856 : Neandertal ou la « vallée de l'homme nouveau »
Le nom de « Néandertalien » est lié à celui de Neandertal, une petite vallée située sur les territoires des deux villes Erkrath et Mettmann, entre Düsseldorf et Wuppertal (Allemagne). Au mois d'août 1856, dans le cadre de l'exploitation d'une carrière, des ouvriers vidèrent une petite cavité de cette vallée, la grotte de Feldhofer. Ils y découvrirent des ossements et un fragment de crâne qu'ils remirent à Johann Carl Fuhlrott, instituteur d'Elberfeld, passionné d'histoire naturelle.
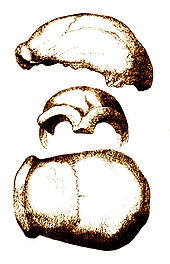 Calotte crânienne découverte à Neandertal en 1856
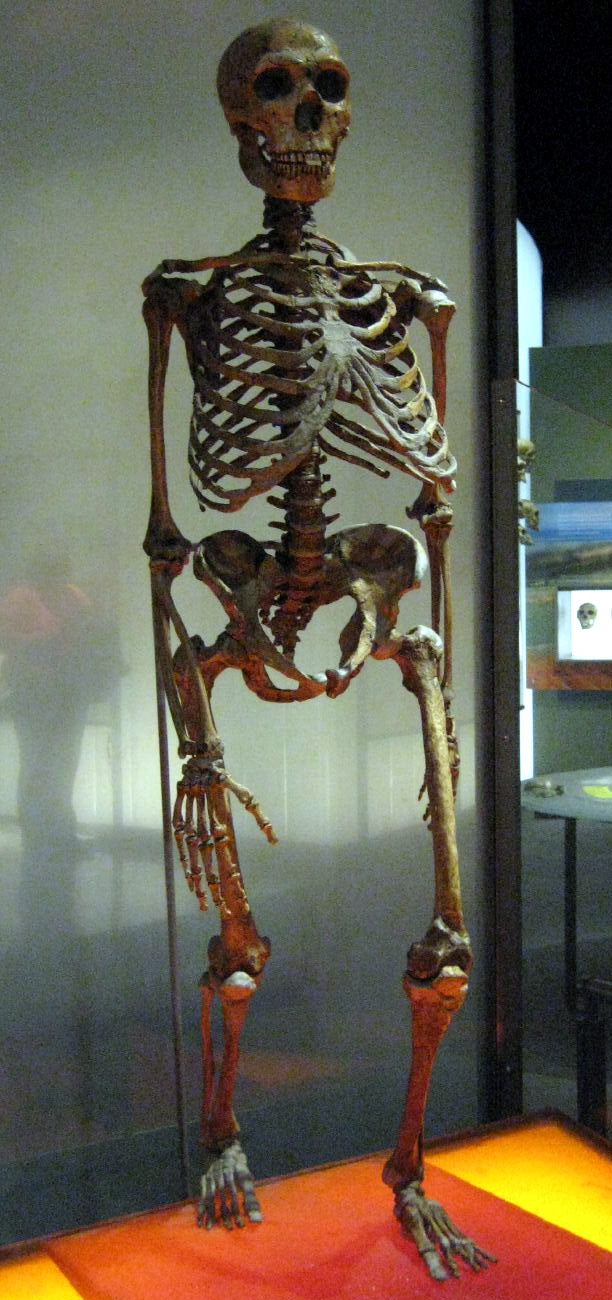
Calotte crânienne découverte à Neandertal en 1856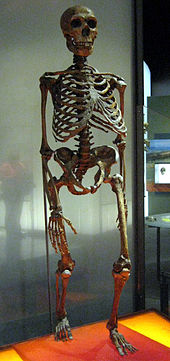 Squelette reconstitué d'un homme de Néandertal
Squelette reconstitué d'un homme de NéandertalPar un heureux hasard, le toponyme Neandertal signifie « vallée de l'homme nouveau ». Le nom de Neander avait été donné à cette vallée (tal en allemand) en l’honneur de Joachim Neumann (1650-1680), dit aussi Joachim Neander car, suivant un usage familial datant de son grand-père, il se faisait appeler par son patronyme traduit en grec. Ce pasteur et compositeur, auteur de cantiques religieux encore populaires dans le protestantisme allemand, aimait chercher son inspiration dans cette vallée, jadis idyllique.
Comme à l'époque le nom de la vallée s'écrivait encore « Neanderthal », l'homme qui y est découvert reçoit le nom latin de Homo neanderthalensis ; quand une réforme orthographique de l'allemand eut balayé les « h » « superflus », les règles de la nomenclature interdisaient de revenir sur les formes latines et on écrit toujours Homo neanderthalensis. L'orthographe française la plus correcte et la plus courante, proposée par Henri Vallois en 1952, est Homme de Néandertal, même si l'on trouve parfois Homme de Néanderthal, Homme de Neandertal ou Homme de Neanderthal. La forme « Neanderthal » est encore très répandue en anglais, induisant une prononciation en contradiction avec le nom allemand originel.
Une reconnaissance difficile
Fuhlrott comprend rapidement l'intérêt de la découverte et se rend sur place pour tenter en vain de découvrir d'autres ossements ou des vestiges qui leur seraient associés. Il se rend compte qu'il s'agit d'ossements anciens mais surtout incroyablement primitifs, correspondant à un homme nouveau, d'une « conformation naturelle jusqu'ici inconnue[7],[8] ».
L'Homme de Néandertal est effectivement le premier homme fossile distinct d'Homo sapiens, et il est découvert le premier. L'idée même qu'une espèce d'homme distincte de la nôtre ait existé par le passé (et ait disparu) fut d'ailleurs particulièrement difficile à admettre. On se souviendra par exemple que Charles Darwin ne publiera L'Origine des espèces par la sélection naturelle qu'en 1859 et qu'il n'élargira explicitement sa théorie à l'homme qu'en 1871 dans La Filiation de l'homme et la sélection liée au sexe.
Malgré des différences importantes avec les os d'hommes modernes, Fuhlrott reconnaît dans ses trouvailles des os humains et les soumet à Hermann Schaaffhausen pour un examen complémentaire. Ce dernier présente ses premières conclusions en 1857[9]. Il estime que les ossements datent d'une période antérieure aux Celtes et aux Germains, et sont ceux d'un individu appartenant à l'une des races sauvages du nord-ouest de l'Europe dont parlent les auteurs latins. Tous les chercheurs n'acceptent pas cette interprétation : pour certains, les os ont appartenu à un genre différent du nôtre, sans doute plus proche du singe, pour d'autres ils renvoient à un individu pathologique ou frappé d'idiotisme. Certains évoquent même un cosaque ayant déserté les armées russes en 1814.
Établissement de l'ancienneté de l'Homme et de son évolution
Peu à peu les découvertes se multiplient, d'abord celles d'Homo sapiens fossiles associés à des vestiges lithiques et à des animaux disparus (dont l'Homme de Cro-Magnon en 1868) puis d'autres Homo neanderthalensis, encore en place dans les sédiments, complets et présentant les mêmes spécificités anatomiques, mais souvent hors contexte archéologique (pas d'ossements d'animaux ou d'outils associés), y compris en mer du Nord[10], ce qui rend difficile leur datation et leur interprétation. Parmi les plus spectaculaires, il faut citer les deux squelettes de la Grotte de Spy en 1886 puis la sépulture de l'Homme de la Chapelle-aux-Saints en 1908. Elles contribuent à faire définitivement accepter l'existence d'une nouvelle espèce d'humain par la communauté scientifique.
Le nom scientifique Homo neanderthalensis est proposé en 1864 par William King, professeur au Queen's College de Galway en Irlande et ancien élève de Charles Lyell[11]. En 1866, Ernst Haeckel propose le nom surprenant d’Homo stupidus, qui n'est pas retenu en vertu des règles de nomenclature donnant priorité à l'appellation antérieure. Les partisans du rattachement à une sous-espèce parleraient sinon d’Homo sapiens stupidus !
Les premières études (et les reconstitutions qui en découlaient) donnent de l'Homme de Néandertal une image déformée, accentuant les traits primitifs, voire simiesques. C'est le cas de l'étude de l'Homme de la Chapelle-aux-Saints publiée par Marcellin Boule en 1911 : même s'il s'agit d'une étude très complète, référence pendant de nombreuses années, elle présente un Homme de Néandertal voûté, la colonne vertébrale courbée (comme chez les gorilles) et les membres inférieurs semi-fléchis[12]. Il faut presque un siècle à la communauté scientifique pour corriger cette perception influencée par des a priori peu scientifiques.
Homo sapiens neanderthalensis ou Homo neanderthalensis ?
 Reconstitution d'un chasseur.
Reconstitution d'un chasseur.La position phylogénétique exacte de l'homme de Néandertal provoque encore de nombreux débats : certains considèrent qu'il représente une sous-espèce au sein de l'espèce Homo sapiens et le nomment donc Homo sapiens neanderthalensis tandis que d'autres considèrent qu'il représente une espèce indépendante et le nomment Homo neanderthalensis. Il ne s’agit pas d’un simple problème de classification, il s’agit aussi de savoir si l’Homme de Néandertal représente une lignée parallèle et éteinte (espèce indépendante) ou bien s’il a pu contribuer en partie au patrimoine génétique de l’homme actuel. Deux sous-espèces peuvent se croiser et avoir une descendance fertile, mais c'est beaucoup plus variable pour deux espèces différentes, certaines le peuvent et d'autres pas - c'est l'infertilité de la descendance qui signe l'existence de deux espèces mais l'inverse n'est pas vrai.
Lors de son identification, l’hypothèse d’une espèce distincte a été privilégiée. Mais dans les années 1960, tous les Hominidés à l’exception des Australopithèques ont été regroupés dans le genre Homo. Les Néandertaliens ont alors été considérés comme une sous-espèce d'Homo sapiens. Cette hypothèse était alors soutenue par de nombreux spécialistes, comme le généticien Theodosius Dobzhansky ou le biologiste Ernst Mayr qui déclarait que « jamais plus d'une seule espèce d'homme n'a existé au même moment ». Aujourd’hui, l’idée d’espèces distinctes est à nouveau proposée, notamment grâce aux apports de la génétique.
En effet, les multiples études paléoanthropologiques effectuées sur les ossements ne permettent pas de se prononcer clairement sur la classification de l'homme de Néandertal. De récentes analyses comparées d'ADN mitochondrial extrait d'ossements de Néandertaliens, d'Homo sapiens anciens et de formes supposées intermédiaires indiqueraient un cheminement séparé des lignées humaine et néandertalienne pendant 500 000 ans[13],[14] ; il pourrait donc s'agir de deux espèces différentes. Dans ce cas, l'ancêtre commun de l'Homme de Néandertal et d’Homo sapiens serait probablement un Homo erectus qui vivait 500 000 ans plus tôt.
Indices défavorables via l'ADN mitochondrial
La question n'est pas résolue, la difficulté étant de concilier deux notions d'espèce (biologique et paléontologique) dont l'unicité est contestée. La possibilité pour ces deux représentants du genre Homo d'avoir une descendance féconde serait une preuve irréfutable, mais là encore les indices sont contradictoires. Aucun élément n'a permis d’établir la possibilité de croisements, même partiels, entre les Néandertaliens et les ancêtres de l'homme moderne qui lui étaient contemporains. Les analyses d’ADN mitochondrial des hommes de Néandertal découverts autrefois et récemment laissent penser que Néandertaliens et sapiens modernes n'ont échangé aucun de leurs gènes lors de leur coexistence (autour de 30 000 ans avant notre ère). Il s’agit pour l’instant de résultats partiels, portant sur les gènes qui ont pu être comparés, à savoir 370 paires de base sur 600 ; jusqu'à ce que l'étude soit complète, un échange de gènes ne peut être complètement exclu.
En Afrique et au Proche-Orient, une coexistence des deux taxons durant plus de 10 000 ans est vraisemblable. En Europe, cette coexistence n'a duré qu'un peu plus de 5 000 ans.
Un squelette d'enfant trouvé en contexte Gravettien à Lagar Velho dans la vallée de Lapedo, au centre du Portugal, porterait des caractéristiques des deux espèces[15],[16],[17]. Cet enfant d'environ quatre ans a été inhumé dans une sépulture intentionnelle, il y a 25 000 ans. Il n’est donc postérieur que de quelques milliers d'années aux derniers restes clairement attribuables aux Néandertaliens, datant d'environ 30 000 ans avant notre ère. Cependant, le caractère hybride de cet enfant est très discuté et difficile à établir : les caractères dérivés des deux taxons sont moins marqués chez les individus juvéniles que chez les adultes et la variabilité individuelle des enfants de l’époque est absolument inconnue[18].
Enfin, l'analyse publiée en 2006[19],[20] d'une séquence d'ADN mitochondrial de la racine d'une molaire d'un enfant néandertalien datant de - 100 000 ans, mise au jour dans la grotte de Scladina à Sclayn (Belgique), vient à l'appui de la thèse faisant de Néandertal une espèce différente d’Homo sapiens. La séquence d'ADN mt de cette dent présente une grande distance par rapport aux autres séquences connues de Néandertaliens, ce qui montre la grande diversité génétique de l'espèce à l'époque. Cette diversité semble s'être fortement réduite ensuite comme le montrent les analyses faites sur les séquences connues entre - 29 000 et - 42 000 ans, au moment où Néandertal cohabite avec l’Homo sapiens. Cette tendance vient à l'appui de la thèse du déclin démographique de Néandertal sur cette longue période conduisant, par un phénomène de goulet d'étranglement de la population, à la disparition progressive de certains génotypes, et donc à l'appauvrissement génétique de l'espèce puis à sa disparition. Reste à savoir ce qui a pu causer cette évolution (voir ci-dessous Extinction des Néandertaliens).
Indices favorables via l'ADN nucléaire
En 2006, un programme de séquençage de l'ADN nucléaire de l'homme de Néandertal a été lancé par le Max Planck Institut en collaboration avec la société 454 Life Sciences fabriquant des séquenceurs de gènes à haut débit. L'objectif est de connaître l'étendue du lien de parenté avec l'homme moderne et d'évaluer l'interfécondité de l'homme de Néandertal et l'homme moderne. Un million de paires de bases d'ADN nucléaire ont été analysées en 2006[21]. Cette étude indique que quelques modifications de gènes ne sont présentes ni dans les os de Néandertaliens (de Vindija en Croatie), ni pour l'ancêtre commun avec le chimpanzé, l'orang-outan et le macaque rhésus. Ces modifications de gènes seraient donc typiques de l'homme actuel. Les modifications concernent la capacité de codage des protéines, le battement du flagelle du sperme (axonème), l'adhésion intercellulaire, la régulation de la transcription des ribosomes et l'épiderme.
Les résultats publiés en 2010, portant sur l'analyse de 4 milliards de paires de bases d'ADN nucléaire issus d'ossements fossiles de trois Néandertaliens, montrent que ceux-ci partagent plus de caractéristiques génétiques avec les Homo sapiens modernes eurasiatiques qu'avec ceux d'Afrique sub-saharienne[1]. Les auteurs envisagent l'hypothèse selon laquelle les Néandertaliens auraient contribué au génome des populations d'humains modernes non africaines.
Cette hypothèse est la façon la plus simple d’expliquer leurs données, cependant d’autres scénarios sont possibles : en particulier, on peut faire l’hypothèse que, par suite de l’existence d’une structuration des populations au sein du continent africain, la population ancestrale des non-africains était plus proche des néandertaliens que ne l’était la population ancestrale des africains contemporains. On peut encore supposer qu’après la divergence des néandertaliens, une homogénéisation incomplète s’est réalisée entre les ancêtres des africains et ceux des non-africains, ce qui permettrait d’expliquer les observations réalisées [réf. nécessaire].
Histoire et répartition des Néandertaliens[18],[22]
L'homme de Néandertal est un représentant du genre Homo dont l'apparition et l'évolution sont liées au continent européen, même si des Néandertaliens ont émigré par la suite au Proche-Orient, sur les territoires actuels de l'Irak, de la Syrie et d'Israël, ainsi qu’en Asie centrale (Techik-Tach, en Ouzbékistan) et en Sibérie. En 2007, la répartition géographique des Néandertaliens est de nouveau précisée : leur expansion est repoussée de 2 000 km vers l'est par rapport au site de Techik-Tach, le plus oriental connu jusqu'alors. Des fragments osseux de la grotte Okladnikov, dans l'Altaï, jusqu'alors mal référencés, sont attribués à des Néandertaliens après une analyse génétique de leur ADN mitochondrial par l'Institut Max Planck d'anthropologie évolutive de Leipzig. D'après les chercheurs, l'ADN mitochondrial des Néandertaliens de l'Altaï est d'ailleurs plus proche de celui des Néandertaliens de la grotte Scladina, en Belgique, que de celui de l'Ouzbékistan, suggérant plusieurs vagues de migrations et de peuplements de la région. L'équipe du généticien Svante Pääbo suggère que la présence de Néandertaliens dans l'Altaï rend envisageable une extension plus orientale, en Mongolie voire en Chine[23],[24].
Il y a plus d'un million d'années, quelques groupes humains seraient arrivés par vagues successives en Europe, lors de brefs passages ayant laissé peu de traces sous forme de galets taillés. Les plus anciens fossiles humains européens datent de 1,8 million d'années et ont été mis au jour à l'Est du continent, à Dmanissi en Géorgie. Leur anatomie, intermédiaire entre celles d'Homo habilis et Homo erectus, a conduit à la définition d'une nouvelle espèce nommée Homo georgicus. À partir de 400 000 ans avant le présent, le peuplement de l'Europe s'intensifie, avec sans doute l'arrivée de l'ancêtre de l'homme de Néandertal. Les plus anciens pré-Néandertaliens dateraient ainsi de 500 à 350 000 ans.
L’évolution qui conduit à l’apparition d’Homo neanderthalensis, parfois appelée « néandertalisation », est un processus lent et progressif. Elle a sans doute comme origine des groupes humains isolés en Europe, qu’il s’agisse d’Homo erectus, d’Homo georgicus ou d’Homo antecessor. Elle peut être suivie depuis différents fossiles, parfois qualifiés d'« anténéandertaliens » ou, mieux, de « pré-Néandertaliens » jusqu’aux Néandertaliens récents.
Les pré-Néandertaliens anciens
La première étape correspond à des fossiles généralement attribués à Homo heidelbergensis : c’est le cas de l’Homme de Tautavel (- 400 000 ans), trouvé dans les Corbières en France, de la mandibule de Mauer (- 600 000 ans), trouvée près de Heidelberg en Allemagne, ou du crâne de Petralona (Grèce).
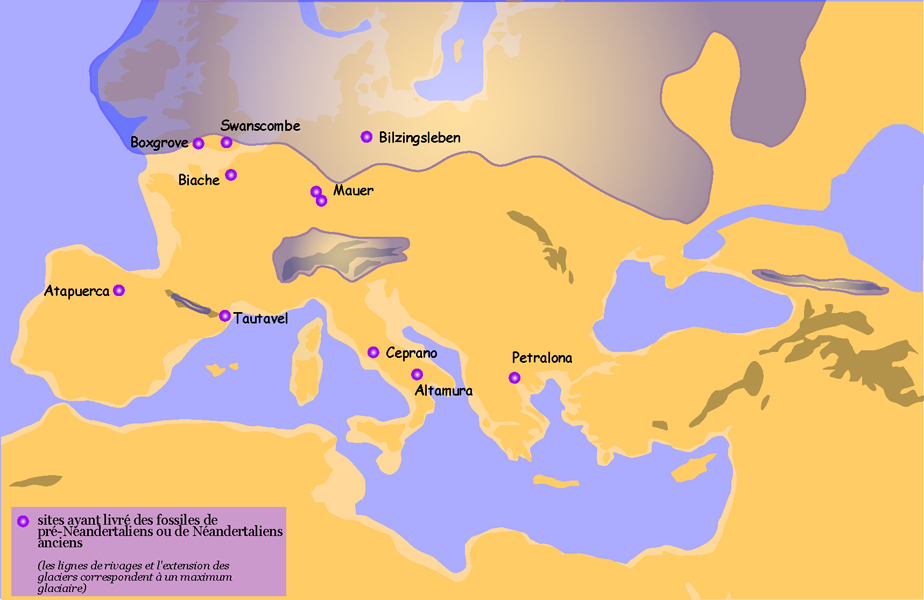
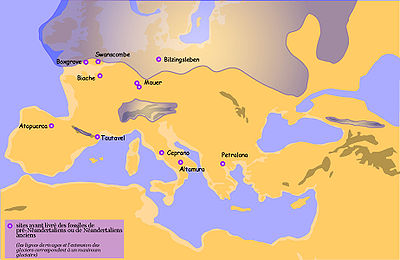 Carte de répartition des principaux sites ayant livré des restes de pré-Néandertaliens ou de Néandertaliens anciens
Carte de répartition des principaux sites ayant livré des restes de pré-Néandertaliens ou de Néandertaliens anciensLes pré-Néandertaliens récents
La deuxième étape est marquée par les fossiles de Swanscombe (Angleterre), de Steinheim (Allemagne) ou de la Sima de los Huesos à Atapuerca (Espagne).
Les Néandertaliens anciens
Les plus anciens fossiles indéniablement néandertaliens ont des âges compris entre - 250 000 et - 110 000 ans. Parmi eux, on peut citer le crâne de Biache-Saint-Vaast (Pas-de-Calais), vieux de 180 000 ans[25], les restes de La Chaise à Vouthon (Charente), la mandibule de Montmaurin (Haute-Garonne), les crânes de Saccopastore près de Rome en Italie ou les nombreux restes de Krapina en Croatie.
Les Néandertaliens classiques
Les Néandertaliens les plus typiques, dont les caractères dérivés sont les plus marqués, ont des âges compris entre - 100 000 ans et - 28 000 ans, date de leur disparition.
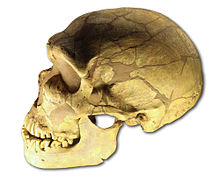 Crâne d'Homo neanderthalensis de La Ferrassie
Crâne d'Homo neanderthalensis de La FerrassieParmi les fossiles de Néandertaliens classiques, outre les vestiges de Néandertal même (env. 42 000 ans), il faut mentionner les squelettes de La Chapelle-aux-Saints, du Moustier, de La Ferrassie[26], de La Quina, de Saint-Césaire dans le Sud-Ouest de la France ou de Spy en Belgique pour ne citer que les plus complets.
Les derniers Néandertaliens connus ont été découverts notamment au Portugal, en Espagne (Zafarraya, -30 000 ans[27],[28]), en Croatie (Vindija, - 32 000 ans[29],[30]) et dans le Nord-Ouest du Caucase (Mezmaiskaya, - 29 000 ans). Toutes ces dates sont toutefois à considérer avec précaution, des réévaluations successives ayant tendance à les vieillir.
Des recherches conduites de 1999 à 2005 dans la grotte de Gorham à Gibraltar suggèrent que les Néandertaliens y ont vécu jusqu'à - 28 000 ans, voire - 24 000 ans[31],[32]. Ils auraient donc longuement cohabité avec les Hommes anatomiquement modernes, présents dans la région depuis 32 000 ans. Ces résultats sont toutefois fortement critiqués, par exemple par Joao Zilhao, de l'université de Bristol.
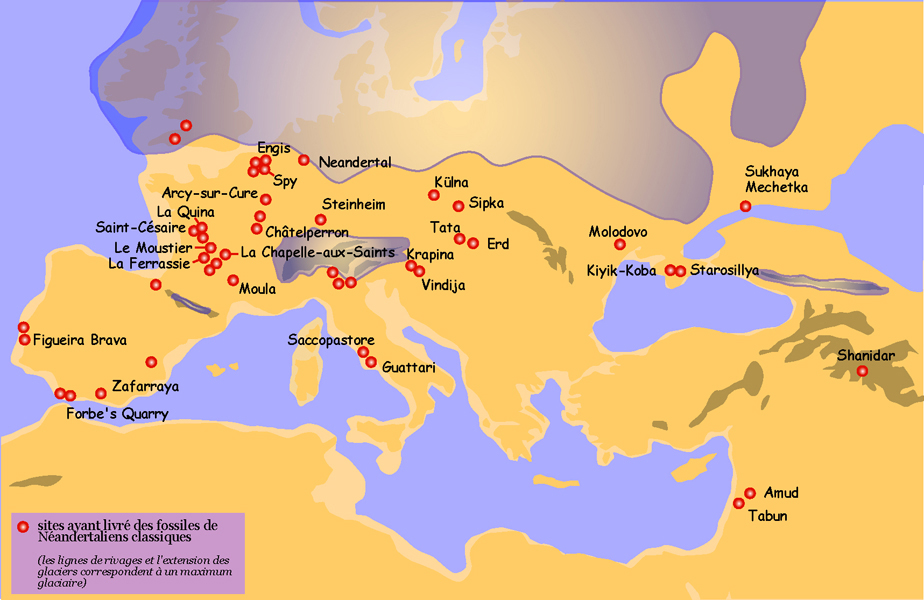
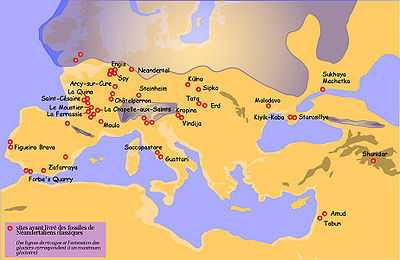 Carte de répartition des principaux sites ayant livré des restes de Néandertaliens classiques
Carte de répartition des principaux sites ayant livré des restes de Néandertaliens classiquesCaractéristiques physiques[18]
Squelette
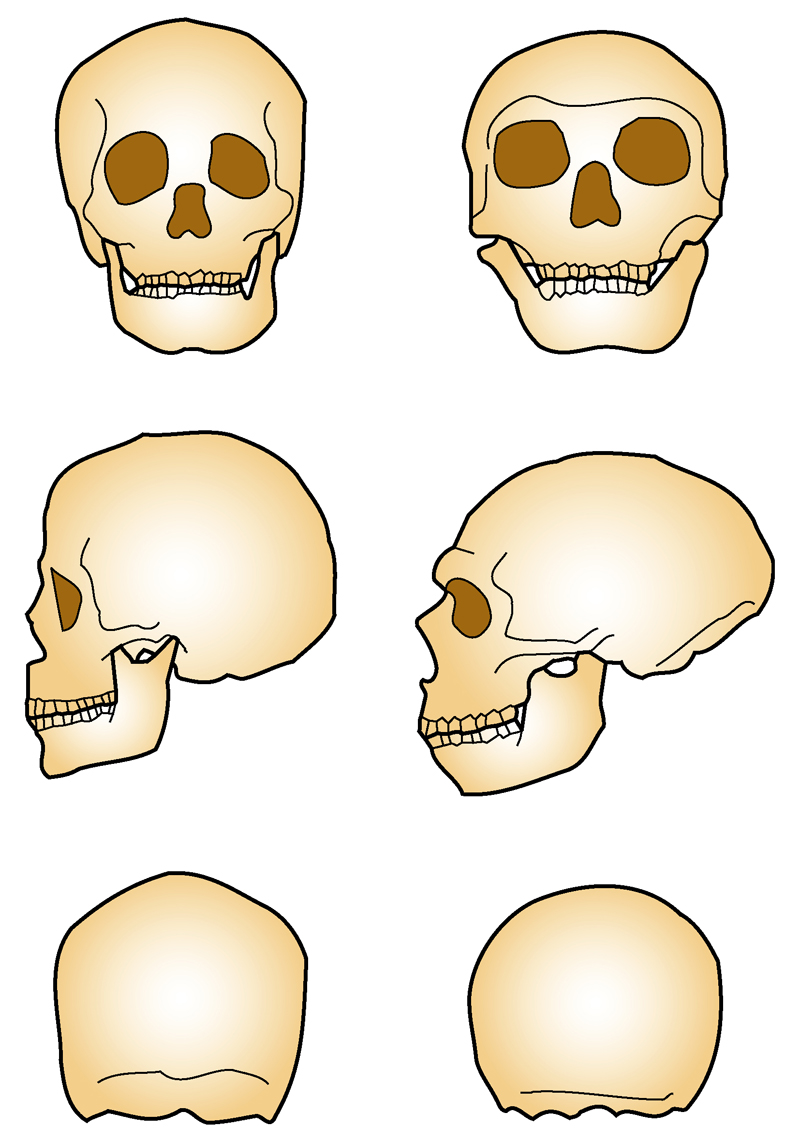
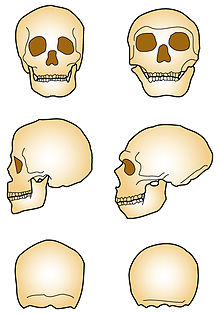 Comparaison des crânes d’Homo sapiens (à gauche) et d’Homo neanderthalensis (à droite)
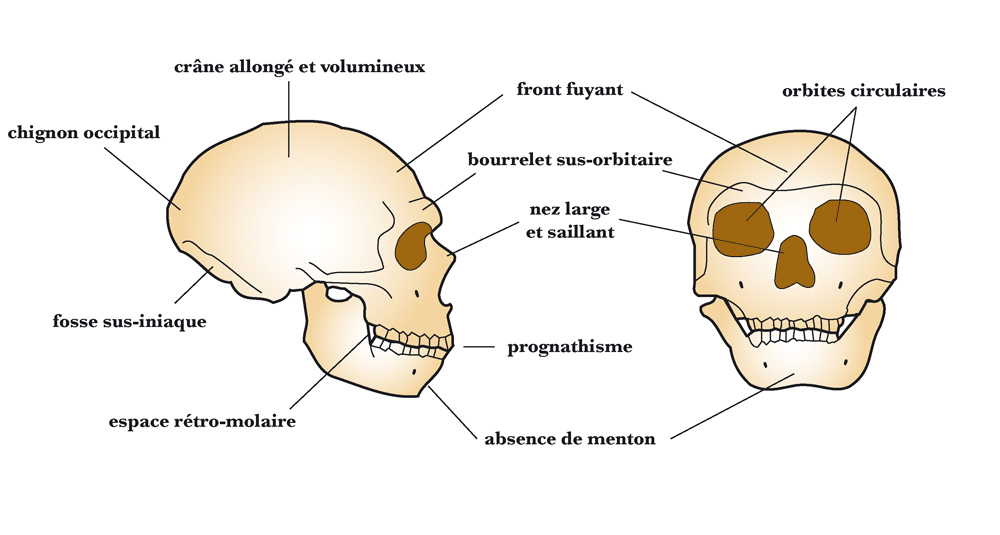
Comparaison des crânes d’Homo sapiens (à gauche) et d’Homo neanderthalensis (à droite)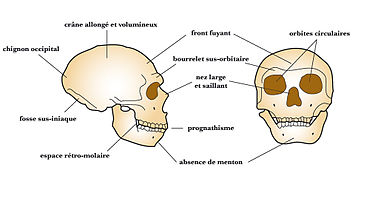 Principales caractéristiques du crâne d'Homo neanderthalensis
Principales caractéristiques du crâne d'Homo neanderthalensisLes Néandertaliens sont de corpulence souvent très massive et robuste : 90 kg et 1,65 m en moyenne pour les hommes et 70 kg et 1,55 m pour les femmes (des individus auraient atteint 1,90 m). L'ensemble de leur structure (os épais avec corticale développée) et leurs attaches musculaires laissent supposer une grande force physique.
Les Néandertaliens présentent quelques caractères archaïques, hérités de leur prédécesseur (caractères plésiomorphes), ainsi que des caractères évolués (caractères apomorphes). Les caractères évolués peuvent être partagés avec les Homo sapiens (caractères synapomorphes) ou bien être des caractères dérivés spécifiques (caractères autapomorphes). Seuls ces derniers permettent d'identifier l'espèce lors de l'examen d'un fossile.
- Les caractères plésiomorphes des Néandertaliens sont :
- la présence d'un épaississement osseux au-dessus des orbites : ce bourrelet sus-orbitaire continu (chez les prédecesseurs des Néandertaliens, ce bourrelet présente un sillon médian, le sulcus supratoralis) forme une visière ;
- un front fuyant ;
- l'absence de menton.
- Les caractères synapomorphes des Néandertaliens sont :
- un cerveau volumineux (leur capacité crânienne moyenne est légèrement supérieure (1 500 cm³) à celle de l'humain moderne) ;
- des molaires de dimensions réduites, comme chez Homo sapiens.
- Les caractères autapomorphes des Néandertaliens sont :
- une face allongée, de forme particulière avec un fort prognathisme;
- des orbites hautes et arrondies ;
- une vaste cavité nasale ;
- une arcade dentaire et un nez avancés ;
- des pommettes en retrait ;
- le trou auditif externe est ovalaire et se place dans le prolongement de la racine de l'arcade zygomatique (en dessous chez Homo sapiens). Il est fermé par un tympanal à deux versants.
- la présence d'un espace séparant les dents du fond de la branche montante de la mandibule, dit « espace rétro-molaire » ;
- un crâne dolichocéphale au profil circulaire en vue postérieure (alors que le crâne de tous les autres Hominidés présente un profil pentagonal), l'arrière crâne en forme de « ballon de rugby » (forme de « ballon de football » chez Homo sapiens) ;
- un os occipital formant une sorte de chignon et présentant une fosse en son centre, dite fosse sus-iniaque.
Les traits spécifiques aux Néandertaliens ont souvent été présentés comme des adaptations au froid. Cela est vrai en partie et les membres courts et robustes des Néandertaliens trouvent des analogues modernes dans les populations vivant dans les régions proches du pôle. Mais le climat ne suffit pas à expliquer cette évolution et des facteurs écologiques tels que l'isolation de la population et le faible brassage génétique liés aux avancées glaciaires ont également dû intervenir.
Pigmentation et couleur des cheveux
La rousseur des Néandertaliens a été évoquée à la suite de différentes études mais il semble s'agir de sur-interprétations de résultats scientifiques de la part des médias. Chez les humains, la rousseur est liée à une mutation du gène MC1R (melanocortin-1 receptor) qui régule la production de mélanine[33]. En 2000, une étude a montré que cette mutation pouvait exister depuis 100 000 ans[34]. Certains en ont conclu qu'elle pouvait être apparue chez les Néandertaliens qui l'auraient transmise aux hommes modernes[35], ce que nuancent les auteurs[36].
En octobre 2007, un article de la revue Science présente les résultats d'une étude portant sur l'extraction d'ADN fossile de deux spécimens néandertaliens, l'un découvert en Italie (Monti Lessini), l'autre en Espagne (El Sidrón 1252)[37]. Les auteurs ont amplifié et séquencé un fragment du gène MC1R et ont mis en évidence chez les deux individus une mutation inconnue chez l'homme moderne. Toutefois, il est impossible de déterminer si cette mutation était présente sur les deux allèles et donc si elle affectait le phénotype des individus en question. Pour les auteurs, la présence de ces mutations permet d'estimer qu'un pour cent environ des Néandertaliens avait une pigmentation réduite se traduisant par une peau claire et des cheveux roux[38]. Alors que cette proportion est très limitée, certains médias ont rapporté que les Néandertaliens étaient roux[39],[40].
Paléopathologie
Les restes osseux de Néandertaliens, tant en Europe qu'en Asie occidentale, présentent parfois des anomalies qui renseignent sur les lésions organiques survenues de leur vivant et parfois responsables de leur décès. Ces anomalies peuvent être classées en quatre catégories principales[41] :
Traumatologie
Article connexe : Pathologie traumatique.Les Néandertaliens semblent avoir souffert fréquemment de fractures, en particulier au niveau des côtes (Shanidar IV, « vieillard » de La Chapelle-aux-Saints), du fémur (La Ferrassie 1), du péroné (La Ferrassie 2 and Tabun 1), de la colonne vertébrale (Kébara 2[42]) et du crâne (Shanidar I, Krapina, Šaľa 1). Ces fractures sont souvent ressoudées et ne montrent pas ou peu de signes d'infection, ce qui suggère que les individus étaient pris en charge au cours de leur période d'invalidité.
En relation avec des fractures, d'autres traces de traumatismes ont été signalés sur de nombreux squelettes de Néandertaliens. Ils semblent liés à des blessures perforantes, comme chez Shanidar III dont le poumon fut certainement perforé par une blessure entre les côtes 8 et 9. Il peut s'agir d'une attaque intentionnelle ou d'un accident de chasse, mais l'individu survécut à sa blessure durant quelques semaines avant d'être tué par la chute d'un bloc rocheux dans la grotte de Shanidar. D'autres traumatismes correspondent à des coups portés à la tête (Shanidar I et IV, Krapina), tous consolidés[43],[44].
Pathologie articulaire
L'arthrite est particulièrement répandue chez les Néandertaliens les plus âgés. Elle concerne de façon spécifique les articulations comme les chevilles (Shanidar III), la colonne vertébrale et les hanches (« vieillard » de La Chapelle-aux-Saints[45],[46]), les bras (La Quina 5, Krapina, Feldhofer), les genoux, les doigts et les orteils. Le tout en relation étroite avec les maladies articulaires dégénératives (arthrose), qui peuvent aller de la dégénérescence normale, liée à l’usure, jusqu’à la restriction des mouvements, douloureuse et handicapante, et à la déformation. C’est ce qu’on observe à des degrés divers sur les squelettes de Shanidar (I-IV).
Pathologie dentaire
L'hypoplasie de l'émail dentaire est l'indicateur d'un stress survenu durant le développement des dents. Les striations et les cannelures de l'émail reflètent les périodes de pénurie alimentaire, les traumatismes ou les maladies. Une étude de 669 couronnes dentaires de Néandertaliens a montré des signes d'hypoplasie plus ou moins prononcés sur 75 % d'entre elles[47]. Les carences alimentaires en étaient la cause principale, pouvant aller jusqu'à entraîner la perte des dents. Les dents appartenant aux squelettes les plus âgés présentaient toutes une hypoplasie, particulièrement nette chez le « vieillard[48] » de La Chapelle-aux-Saints et l'individu 1 de La Ferrassie.
Pathologie infectieuse
On trouve occasionnellement sur des squelettes de Néandertaliens des lésions osseuses secondaires liées à une infection des tissus mous du voisinage. Shanidar I présente des traces manifestes de lésions dégénératives de même que La Ferrassie 1, où les lésions sur les deux fémurs, les tibias et les fibules indiquent une infection systémique ou peut-être un cancer.
Langage[49],[50]
L'aptitude physique au langage articulé des Néandertaliens a longtemps été controversée. Les discussions portent sur l'aptitude physique des Néandertaliens au langage, pour laquelle la morphologie de l'os hyoïde est importante. L'os hyoïde est un petit os qui maintient la base de la langue. Il est présent chez tous les mammifères mais, au sein de la lignée humaine, sa morphologie est déterminante pour l'aptitude à l'élocution. Très peu d'os hyoïdes de Néandertaliens ont été mis au jour : un premier a été découvert en 1983 à Kébara, sur le Mont Carmel en Israël (60 ka) et un autre dans le site d’El Sidron en Espagne (43 ka). Les deux os sont très peu différents de ceux des humains actuels[51]. Des os hyoïdes appartenant à des pré-néandertaliens ont été découverts dans le site de la Sima de los Huesos à Atapuerca en Espagne (au moins 530 ka) ; ils ont également des caractéristiques proches de celui des Homo sapiens[52].
Même en faisant abstraction de cet argument de poids, de nombreux chercheurs considèrent que la complexité de l'outillage moustérien attribué à l'Homme de Néandertal est une preuve indirecte de ses capacités cognitives, incluant une forme de langage articulé.
Concernant le conduit vocal des Néandertaliens, Philip Lieberman maintient depuis 1971 que ceux-ci ne disposaient pas d'un pharynx de taille suffisante pour produire tous les sons que l'on observe dans les langues du monde. Malgré de nombreuses critiques concernant cette argumentation, cette théorie s'est largement diffusée pendant une trentaine d'années. À la suite d'une longue controverse[53],[54],[55], il semble que les arguments avancés par Lieberman ne sont plus tenables. La reconstruction anatomique du conduit vocal qu'il avait utilisée n'était pas réaliste et ses simulations peu convaincantes. Ce n'est pas la taille du pharynx qui permet de parler mais le contrôle des articulateurs (cordes vocales, langue, mandibule, voile du palais, lèvres). Les nouvelles simulations montrent bien que les Néandertaliens avaient la capacité physique de parler.
Une étude publiée en 2007[56] et portant sur l'analyse de l'ADN provenant des restes de deux Néandertaliens découverts à El Sidrón (Espagne) aurait permis d'y détecter la même version du gène FOXP2 (en) (forkhead box P2) que celle présente chez les hommes modernes[57]. Cela pourrait plaider en faveur de l'aptitude des Néandertaliens au langage puisqu'on estime que ce gène joue un rôle important dans le développement des parties du cerveau liées à la maîtrise du langage articulé.
Culture et techniques
 Nucléus et éclat Levallois en silex de Haute-Saône
Nucléus et éclat Levallois en silex de Haute-SaôneAprès avoir longtemps été considéré comme un être archaïque et encore proche de l'animalité, y compris par une partie de la communauté scientifique, l'Homme de Néandertal commence à apparaître comme un être doté de capacités intellectuelles et de traditions culturelles.
Le tableau ci-dessous liste un certain nombre de comportements considérés comme modernes et courants chez Homo sapiens ainsi que la fréquence relative de leur mise en œuvre par Homo neanderthalensis[58] :
Comportement Fréquent Occasionnel Absent Incertain Art √ Utilisation de pigments √ Fabrication de parures √ Sépultures √ Rites funéraires √ Échanges à longue distance √ Production de microlithes √ Production de pointes barbelées √ Production d'outils en os √ Production de lames √ Utilisation d'aiguilles √ Exploitation des ressources marines √ Chasse aux oiseaux √ Artisan de la pierre[22]
Articles détaillés : Moustérien et Châtelperronien.Il est l'auteur d'un outillage complexe et élaboré, et notamment des industries du Moustérien. Ses méthodes de débitage apportent en outre la preuve de ses capacités d'abstraction et d'anticipation, en particulier en ce qui concerne le débitage Levallois. Les éclats obtenus par cette méthode ou par d'autres pouvaient être utilisés bruts ou bien retouchés, légèrement modifiés sur leurs bords pour obtenir des outils plus spécialisés tels que les racloirs ou les denticulés.
 Racloir moustérien en silex de la grotte du Noisetier
Racloir moustérien en silex de la grotte du NoisetierDes preuves directes (traces d'adhésif naturel en bitume ou en résine) ou indirectes (répartition des traces d'utilisation) montrent que certains outils étaient utilisés emmanchés. Les manches eux-mêmes, réalisés en matériaux périssables, n'ont pas été conservés. En revanche, des conditions particulièrement favorables ont permis la conservation de quelques objets en bois. Le plus spectaculaire est sans conteste un fragment d'épieu en if fiché dans le thorax d'un éléphant, mis au jour à Lehringen (Basse-Saxe). Dans le même site, ont été découverts des éclats Levallois ayant servi à découper de la peau et de la viande.
Il est probable que les derniers Néandertaliens soient les auteurs du Châtelperronien, un faciès culturel de transition entre le Paléolithique moyen et le Paléolithique supérieur en Europe occidentale. Ce faciès est caractérisé par des comportements longtemps considérés comme propres aux hommes modernes : débitage de lames, utilisation de parure, fabrication d'outils en os, etc.[59],[60].
Un grand chasseur[22]
En 1981, l'archéologue Lewis Binford soutient une théorie selon laquelle les premiers hominidés (dont les Néandertaliens) jusqu'au Paléolithique moyen ne pratiquaient que la cueillette ou le charognage passif, seuls les hommes modernes pratiquant la chasse de grand gibier rapide[61]. Cette théorie est aujourd'hui abandonnée.
Rares sont les preuves directes de la pratique de chasse aux grands herbivores par les Néandertaliens telles que le fragment d'épieu en if de Lehringen ou des pointes emmanchées en silex ou en os. En revanche, les sites livrent des accumulations impressionnantes d'ossements de grands mammifères (Bison à Coudoulous, Lot et à Mauran, Haute-Garonne ; Saïga en Crimée ; Bouquetin dans la grotte du Lazaret ; Aurochs à La Borde, Lot ; Cheval à Saint-Césaire, Charente-Maritime) interprétées comme le résultat de chasses saisonnières, parfois avec utilisation d'avens ou de fondrière comme pièges naturels. De plus, les analyses biogéochimiques sur le collagène osseux des Néandertaliens et des mammifères associés[62] montrent une alimentation carnée proche de celle du loup, même en période tempérée : chasseurs de grands mammifères, les Néandertaliens avaient des stratégies (communautés spécialisées dans la chasse de deux ou trois espèces, technique à l'approche ou à la poursuite de proies en fonction de l'âge et du sexe) qui attestent d'une parfaite connaissance de l'environnement, de l'éco-éthologie des animaux et d'un savoir-faire technique. Enfin les études anatomiques montrent leur adaptation à la chasse : ils étaient en effet trapus et musclés, pesant en moyenne 90 kg pour 1,65 m pour les hommes, 70 kg pour 1,55 m pour les femmes[63].
La pratique ponctuelle d'un charognage actif (accès primaire à la carcasse en écartant les prédateurs - hyène des cavernes, loup, lion des cavernes - ou en recherchant les animaux morts dans des pièges naturels) a également été évoquée, notamment pour les grands mammifères (mammouth, rhinocéros laineux)[64],[65].
Alimentation
De précédentes recherches considéraient que les Néandertaliens étaient exclusivement des chasseurs carnivores faisant partie des superprédateurs (chassant essentiellement de grands herbivores alors qu’Homo sapiens avait diversifié son alimentation - petit gibier, fruits de mer, poissons d'eau douce et de mer), ce régime alimentaire différent étant considéré comme une des causes de leur extinction[66].
En 2010, des analyses ont porté sur des phytolithes piégés dans des plaques de tartre de dents fossilisées néandertaliennes provenant de différents sites. Elles ont révélé des traces de plantes fossilisées (palmier-dattier, légumineuses, rhizomes de nénuphar, graminées du genre Triticum ou Hordeum), indiquant un régime alimentaire lui aussi diversifié et un comportement de chasseur-cueilleur. De plus, certains grains d'amidon retrouvés montrent des processus de cuisson, suggérant que les Néandertaliens, grâce à leur maîtrise du feu, cuisaient ces végétaux en les faisant bouillir. En effet, on pensait auparavant que seules les viandes étaient cuites (d'après l'analyse des ossements d’animaux retrouvés dans de nombreux foyers) alors que les végétaux étaient simplement grillés[67]. Les Néandertaliens ont ainsi une alimentation carnée composée de grands mammifères mais incluant également des végétaux ou de petits animaux (lagomorphes, oiseaux, mollusques terrestres[68]) lorsque les conditions s'y prêtent.
La découverte de restes de coquillages dans la grotte de Bajondillo, dans le sud de l'Espagne, indique que des Néandertaliens mangeaient des fruits de mer en Europe dès 150 000 ans avant le présent[69],[70].
Des recherches ont également montré que les Néandertaliens chassaient des animaux marins tels que les dauphins ou les phoques. Globalement, les découvertes montrent de plus en plus que leur alimentation était semblable à celle des humains anatomiquement modernes [71],[72].
Sépultures[73]
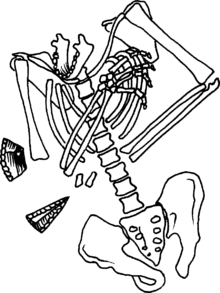 L'homme de Néandertal surnommé Moshe mis au jour dans la grotte de Kébara
L'homme de Néandertal surnommé Moshe mis au jour dans la grotte de KébaraS'il est possible qu’Homo heidelbergensis, l'un des ancêtres probables de l'Homme de Néandertal, ait adopté un comportement particulier vis-à-vis de ses morts à Atapuerca, les premières véritables sépultures connues sont néandertaliennes. Les plus anciennes datent d'environ - 100 000 ans et ont été mises au jour au Proche-Orient. Elles se multiplient ensuite et on en trouve en France (La Chapelle-aux-Saints, La Ferrassie, La Quina, Le Moustier, Saint-Césaire), en Belgique (Spy), en Israël (Kébara, Amud), au Kurdistan irakien (Shanidar), en Ouzbékistan (Teshik-Tash). Dans certains cas, elles comprennent des dépôts funéraires (outils lithiques, fragments de faune).
L'une des sépultures de Shanidar renfermait un Néandertalien enterré sous une grande dalle. Une grande quantité de pollens de plantes à fleurs était présente autour du corps[74],[75]. Ces pollens ont longtemps été considérés comme la preuve du dépôt de nombreuses fleurs lors de l'enfouissement. Cette interprétation est aujourd'hui remise en question, des phénomènes post-dépositionnels ou l'action de rongeurs ayant pu contribuer à l'accumulation de pollens[76].
Ces sépultures comportent souvent des fosses intentionnelles et sont pratiquement toujours associées à des habitats. Il est peu probable qu'elles n'aient eu qu'un rôle fonctionnel simplement destiné à se débarrasser d'une dépouille, même si leur interprétation en termes de religiosité est sujette à discussion.
Les Néandertaliens ont plusieurs types de rites funéraires : sépultures, inhumations en deux temps après décomposition du corps à l'air libre (traces de désarticulation et décharnement à Shanidar VI et VIII, Krapina, Kébara[77], combe Grenal[78]), « culture des ancêtres » (analyses de taphonomie sur des crânes isolés et corps sans tête à Kébara), endocannibalisme (Moula-Guercy, Vindija et Krapina en Croatie : stries de fracturation sur os frais)[79].
Culte de l'ours
Article détaillé : Culte de l'ours.Dans certains sites tels que le Regourdou en Dordogne, des accumulations de crânes d'ours qui semblaient disposés intentionnellement ont été interprétées comme le résultat d'un « culte de l'ours ».
Au Regourdou, un squelette d'ours brun reposait sous une dalle monolithe d'un poids de 850 kg, dans une fosse peu profonde. À proximité, le corps d'un Néandertalien était couché sur le côté gauche, la tête vers le nord, en position fœtale. Le crâne manquait, mais il restait la mandibule. L’absence du crâne a également été observée dans le cas de la sépulture néandertalienne de Kébara. D'après E. Bonifay, il s'agissait d'une véritable tombe composée d’une fosse dallée, empierrée et couverte de sable et de cendres de foyer[80],[81]. Cette interprétation a largement été remise en question depuis, les accumulations d'ossements d'ours du Regourdou pouvant être liés à des phénomènes taphonomiques liés à l'occupation de la cavité par des ours hibernants[82].
Plus généralement, l'existence du culte de l'ours, évoquée récemment par l'écrivain Jean M. Auel, est aujourd'hui contestée par de nombreux scientifiques. Les crânes d'ours sont extrêmement résistants et peuvent être déplacés par des phénomènes naturels jusqu'à acquérir des positions évoquant une organisation volontaire mais en fait seulement due au hasard[83].
Cannibalisme
Article détaillé : Anthropophagie.La présence de traces de désarticulation, de décharnement, de fracturation intentionnelle ou de calcination sur certains os de Néandertaliens a été interprétée comme un témoignage de la pratique du cannibalisme. Des ossements de sites tels que l'Abri Moula[84], en Ardèche ou Krapina[85],[86],[87] en Croatie présentent de telles traces de découpe. Il est toutefois difficile de démontrer s'il s'agit de cannibalisme plutôt que d'un traitement post mortem des dépouilles dans le cadre d'un rite funéraire. Les fragments d'os de Krapina présentent des marques comparables à celles de sépultures secondaires d'une nécropole du XIVe siècle découverte au Michigan, correspondant à l'ablation de la chair sur une dépouille partiellement décomposée. Certains os crâniens du site des Pradelles à Marillac-le-Franc présentent des traces de découpe correspondant sans doute au prélèvement du cuir chevelu par scalpation.
Le crâne de Néandertalien découvert dans la Grotte Guattari (Mont Circé, Italie) a longtemps été considéré comme une preuve irréfutable de rituel anthropophagique : il aurait été déposé dans un cercle de pierre après que le trou occipital avait été élargi pour consommer le cerveau. Des examens approfondis ont montré que le cercle de pierre était probablement naturel et que l'élargissement du trou occipital avait été causé par une hyène, ce que confirme la présence de traces de dents en différents points du crâne[88],[89].
La naissance de l'art
 Relevé du motif en zigzag gravé sur un os découvert dans le site paléolithique moyen de Bacho Kiro en Bulgarie
Relevé du motif en zigzag gravé sur un os découvert dans le site paléolithique moyen de Bacho Kiro en BulgarieAu Paléolithique moyen apparaissent également les premières manifestations de préoccupations esthétiques ou symboliques :
- collecte de fossiles ou de minéraux rares ;
- utilisation d'ocre (même si dans certains cas des utilisations fonctionnelles peuvent être évoquées)[90] ;
- utilisation de plumes comme le montre la disposition des traces de découpe sur les ossements d'oiseau de la grotte de Fumane en Italie[91] ;
- gravure de traits, de lignes ou de signes géométriques simples sur des os ou des pierres[92],[22].
Cette reconnaissance de la culture néandertalienne remet en cause la primauté culturelle de l'homme moderne : alors que l'on pensait il y a peu que la culture technique et symbolique des Néandertaliens était très nettement inférieure quantitativement et qualitativement à celle de l'Homo sapiens, les découvertes récentes font apparaître que l'Homme de Néandertal avait lui aussi développé certaines techniques évoluées (débitage de lames[93]) ou adopté des traits culturels modernes (sépultures, signes gravés). La thèse du rôle capital de l'arrivée de l’Homo sapiens en Europe et celle d'une corrélation entre l'évolution biologique et l'évolution culturelle expliquant le plus grand développement de l’Homo sapiens par son évolution biologique s'en trouvent donc remises en question...
Extinction
Les Néandertaliens disparaissent il y a environ 29 000 ans, disparition encore en partie inexpliquée et suscitant de nombreuses hypothèses[94],[95],[96], faisant parfois intervenir des modèles mathématiques[97] ou économiques[98] plus ou moins insolites. Les données archéologiques montrent qu'il n'y a pas eu une extinction massive mais au contraire une disparition progressive.
La disparition des Néandertaliens coïncide apparemment avec l'arrivée de groupes d'Hommes anatomiquement modernes ayant quitté le Proche-Orient pour l'Europe, il y a environ 40 000 ans, sans doute à la faveur d'un épisode climatique tempéré de la dernière glaciation. Ces hommes modernes, parfois appelés « Hommes de Cro-Magnon », sont porteurs d'une nouvelle culture matérielle, appelée Aurignacien et caractérisée par la généralisation du débitage laminaire et lamellaire, l'utilisation du percuteur tendre pour ces débitages, la fabrication d'outils en matières dures animales (notamment des pointes de sagaies en os). Les hommes de l'Aurignacien sont également les auteurs des plus anciennes œuvres de l'art pariétal et mobilier d'Europe.
Les Hommes de Néandertal et les Hommes modernes ont probablement cohabité ainsi pendant quelques millénaires, quoiqu'aucune trace directe d'interaction n'ait été découverte.
Hypothèses abandonnées ou peu vraisemblables
On ne cite même plus l'explication suivant laquelle l'homme moderne serait tout simplement le descendant de l'homme de Néandertal. Cette théorie était pourtant, voici moins d'un demi-siècle, défendue encore avec passion par Camille Arambourg, membre de l'Institut et directeur de l'Institut de paléontologie humaine de Paris, aux yeux duquel ceux qui ne reconnaissaient pas l'évidence de cette solution obéissaient manifestement à d'obscures considérations métaphysiques : « La naïve et pitoyable vanité humaine se refuse à admettre que le "roi de la création" ait pour ancêtre le Néanderthalien, si proche encore de l'animalité, en lequel on voudrait s'efforcer de voir un rameau "éteint" ou même "dégénéré", sans rapport avec l'Homo sapiens[99]. »
D'autres hypothèses ont encore quelques partisans :
- Disparition liée aux difficultés d'adaptation des Néandertaliens face à un changement environnemental (disparition du gibier, modifications climatiques, etc.)
- Intoxication alimentaire par des produits toxiques (champignons vénéneux ou levures, sels métalliques de plomb, de cuivre, d'arsenic…).
- Problèmes d'ordre génétique liés à une forte consanguinité et/ou des mutations spontanées ayant entraîné des maladies congénitales telles que l'hémophilie, le diabète insulino-dépendant ou une forme de stérilité, ayant suffisamment affecté la démographie de la population pour la faire disparaître.
- Carence alimentaire notamment en vitamine (A, B6 ou B12, E, F, etc.) ou en sels minéraux essentiels (iodure, iodate, ferrique…), due à un régime alimentaire essentiellement carné, ayant entraîné progressivement un taux de mortalité très élevé non compensable par la reproduction.
L'ensemble de ces hypothèses est peu probable à l'échelle d'une population continentale, et difficiles à concilier avec la longue histoire des Néandertaliens (près de 200 000 ans) et leurs capacités d'adaptation à des conditions climatiques et environnementales variées et fluctuantes[100].
- Décalage entre les dynamiques démographiques respectives des deux populations, les Néandertaliennes ayant légèrement moins d'enfants que les femelles Homo sapiens : les facteurs démographiques sont néanmoins très difficiles à appréhender pour des populations préhistoriques. Par ailleurs, cette hypothèse est finalement celle d'une concurrence avec l'espèce Homo sapiens.
Hypothèses en cours d'examen
Extinction d'origine infectieuse
Épidémie liée à une infection virale (fièvre hémorragique) ou bactérienne (équivalent de lèpre, de peste…), ou parasitaire (paludisme) ayant décimé préférentiellement la population néandertalienne : cette hypothèse est plausible dans la mesure où les groupes d'hommes modernes ont pu apporter des maladies épidémiques d'origine tropicale auxquelles ils étaient résistants, mais le temps de cohabitation (12000 ans environ) semble trop long pour une épidémie.
Extinction par stérilité de l'hybridation
Disparition progressive de la population néandertalienne liée à la possibilité d'accouplements féconds mais donnant des hybrides stériles, au moins chez les Néandertaliennes : hypothèse émise par le paléontologue finlandais Björn Kurtén[101], difficile à tester.
Extinction violente liée à Homo sapiens
Disparition des Néandertaliens liée à l'arrivée des hommes modernes et à la compétition territoriale pour l'exploitation des ressources[100], voire leur élimination physique par les hommes modernes, par conflits violents sur les zones de contact et réduction des territoires néandertaliens : l'hypothèse s'appuie en particulier sur la concordance des périodes d'expansion d’Homo sapiens avec cette disparition[102]. Elle est contestée du fait de l'absence de traces de morts violentes ou de traces de cohabitation prolongée sur un même territoire[103], ou bien en avançant que les deux groupes ne devaient pas occuper l'ensemble du territoire européen et que les Néandertaliens avaient une meilleure connaissance de ce territoire et de ses ressources que les nouveaux arrivants[104].
Fusion génétique partielle avec Homo sapiens
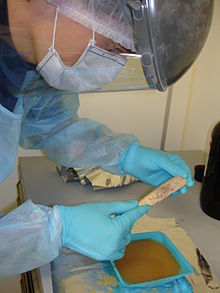 Chercheur de l'Institut Max-Planck préparant un échantillon d'os pour en extraire l'ADN.
Chercheur de l'Institut Max-Planck préparant un échantillon d'os pour en extraire l'ADN.La dissolution de la population néandertalienne dans la population Homo sapiens par accouplement fécond est l'une des hypothèses envisagées pour expliquer sa disparition.
Les données du débat
Un squelette d'enfant découvert au Portugal a été présenté comme un hybride Néandertal/sapiens, impliquant que le patrimoine génétique des Néandertaliens ait pu contribuer au patrimoine génétique de l'homme actuel. Cette hypothèse est fortement discutée en raison des difficultés à démontrer la véracité de l'hybridation chez un individu juvénile, alors que la variabilité de la population concernée est mal connue[18]. Par ailleurs, les études génétiques consacrées à ce sujet aboutissaient à des résultats contradictoires : pour certains auteurs, il n'y a aucun indice de contribution des Néandertaliens à l'ADN mitochondrial des premiers hommes modernes[105] tandis que pour d'autres les gènes d'Homo sapiens ne seraient pas exclusivement d'origine africaine[106],[107] et le génome néandertalien pourrait représenter 5 % de celui des humains modernes[108]. Une étude de l'université de Chicago a permis d'identifier un gène lié à la croissance du cerveau qui aurait été transmis à l'homme par les Néandertaliens et qui est présent chez 70 % des humains actuels[109].
En 2010, séquençage d'ADN de Néandertaliens
Une équipe de l'Institut Max Planck à Leipzig[1],[110], sous la direction de Svante Pääbo, a procédé au séquençage d'ADN nucléaire d'ossements fossiles de Néandertaliens. La comparaison avec les mêmes séquences d'humains modernes montre que 1 à 4 % des polymorphismes nucléotidiques de l'ADN est commun aux Néandertaliens et aux Homo sapiens eurasiatiques mais pas africains ; ces croisements auraient pu survenir il y a 150 000 à 80 000 ans au Moyen-Orient[111]. L'une des hypothèses des auteurs, dans leur étude publiée en mai 2010, est que les Néandertaliens ont contribué au génome des populations d'humains modernes non africaines mais d'autres hypothèses expliquent également convenablement les données recueillies (cf. supra). Le décryptage de l'ADN d'un Homo sapiens archaïque (vers - 100 000 ans), en cours, permettra peut-être de trancher. En 2010, le séquençage à 63 % du génome de Néandertal a permis de distinguer 20 régions génétiques spécifiques à l'homme moderne.
L'homme de Néandertal dans les œuvres de fiction
Littérature
- The Clan of the Cave Bear, 1980, de Jean M. Auel, traduit en français, d'abord sous le titre Ayla, l'enfant de la Terre, puis plus tard, sous le titre Le Clan de l'Ours des cavernes : les aventures d’Ayla, une fillette de Cro-Magnon orpheline recueillie vers l'âge de cinq ans par une tribu de Néandertaliens.
- Neanderthal - Their time has come, 1996 de John Darnton : récit d'une rencontre entre des hommes modernes et des Néandertaliens ayant survécu jusqu'à nos jours.
- Hominids - The Neanderthal Parallax, 2002 de Robert J. Sawyer : récit d'un monde imaginaire où les rôles d'Homo sapiens et de l'homme de Néandertal sont inversés.
- L'Enfant du Temps (en), 2006 d'Isaac Asimov & Robert Silverberg : histoire d'un enfant néandertalien, ramené à l'époque contemporaine pour être étudié. Un conte scientifique et philosophique.
- Philip José Farmer, dans son cycle de romans Le Fleuve de l'éternité, met en scène un néandertalien du nom de Kazz, comme personnage de second plan.
Cinéma
- La Guerre du feu, film de Jean-Jacques Annaud, 1981, d'après le roman de J.-H. Rosny (1911).
- Humains, film avec Lorant Deutsch sorti en France le 22 avril 2009.
- Ao, le dernier Néandertal, film de Jacques Malaterre, sorti en France le 29 septembre 2010.
Notes et références
- Richard E. Green et al. (2010) « A draft sequence of the Neandertal genome », Science, Vol. 328. no. 5979, pp. 710-722.
- Science, Comparative Genomics : the Neandertal Genome
- Welcome to the family, Homo sapiens neanderthalensis, New Scientist]
- Cette durée de vie de l'espèce de 300 000 ans semble en contradiction avec une apparition bien confirmée vers 250 000 ans. En fait le moment de l'apparition de l'espèce est très incertain, de -500 000 à -200 000 ans selon les fossiles et les spécialistes qui les observent, d'où l'approximation de 300 000 ans.
- Groenen, M. Pour une histoire de la préhistoire, Éd. J. Millon, (1994) (ISBN 978-2-905614-93-3)
- Trinkaus, E. et Shipman, P. Les Hommes de Neandertal, Seuil, (1996) (ISBN 978-2-02-013178-0)
- Fuhlrott, C.J. (1859) - « Menschliche Ueberreste aus einer Felsengrotte des Düsselthals. Ein Beitrag zur Frage über die Existenz fossiler Menschen », Verhandl. Naturhist. Ver. Preuss. Rheinlande Westphalen, 16, p. 131-153.
- Fuhlrott, C.J. (1865) - Der fossile Mensch aus dem Neanderthal und sein Verhältniß zum Alter des Menschengeschlechts, Duisburg, 78 S. 2 Abb.
- Schaaffhausen, H. (1858) - « Zur Kentniss der ältesten Rassenschädel », Archiv Verdinbung Mehrereh Gelehrten, p. 453-488.
- Pêche miraculeuse en mer du Nord - Le Soir, 17 juin 2009
- King, W. (1864) - « The reputed fossil man of the Neanderthal », Quarterly Journal of Science, vol. 1, p. 88-97.
- Boule, M. (1911-1913) - « L’homme fossile de la Chapelle-aux-Saints », Annales de paléontologie, t. VI-VII-VIII.
- Krings, M., Stone, A., Schmitz, RW., Krainitzki, H., Stoneking, M., et al. (1997) « Neandertal DNA sequences and the origin of modern humans », Cell, 90, p. 19–30.
- D. Serre, A. Langaney, M. Chech, M. Teschler-Nicola, M. Paunovic, P. Mennecier, M. Hofreiter, G. Possnert, S. Pääbo, (2004), « No evidence of Neandertal mtDNA contribution to Early Modern Humans » PLoS Biology Vol. 2, No. 3
- Zilhão, J. et Trinkaus, E. (2002), Portrait of the Artist as a Child. The Gravettian Human Skeleton from the Abrigo do Lagar Velho and its Archeological Context, Trabalhos de Arqueologia, Vol. 22, 609 p.
- Plagnol V., Wall J.D., (2006), « Possible ancestral structure in human populations », PLoS Genetics, 2 : e105 ; citation : ..strong evidence for ancient admixture in both a European and a West African population (p ≈ 10−7), with contributions to the modern gene pool of at least 5%. While Neanderthals form an obvious archaic source population candidate in Europe..
- Duarte, C., Maurício, J., Pettitt, P.B., Souto, P., Trinkaus, E., van der Plicht, H. et Zilhão, J., (1999), « The early Upper Paleolithic human skeleton from the Abrigo do Lagar Velho (Portugal) and modern human emergence in Iberia », Proceedings of the National Academy of Sciences, vol. 96, n° 13, pp. 7604-7609, citation : [...] the persistence of a morphological mosaic for that time period, indicates that this child was not the result of a rare Neandertal/early modern interbreeding but the descendant of extensively admixed populations.
- Hublin, J.-J. « Origine et évolution des Néandertaliens », in Aux origines de l'humanité, vol. 1, Y. Coppens et P. Picq (dir.), Fayard, (2001)
- Ludovic Orlando, Pierre Darlu, Michel Toussaint, Dominique Bonjean, Marcel Otte and Hänni Catherine, CNRS, ENS Lyon « Revisiting neandertal diversity with a 100.000 year mtDNA sequence», Current Biology 6 juin 2006, volume 16, issue 11
- L'ADN humain le plus vieux du monde vient d'être publié
- Richard E. Green et al. (2006) « Analysis of one million base pairs of Neanderthal DNA », Nature, 444, 330-336 (16 novembre 2006).
- Jaubert, J. Chasseurs et artisans du Moustérien, La Maison des Roches (1999).
- Krause, J., Orlando, L., Serre, D., Viola, B., Prufer, K., Richards, M. P., Hublin, J.-J., Hanni, C., Derevianko, A. P. et Paabo, S. (2007) - « Neanderthals in central Asia and Siberia », Nature, 449, 7164, pp. 902-904. (résumé (en))
- « Des néandertaliens complétement à l'est ! », sur hominidés.com.
- Demoule, J.-P., « De l’archéologie de sauvetage à l’archéologie préventive », préface de La France archéologique, éditions Hazan/Inrap.
- Heim, J.L., (1976) - Les Hommes fossiles de La Ferrassie, Paris, Masson.
- Hublin J.J., Barroso Ruiz C., Medina Lara P., Fontugne M., Reyss J.-L., 1995 - The Mousterian site of Zafarraya (Andalucia, Spain): dating and implications on the palaeolithic peopling processes of Western Europe. C. R. Acad. Sc. Paris. 321 (IIa): 931-937
- Hublin J.-J et E.Trinkaus, 1998 - The Mousterian human remains from Zafarraya (Andalucia, Spain). American Journal of Physical Anthropology suppl 26 : 122-123
- Smith, F.H., Trinkaus, E. Pettitt, P.B., Karavanic', I. et Paunovic, M., (1999) « Direct radiocarbon dates for Vindija G1 and Velika Pećina Late Pleistocene hominid remains », PNAS, Vol. 96, Issue 22, 12281-12286, October 26, 1999.
- Higham, T., Ramsey, C.B., Karavanic, I., Smith, F.H. et Trinkaus, E., (2006), « Revised direct radiocarbon dating of the Vindija G1 Upper Paleolithic Neandertals », PNAS, January 17, 2006, vol. 103, no. 3, p. 553-557.
- Finlayson, C. et al. (2006) « Late survival of Neanderthals at the southernmost extreme of Europe », Nature, advanced online publication, 13 septembre 2006.
- Brill, D. (2006) « Neanderthal's last stand », news@nature.com, 13 septembre 2006.
- Rees, J. L. (2000) - « The Melanocortin 1 Receptor (MC1R): more than just red hair », Pigment Cell Res, 13, p. 135–140.
- Harding, R.M., Healy, E., Ray, A.J., Ellis, N.S., Flanagan, N., Todd, C., Dixon, C., Sajantila, A., Jackson, I.J., Birch-Machin, M.A. et Rees, J.L. (2000) - « Evidence for variable selective pressures at MC1R », American Journal of Human Genetics, 66, 4, p. 1351-1361.
- Red-Heads and Neanderthals
- Red hair genes 100,000 years old
- Lalueza-Fox, C., Römpler, H., Caramelli, D., Stäubert, C., Catalano, G., Hughes, D., Rohland, N., Pilli, E., Longo, L., Condemi, S., de la Rasilla, M., Fortea, J., Rosas, A., Stoneking, M., Schöneberg, T., Bertranpetit, J. et Hofreiter, M. (2007) - « A melanocortin 1 receptor allele suggests varying pigmentation among Neanderthals », Science, Published Online October 25, 2007. résumé
- « Certains hommes de Néandertal avaient les cheveux roux, selon leur ADN »
- « Neanderthals 'were flame-haired' »
- « L'homme de Neandertal, ce rouquin au teint pâle »
- Tropel, Ch. (2004) « Approche synthétique de la pathologie des Néanderthaliens », Paléobios, 13. texte intégral
- Bar Yosef O. et Vandermeersch B. (1991) Le squelette moustérien de Kébara 2, Cahiers de Paléoanthropologie, CNRS, 197 p.
- Trinkaus, E. (1983) The Shanidar Neandertals, New York : Academic Press, 502 p. (pour Shanidar)
- Montet-White, A. (1996) Le Paléolithique en ancienne Yougoslavie, Jérôme Millon, 268 p. (pour Krapina)
- Tappen N.C. (1985) « The dentition of the « Old man » of la Chapelle-aux-Saints and inferences concerning Neandertal behavior », Am. J. Phys. Anthropol., 67 : p. 43-50.
- Trinkaus E. (1985) « Pathology and the posture of the la Chapelle-aux-Saints Neandertal », Am. J. Phys Anthropol., 67 : p. 19-41.
- Ogilvie M.D., Curvan B.K. et Trinkaus E. (1989) « Incidence and patterning of dental enamel hypoplasia among the Neandertals », Am. J. Phys. Anthropol., 79 : p. 25-41.
- L'individu aurait vécu jusqu'à l'âge de 50 à 60 ans. Homme de La Chapelle-aux-Saints - Homo neanderthalensis sur Hominidés.com.
- Arensburg, B. et Tillier, A.-M. (1990) « Le langage des néandertaliens », La Recherche, vol. 21, no 224, p. 1084-1086.
- Heim, J.-L, Boë, L.J. et Abry, Ch. (2002) « La parole à la portée du conduit vocal de l'homme de Neandertal. Nouvelles recherches, nouvelles perspectives », Comptes Rendus Palevol, Volume 1, Issue 2, p. 129-134
- Arensburg 1991, Arensburg et al. 1989 ; Rodriguez et al. 2003.[réf. incomplète]
- Martinez 2008[réf. incomplète]
- Boë, L.-J., Heim, J.-L., Honda, K. Maeda, S. (2002) « The potential of Neandertal vowel space was as large as that of modern humans », Journal of Phonetics, 30, 465-484.
- Lieberman, P. (2007) « Current views on Neanderthal speech capabilities: A reply to Boë et al. (2002) », Journal of Phonetics, 35, 552-563.
- Boë, L.J., Heim, J.L., Honda, K., Maeda, S., Badin, P., Abry, C. (2007) « The vocal tract of newborn humans and Neanderthals: Acoustic capabilities and consequences for the debate on the origin of language. A reply to Lieberman », Journal of Phonetics, 35, 564–581.
- Krause, J., Lalueza-Fox, C., Orlando, L., Enard, W., Green, R.E., Burbano, H.A., Hublin, J.-J., Hänni, C., Fortea, J., de la Rasilla, M., Bertranpetit, J., Rosas, A. et Pääbo, S. (2007) - « The derived FOXP2 variant of modern humans was shared with Neandertals », Current Biology, vol. 17, 21, p. 1908-1912.
- Néandertal, un grand bavard de la préhistoire ?
- Kate Wong, « Le crépuscule de l'homme de Néandertal », Pour La Science, n° 386, décembre 2009.
- Baffier D., Les derniers Néandertaliens, le Châtelperronien, La Maison des roches, Paris, 1999
- Pelegrin J., Technologie lithique : Le Châtelperronien de Roc-de-Combe (Lot) et de La Côte (Dordogne), Cahiers du Quaternaire, CNRS, Paris, 1995
- Lewis Binford, Bones, Ancient Men and Modern Myths, Academic Press, 1981
- (en) Hervé Bocherens, « Isotopic biogeochemistry as a marker of Neandertal diet (La biogéochimie isotopique comme marqueur de l'alimentation des Néandertaliens) », dans Anthropologischer Anzeiger, vol. 55, no 2, 1997, p. 101-120
- Cyril Bousquet, « Une spécialiste de Néandertal « pessimiste sur notre avenir » » sur Rue89, 13 février 2011
- Brugal, J.-Ph. et Jaubert, J. (1991) - « Les gisements paléontologiques pléistocènes à indices de fréquentation humaine : un nouveau type de comportement de prédation ? », Paléo, n° 3, pp. 15-41.
- Science & Vie junior, Chasseurs ou charognards ?, Hors série, n°22, octobre 1995, p.68-72
- (en) Peter Ungar, Evolution of the human diet : the known, the unknown, and the unknowable, New York, Oxford University Press, Inc., 2007, relié (ISBN 978-0-19-518346-7) (LCCN 2005036120)
- (en) Amanda G. Henry et coll, « Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium) », dans Proceedings of the National Academy of Sciences, 12 novembre 2010 [texte intégral]
- Agusti, B., Alcalde, G., Güell, A., Juan-Muns, N., Nebot, J, Rueda, J.M. et Terradas, X. (1992) - « Caracoles terrestres en la cueva 120 - Aportacion al conocimiento de la dieta alimenticia durante el Paleolitico medio », Revista de Arqueologia, Ano XII, n° 131, pp. 8-11.
- Cortés-Sánchez, Miguel, Morales-Muñiz, Arturo, Simón-Vallejo, M.D., Lozano-Francisco, M.C., Vera-Peláez, J.L., Finlayson, C., Rodríguez-Vidal, J., Delgado-Huertas, A., Jiménez-Espejo, F.J., Martínez-Ruiz, F., Martínez-Aguirre, M.A., Pascual-Granged, A.J., Bergadà -Zapata, M.M., Gibaja-Bao, J.F., Riquelme-Cantal, J.A., López-Sáez, J.A., Rodrigo-Gámiz, M., Sakai, S., Sugisaki, S., Finlayson, G., Fa, D.A. et Bicho, N.F. (2011) - « Earliest known use of marine resources by Neanderthals », PLoS ONE, 6, 9, pp. e24026.
- Les néandertaliens mangeaient des fruits de mer il y a 150 000 ans, Le Monde, 15 septembre 2011.
- Stringer, C.B., Finlayson, C., Barton, R.N.E., Fernández-Jalvo, Y., Cáceres, I., Sabin, R.C., Rhodes, E.J., Currant, A.P., Rodríguez-Vidal, J., Giles-Pacheco, F. et Riquelme-Cantall, J.A. (2008) - « Neanderthal exploitation of marine mammals in Gibraltar », Proceedings of the National Academy of Sciences, vol. 105, n° 38, pp. 14319-14324.
- Neanderthal diet like early modern human's Preuve de chasse maritime & composition du régime de Néandertal, sur le site du Natural History Museum, vidéo & texte.
- Maureille, B. Les premières sépultures, Le Pommier / Cité des sciences et de l'industrie, (2004)
- Leroi-Gourhan, Arl. (1968) « Le Néanderthalien IV de Shanidar », Bulletin de la Société Préhistorique Française, t. LXV, no 3, p. 79-83.
- Solecki, R.S. (1971) Shanidar, the first flower people, New York, A. Knopf.
- Sommer, J.D. (1999) « The Shanidar IV 'Flower Burial': A Reevaluation of Neanderthal Burial Ritual », Cambridge Archaeological Journal, 9 127-129
- Bar-Yosef et col., La Sépulture Néandertalienne de Kébara (Unité XII) In Otte, M (ed), L'Homme de Néanderthal, vol 5 La Pensée, 1988, p.17-24.
- Maria Dolores Garralda, Bernard Vandermeersch, Les Néandertaliens de la grotte de Combe-Grenal (Domme, France), Paléo, 2000, n°12, p. 213-259
- Bernard Vandermeersch et Bruno Maureille, Les neandertaliens, biologies et cultures, chapitre Les sépultures néandertaliennes, éd. Comité des travaux historiques et scientifiques, 2007, 342 p.
- Bonifay, E. (1965) - « Un ensemble rituel moustérien à la grotte du Regourdou (Montignac, Dordogne) », in: Actes du IVème Congrès de l'UISPP, Rome, vol. II, p. 136-140.
- Bonifay, E. (2002) - « L'Homme de Neandertal et l'ours (Ursus arctos) dans la grotte du Regourdou (Montignac-sur-Vézère, Dordogne, France) », in: L'ours et l'Homme, Tillet, T. et Binford, L., (Éds.), Liège, ERAUL 100, p. 247-254.
- Cavanhié, N. (2007) - Étude archéozoologique et taphonomique des grands carnivores du site paléolithique moyen de Régourdou (Montignac, Dordogne), Université de Toulouse II le Mirail, Mémoire de Master 2.
- « Le Regourdou aujourd’hui… »
- Defleur, A., White, T., Valensi, P., Slimak, L. et Crégut-Bonnoure, E. (1999) - « Neanderthal cannibalism at Moula-Guercy, Ardèche, France », Science, vol. 286, p. 128-131
- Gorjanoviv-Kramberger, D. (1906), Der diluviale Mensh von Krapina in Kroatia, Ein Betrag zur Paläoanthropologie, Kreidel, Wiesbaden.
- Smith, F.H. (1976), The Neandertal remains from Krapina. A descriptive and comparative study, Report of investigation no 15, Department of Anthropology University of Tennessee, Knoxville, 359 p.
- White, T.D., (1992) Prehistoric cannibalism at Mancos 5MTUMR-2346, Princeton University Press, 462 p.
- White, T.D., Toth, N. (1991) « The Question of Ritual Cannibalism at Grotta Guattari », Current Anthropology, Vol. 32, No. 2 (Apr., 1991), p. 118-138
- cf. également les références de l'article Grotte Guattari
- Soressi, M. et d’Errico, F. (2007) - « Pigments, gravures, parures : les comportements symboliques controversés des Néandertaliens » in Bernard Vandermeersch et Bruno Maureille (dir.), Les Néandertaliens, biologie et culture, CTHS, p. 297-309.
- Peresani, M., Fiore, I., Gala, M., Romandini, M. et Tagliacozzo, A. (2011) - « Late Neandertals and the intentional removal of feathers as evidenced from bird bone taphonomy at Fumane Cave 44 ky B.P., Italy », Proceedings of the National Academy of Sciences, 108, 10, pp. 3888-3893.
- Lorblanchet, M. (1999) La naissance de l'Art. Genèse de l'art préhistorique, Paris, Éd. Errance, 304 p.
- Révillion, S. et Tuffreau, A. (1994) Les industries laminaires du Paléolithique moyen, Éditions du CNRS, Dossiers de Documentation Archéologique no 18, 193 p.
- Pettitt. P.B. (1999) « Disappearing from the World: An Archeological Perspective on Neanderthal Extinction », Oxford Journal of Archeology, 18, p. 217-239.
- Guk, Y. (1997) « Why's the Neanderthal Gone ? », Science in Russia, 4, p. 50-54.
- Gelly, R. (1992) « L'énigme de Néandertal », Sciences et Avenir, no d'avril, p. 64-69.
- Flores, J. C. (1998) « A Mathematical Model for Neanderthal Extinction », Journal of Theoretical Biology, Volume 191, Issue 3, p. 295-298.
- Horan, R.D., Bulte, E. et Shogren, J.F. (2005) « How trade saved humanity from biological exclusion: an economic theory of Neanderthal extinction », Journal of Economic Behavior & Organization, Volume 58, Issue 1, p. 1-29.
- La Genèse de l'Humanité, P.U.F., 6e édition, 4e trimestre 1961, Paris
- d’Errico, F. et Sánchez Goñi, M.F. (2003), « Neanderthal extinction and the millennial scale climatic variability of OIS 3 », Quaternary Science Reviews, 22, p. 769–788.
- hypothèse présentée dans le roman Dance of the Tiger. A novel of the Ice Age (1978) et explicitée dans une note de l'auteur à la fin de l'ouvrage.
- L'homme de Néandertal n'est pas mort de froid sur Agence Science-Presse, 17 septembre 2007.
- « 10 scénarios pour une disparition » in Neandertal - Enquête sur une disparition, Les Dossiers de la Recherche, n° 24, août-octobre 2006.
- Finlayson, C., Fa, D.A., Finlayson, G., Pacheco, F.G. et Vidal, J.R., (2004), « Did the moderns kill off the Neanderthals? A reply to F. d’Errico and Sánchez Goñi », Quaternary Science Reviews, Volume 23, Issues 9-10, p. 1205-1209.
- Serre, D., et al., (2004) - « No evidence of Neandertal mtDNA contribution to early modern humans », PLoS Biology, vol. II, n° 3, mars 2004.
- Eswaran, V. et al., (2005) - « Genomics refutes an exclusively African origin of humans », Journal of Human Evolution, vol XLIX, juillet 2005.
- Nouveau ramdam chez nos vieux ancêtres
- Plagnol, V., Wall, J.D., (2006) - « Possible Ancestral Structure in Human Populations », PLoS Genetics, vol. 2, n° 7, juillet 2006.
- Evans, P.D., Mekel-Bobrov, N., Vallender, E.J., Hudson R.R. et Lahn, B.T., (2006) - « Evidence that the adaptive allele of the brain size gene microcephalin introgressed into Homo sapiens from an archaic Homo lineage », Proceedings of the National Academy of Sciences, vol. 103, n°. 48, pp. 18178-18183.
- Hervé Morin, « Il y a du Néandertal en nous », Le Monde, 8 mai 2010
- On a tous quelque chose de Neandertal, Rachel Mulot, Sciences et Avenir, juin 2010, page 30.
Voir aussi
Bibliographie
- Juan Luis Arsuaga, Le Collier de Néandertal, Odile Jacob, 2004 (ISBN 978-2-7381-1422-8)
- Claudine Cohen, Un Néandertalien dans le métro, coll. Science ouverte, Seuil, 2007 (ISBN 978-2-02-092557-0)
- collectif, Neandertal, enquête sur une disparition, Les Dossiers de La Recherche n°24, août-octobre 2006
- collectif, Neandertal, Tallandier, 2006, (ISBN 978-2-84734-366-3)
- Jean-Jacques Hublin, « Origine et évolution des Néandertaliens », in Y. Coppens et P. Picq (dir.), Aux origines de l'humanité, Fayard, 2001 (ISBN 978-2-213-60369-8), vol. 1
- Jacques Jaubert, Chasseurs et artisans du Moustérien, La Maison des Roches, 1999 (ISBN 978-2-912691-05-7)
- Bruno Maureille, Les premières sépultures, Le Pommier / Cité des sciences et de l'industrie, 2004 (ISBN 978-2-7465-0203-1)
- Bruno Maureille, Qu'est-il arrivé à l'homme de Néandertal ?, coll. Les petites pommes du savoir, le Pommier, 2008, (ISBN 978-2-7465-0389-2)
- Étienne Patte, Les Néanderthaliens - Anatomie, physiologie, comparaisons, Paris, Masson, 1955.
- Eric Trinkaus et Pat Shipman, Les Hommes de Neandertal, Seuil, 1996 (ISBN 978-2-02-013178-0)
- Bernard Vandermeersch et Bruno Maureille, Les Néandertaliens, biologie et culture, CTHS, 2007, (ISBN 978-2-7355-0638-5)
Filmographie
- Où est passé Néandertal ?, interview de Jean-Luc Voisin, docteur en paléontologie humaine.
- Neanderthal, docu-fiction télévisé britannique (durée 100 min) réalisée par Tony Mitchell en 2000 (diffusion en 2001)
- Reconstitution du crâne néandertalien de la Chapelle aux Saints par J.-L. Heim, film realmedia de 18 minutes.
- Il était une fois... l'Homme : l'homme du Néanderthal, dessin animé de Albert Barillé et Fabrice Matéo.
- Interview de Jean-Louis Heim et Marylène Patou-Matis au sujet de l'Homme de Néandertal
Articles connexes
- Paléolithique moyen
- Moustérien
- Châtelperronien
- Néandertal en Poitou-Charentes
- Neanderthal genome project
Liens externes
- (de) (en) (nl) Le musée de Neandertal
- Néandertaliens et Hommes de la Solo : deux exceptions naturelles ?, article de Bruno Maureille et Bernard Vandermeersch sur le site du CNRS.
- Le Musée de l'Homme de Néandertal de la Chapelle-aux-Saints
- Les dents de l'homme de Néandertal plaident pour son appartenance à une espèce distincte
- (en) La vie des Néandertaliens n'était pas plus dure que celle des Inuit modernes, travaux de Debbie Guatelli-Steinberg (images de dents présentant des hypoplasies)
- Néandertal et Cro-Magnon n'auraient jamais mélangé leurs gènes
- (en) Neanderthal or Neandertal ?

La version du 14 juin 2006 de cet article a été reconnue comme « article de qualité », c'est-à-dire qu'elle répond à des critères de qualité concernant le style, la clarté, la pertinence, la citation des sources et l'illustration.  Portail de la Préhistoire
Portail de la Préhistoire Portail de la paléontologie
Portail de la paléontologie Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant Portail de l’anthropologie
Portail de l’anthropologie Portail des mammifères
Portail des mammifères
Catégories :- Hominidé disparu
- Préhistoire
- Les caractères plésiomorphes des Néandertaliens sont :

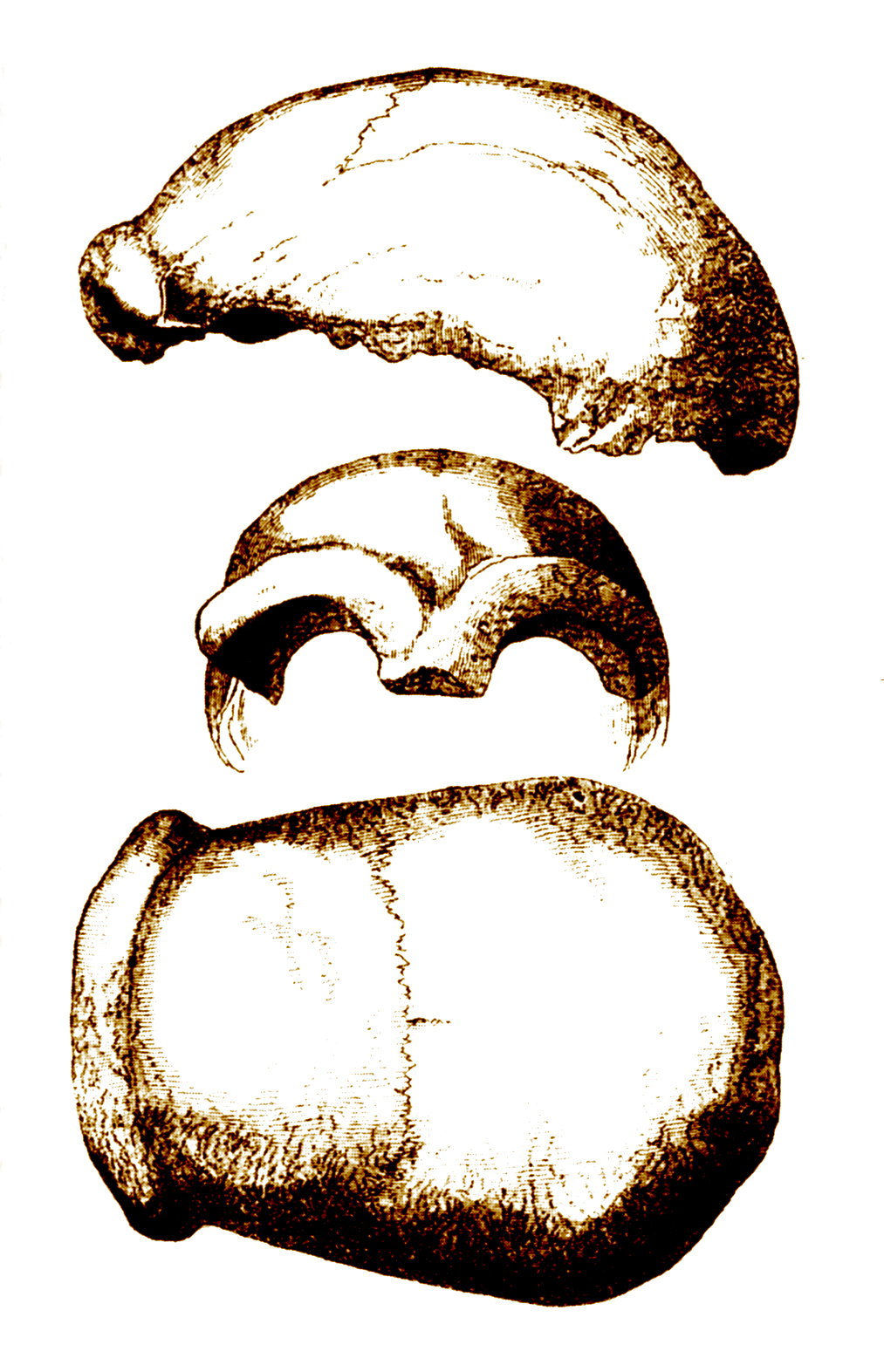




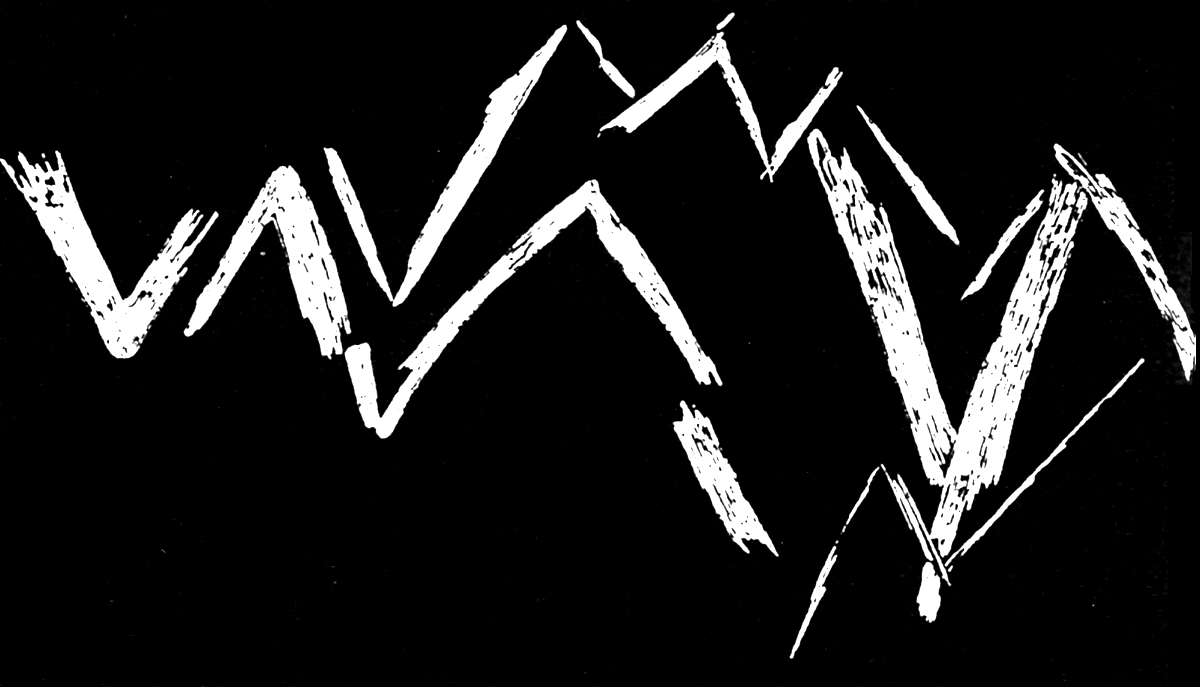

Wikimedia Foundation. 2010.