- Acide glutamique
-
Acide glutamique 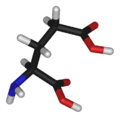
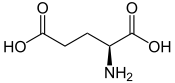
Acide L ou S(+)-glutamique (énantiomère biologique)
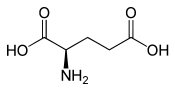
Acide D ou R(-)-glutamiqueGénéral Nom IUPAC Acide (S)-2-aminopentanedioïque Synonymes E, Glu No CAS (racémique)
(D) ou R(–)
(L) ou S(+)No EINECS Code ATC AB01 No E E620 FEMA SMILES InChI Propriétés chimiques Formule brute C5H9NO4 [Isomères] Masse molaire[1] 147,1293 ± 0,006 g·mol-1
C 40,82 %, H 6,17 %, N 9,52 %, O 43,5 %,pKa 2,16
4,15
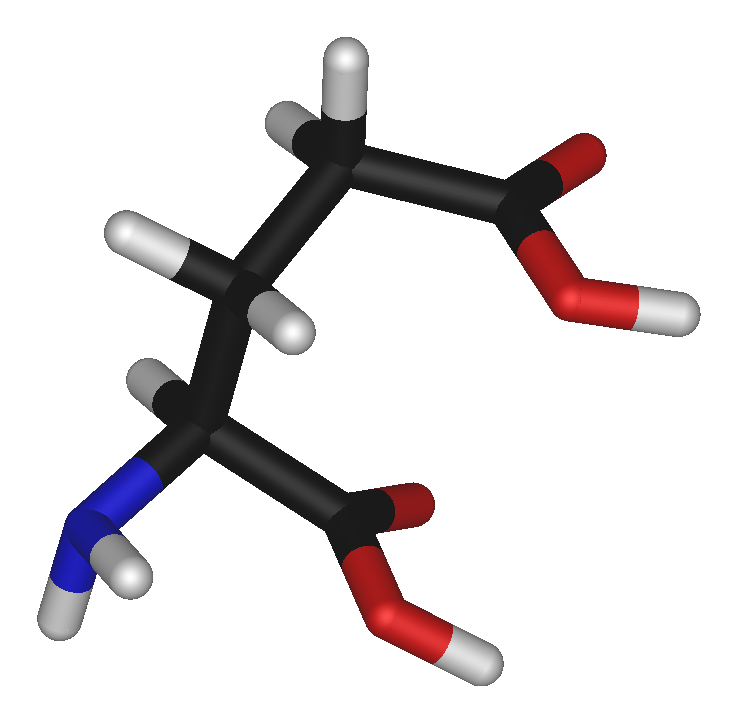
9,58Propriétés physiques T° fusion 247 °C à 249 °C Solubilité Peu soluble dans l'éthanol Masse volumique 1,538 Propriétés biochimiques Codons GAA, GAG pH isoélectrique 3,22 Acide aminé essentiel Non Occurrence dans les protéines humaines 6,3 % Précautions SIMDUT[2] Produit non contrôlé Unités du SI & CNTP, sauf indication contraire. L'acide glutamique ou glutamate (la forme anionique de l'acide glutamique) est l'un des 20 acides-α-aminés naturels constituant les protéines. Sa chaîne latérale contient un résidu carboxyle, ce qui en fait un acide aminé « acide », dicarboxylique, polaire.
On l'abrège Glu, ou E s'il est incorporé à une chaîne peptidique. Glx (Z) est une désignation commune à l'acide glutamique et à la glutamine (Gln, Q).
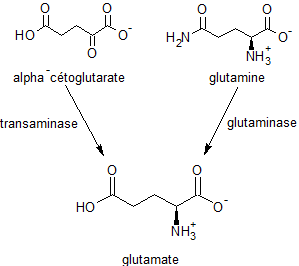
L'acide glutamique joue un rôle critique pour sa propre fonction cellulaire, mais n'est pas considéré comme un nutriment essentiel chez les humains car le corps peut le fabriquer à partir de composés plus simples, comme par transamination de l'α-cétoglutarate, produit intermédiaire de l'oxydation des sucres simples dans la mitochondrie (cycle de Krebs).
Le glutamate, forme ionisée de l'acide glutamique, est le neurotransmetteur excitateur le plus important du système nerveux central. Son action est contrebalancée par les effets inhibiteurs du GABA dont il est d'ailleurs, le précurseur principal. Lorsque l'équilibre est rompu et que la concentration en glutamate atteint un niveau excessif dans la fente synaptique, il peut hyperstimuler les neurones et provoquer leur mort.
Sommaire
Neurotransmetteur
L'acide glutamique n'est pas seulement l'une des briques élémentaires utilisées pour la synthèse des protéines, c'est aussi le neurotransmetteur excitateur le plus répandu dans le système nerveux central (cerveau + moelle épinière) et un précurseur du GABA dans les neurones GABAergiques. Il serait le médiateur de près de 50% des neurones centraux[3]. C'est le neurotransmetteur excitateur principal des neurones pyramidaux. Ce sont des neurones de projection trouvés dans les connexions corticostriatales et corticothalamiques.
Le glutamate active les récepteurs ionotropes AMPA, NMDA et kaïnate KAR, ainsi que les récepteurs métabotropes. L'acide glutamique lui même ne franchit que mal la barrière hémato-encéphalique, à moins d'avoir été transformé en L-glutamine utilisable par le cerveau comme carburant cellulaire et pour la synthèse protéique.
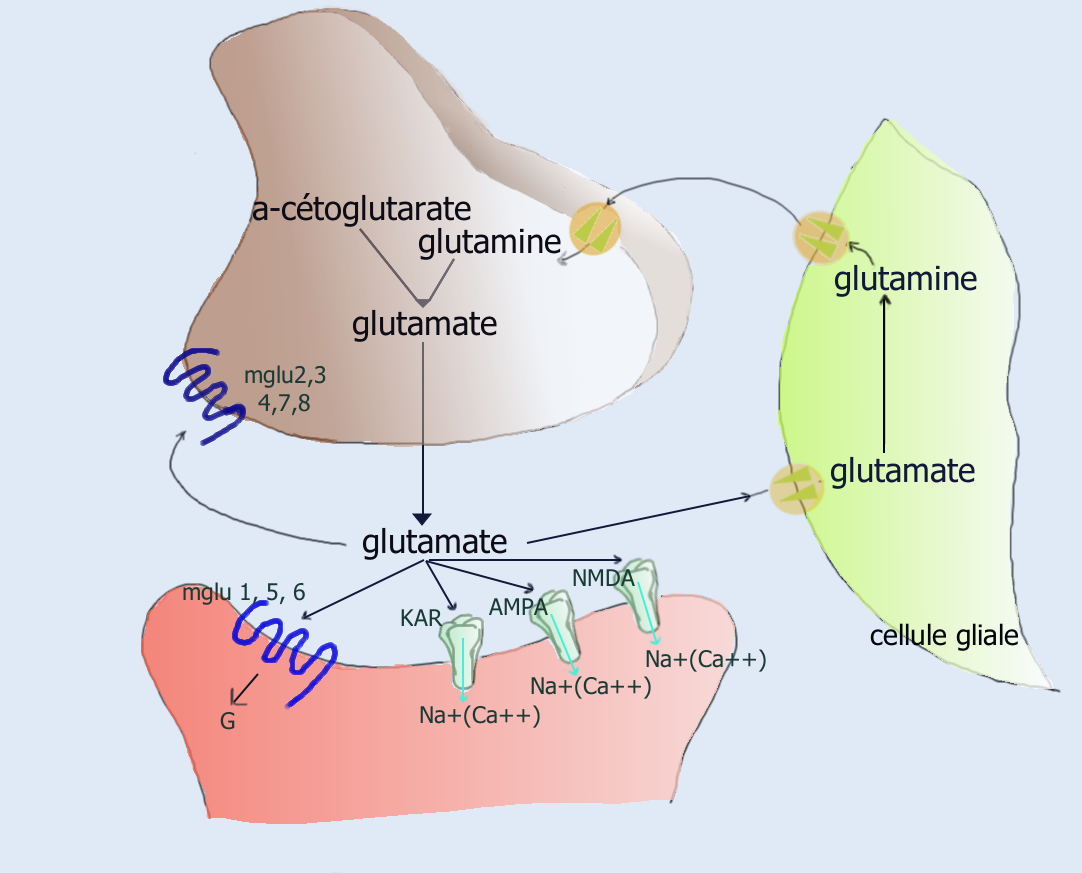
Le glutamate est formé dans les mitochondries des neurones suivant deux voies : soit par une transamination de l'alpha-cétoglutarate (un métabolite du cycle de Krebs mitochondrial), soit par une désamination oxydative de la glutamine par l'enzyme glutaminase. Il est ensuite stocké dans les vésicules des neurones glutaminergiques.
Lorsqu'un potentiel d'action arrive à l'extrémité de l'axone, les vésicules déversent par exocytose, leur glutamate dans la fente synaptique.
Le glutamate se lie alors à divers récepteurs post-synaptiques. Les récepteurs-canaux, NMDA, AMPA et KAR (ou récepteurs ionotropes) laissent entrer des cations (Na+, K+, Ca2+). L'entrée de charges positives, active les canaux sodiques voltage-dépendants qui déclenchent la décharge d'un potentiel d'action.
Les récepteurs couplés aux protéines G, mglu1 à mglu8 (ou récepteurs métabotropes) activent indirectement des canaux ioniques par l'intermédiaire d'une cascade de réactions impliquant les protéines G. L'intervention de ces nombreux intermédiaires ralentit le traitement de l'information mais permet des activations multiples.
Dans la fente synaptique le glutamate n'est pas métabolisé. Les molécules qui ne sont pas captées par les récepteurs post-synaptiques sont alors principalement captées par les cellules gliales voisines et métabolisées en glutamine :
Glutamate + ATP + NH3 → Glutamine + ADP + phosphate
 Synthèse du glutamate
Synthèse du glutamate
 Synapse glutamatergique
Synapse glutamatergiqueLa glutamine est ensuite exportée dans le milieu extracellulaire où elle sera captée par les neurones glutaminergiques. Ce recyclage bicellulaire est nommé cycle glutamate-glutamine.
Les récepteurs ionotropes du glutamate, NMDA, AMPA, KAR
Ces récepteurs-canaux sont constitués de l'association de sous-unités différentes, sous forme généralement de tétramères. Ces sous-unités sont des protéines transmembranaires codées par des gènes donnés dans le tableau ci-dessous. Cette diversité serait encore augmentée par l'épissage alternatif de plusieurs sous-unités[4],[5].
Famille de récepteurs Sous-unité Gène Chromosome
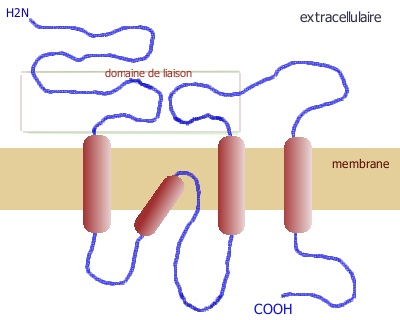
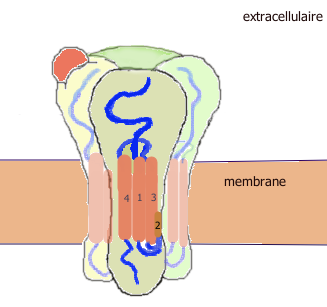
(humain)NMDA NR1 GRIN1 9q34.3 NR2A GRIN2A 16p13.2 NR2B GRIN2B 12p12 NR2C GRIN2C 17q24-q25 NR2D GRIN2D 19q13.1qter NR3A GRIN3A 9q31.1 NR3B GRIN3B 19p13.3 AMPA GluR1 GRIA1 5q33 GluR2 GRIA2 4q32-33 GluR3 GRIA3 Xq25-26 GluR4 GRIA4 11q22-23 KAR GluR5 GRIK1 21q21.1-22.1 GluR6 GRIK2 6q16.3-q21 GluR7 GRIK3 1p34-p33 KA-1 GRIK4 11q22.3 KA-2 GRIK5 19q13.2 Les sous-unités des récepteurs ionotropes du glutamate comportent toutes une extrémité C-terminale intracellulaire et 3 hélices transmembranaires.
 Structure d'une sous-unité des récepteurs ionotropes du glutamate
Structure d'une sous-unité des récepteurs ionotropes du glutamate Récepteur tétramérique, formé de 4 sous-unités
Récepteur tétramérique, formé de 4 sous-unitésLes récepteurs NMDA sont des tétramères, formés de deux sous-unités NR1 et de deux sous-unités NR2 (NR2A, NR2B, NR2C ou NR2D). Les sous-unités NR1 sont obligatoires, tandis que les sous-unités NR2 spécifient les propriétés électrophysiologiques des récepteurs NMDA, tel leur sensibilité au glutamate, leur perméabilité au calcium, l'inhibition du magnésium. Les sous-unités NR3 A et B ont un effet inhibiteur sur l'activité des récepteurs.
Le L-glutamate se fixe à l'extrémité N-terminale des sous-unités NR2A et NR2B du récepteur NMDA. Pour l'activer, la glycine doit aussi se fixer sur NR1. C'est un co-agoniste du glutamate. La fixation du glutamate sur le domaine extracellulaire entraîne un changement de conformation des sous-unités et l'ouverture du pore de façon progressive.
Plusieurs molécules sont aussi susceptibles de moduler l'activité des récepteurs NMDA. C'est le cas des ions magnésium Mg2+ qui se fixent sur des sites situés dans le canal ionique et les obstruent. La dépolarisation membranaire, induite par l'activation des récepteurs AMPA permet de déloger ces ions et de lever le blocage.
Les récepteurs NMDA sont largement répandus dans tout le système nerveux, généralement colocalisés avec les récepteurs AMPA. Ils sont très nombreux dans le cerveau antérieur avec une densité élevée dans la région CA1 de l'hippocampe[3].
La stimulation des récepteurs post-synaptiques génère des potentiels post-synaptiques excitateurs. Elle s'accompagne d'entrée de calcium entraînant la stimulation de la protéine kinase activée par le couple calcium-calmoduline, CAM-kinase 2, impliquée dans le phénomène de potentialisation à long terme (LTP long term potentialisation). Ce phénomène de LTP est à la base des processus d'apprentissage et de mémorisation.
L'entrée excessive de calcium par hyperstimulation des récepteurs NMDA peut induire une mort neuronale par apoptose qui serait impliquée dans la neurodégénérescence observée dans la maladie de Parkinson, la maladie d'Alzheimer et les accidents ischémiques cérébraux.
L'activation des récepteur AMPA déclenche l'ouverture d'un canal perméable au potassium et au sodium. Ainsi, le potassium sortira de la cellule alors que le sodium y entrera. Dans certain cas, il y a en plus une entrée de calcium, cela dépendant de la structure de l'AMPA (plus particulièrement de la présence ou non d'une sous-unité GluR2 éditée). Ainsi, il permettra la dépolarisation de la membrane du neurone cible du glutamate.
Les récepteurs kaïnate KAR sont en général pré-synaptiques et situés sur des synapses glutamatergiques ou gabaergiques. On en trouve principalement dans l'hippocampe et dans les couches infragranuleuses du cortex cérébral. Les récepteurs KAR pré-synaptiques sont inhibiteurs par activation des protéines G trimériques Gi et Gq qui diminuent l'exocytose du glutamate ou du GABA. Il existe aussi des récepteurs KAR postsynaptiques excitateurs.
Les récepteurs métabotropes, mglu
Les récepteurs métabotropes du glutamate se présentent sous la forme d'homodimères reliés par des ponts disulfures[3].
Ces récepteurs mglu sont localisés sur les neurones du système nerveux central et sur les cellules gliales.
Les récepteurs mglu1 et mglu5 post-synaptiques participent au phénomène de potentialisation à long terme. Les mglu1 sont abondants dans l'hippocampe et le thalamus. Les mglu5 se trouvent dans le cortex, le striatum ventral et des parties de l'hippocampe, de l'amygdale et du thalamus. Ces deux récepteurs sont couplés aux proteines Gq et aux phospholipases C, ces dernières catalysant la formation du diacylglycerol (DAG) et de l'inositol trisphosphate (IP3). Le DAG est un activateur de la proteine kinase C. L'IP3, lui, se lie a son récepteur situé sur le reticulum endoplasmique et permet la libération de calcium du reticulum vers le cytoplasme.
Le récepteur mglu6 lui aussi présent au niveau postsynaptique, produit une dépolarisation lente.
Les récepteurs mglu2, 3, 4, 7, et 8 sont présynaptiques et provoquent une diminution de libération du glutamate.
Rôles
Le glutamate est soupçonné de jouer un rôle dans les fonctions cérébrales d'apprentissage et de mémorisation, mais en trop grande concentration il est susceptible d'entraîner des effets pathologiques comme ceux rencontrés dans la sclérose latérale amyotrophique, la maladie d'Alzheimer, le lathyrisme.
Le psychotrope phencyclidine (plus connu sous le nom de PCP ou poudre d'ange) est un inhibiteur non compétitif du glutamate pour le récepteur NMDA par stabilisation des canaux ioniques de ces récepteurs dans l'état ouvert. Il provoque un comportement semblable à la schizophrénie. Pour la même raison, des doses de kétamine inférieures à celles utilisées en anesthésiologie entraînent des effets dissociatifs et des hallucinations.
L'action des glutamates se révèle très difficile à étudier de par sa nature éphémère. Une équipe de l'université Stanford a développé un nanodétecteur pour détecter la sortie neuronale du glutamate.
Ce détecteur, une protéine, possède une paire de lobes articulés autour d'une charnière, comme le piège de la plante carnivore Dionée attrape-mouche. Quand le glutamate se lie aux protéines, les lobes se referment. Deux protéines fluorescentes, extraites d'une méduse, sont fixées au détecteur. L'une d'entre elles émet une luminescence bleue tout en excitant la seconde qui émet à son tour une luminescence jaune. Au moment où les lobes se referment sur le glutamate, la protéine bleue s'éloigne de la jaune diminuant l'intensité de la luminescence. Cette diminution signale que le glutamate est sorti du neurone. Ce détecteur ne peut, pour le moment, que se situer sur la surface de la cellule : il ne peut donc indiquer la présence de glutamate qu'à l'extérieur de la cellule[6].
On peut provoquer par ultraviolet la libération d'une forme spécifique de glutamate (glutamate en cage, "caged glutamate", après la stimulation UV, le glutamate est actif, "uncaged glutamate") à un endroit choisi d'un ou de plusieurs neurones. Cette méthode de photostimulation s'est révélée très efficace pour cartographier les connexions inter-neuronales.
Le glutamate monosodique est utilisé dans le cadre d'expérience pour induire l'obésité de rats avec des objectifs expérimentaux tels que la détermination des mécanismes de l'obésité[7].
Neurotoxicité
En concentrations excessives, le glutamate déclenche un processus dit d'excitotoxicité, délétère, voire mortel, pour les neurones, particulièrement en cas d'activation des récepteurs NMDA.
La toxicité peut être due à :
- un influx excessif et incontrôlé de Ca2+ dans la cellule, dépassant sa capacité de stockage. Il s'ensuit des altérations mitochondriales[8], conduisant à une libération de cytochrome p450, menant à l'apoptose.
- une surexpression de facteurs de transcription de gènes pro-apoptotiques, ou une répression des facteurs anti-apoptotiques, médié par le glutamate et le calcium.
Ces théories se fondent sur l'observation post-mortem de neurodégénération chez des patients épileptiques connus.
Accès d'épilepsie
Le glutamate a été impliqué dans les crises d'épilepsie, au vu de la dépolarisation foudroyante (une seconde) qu'il provoque in vitro par microinjection intraneuronale, laquelle dépolarisation reproduit le phénomène de shift de dépolarisation paroxystique observé sur EEG lors d'une crise in vivo[9].
Un mécanisme suggéré est la baisse du potentiel membranaire de repos au niveau du foyer épileptique, qui provoquerait l'ouverture de canaux voltage-dépendants, provoquant un influx de glutamate qui entretiendrait la dépolarisation.
Glutamate libre naturellement présents dans les aliments
L'acide glutamique est un acide aminé très commun, présent dans de nombreuses protéines végétales et animales. C'est même l'acide aminé le plus abondant de l'alimentation humaine. Sa saveur spécifique n'est détectable que s'il est présent à l'état libre. Cette saveur, différente du sucré, du salé, de l'acide et de l'amer, a été reconnue pour la première fois en 1908, par le scientifique japonais Kikunae Ikeda, qui la nomma umami (savoureux).
En tant que composant des protéines, le glutamate reste sans saveur, mais il peut exprimer toute sa sapidité en étant libéré par hydrolyse des protéines lors des processus de fermentation, vieillissement, mûrissement ou cuisson des produits alimentaires[10].
L'acide glutamique libre est naturellement abondant dans des fromages comme le parmesan, la sauce soja ou les tomates. C'est lui qui joue un rôle fondamental dans la saveur des fromages, des crustacés et des bouillons de viande. Voici les principales mesures faites par Kumiko Ninomiya[11] (1998)
Glutamate libre dans les aliments (d'après Ninomiya 1998) Classe Produit alimentaire Glutamate libre (mg/100g) FROMAGES Parmesan 1680 Emmental 308 ALGUES Kelp 1608 Wakame (Undaria) 9 SAUCE SOJA chinoise 926 japonaise 782 coréenne 1264 VEGETAUX Tomate 246 Petits pois 106 Chou 50 VIANDES Bœuf 10 Poulet 22 CRUSTACES Coquille Saint-Jacques 140 Crabe royal 72 Dans certains aliments, le glutamate peut être très abondant sous sa forme liée et être libéré en partie lors de la préparation culinaire. Voici quelques mesures plus anciennes de Giacometti[12] (1979) :
Glutamate lié ou libre dans les aliments,
en mg/100g (d'après Giacometti 1979 )Classe Produit alimentaire Glutamate lié Glutamate libre POISSON Morue 2101 9 Saumon 2216 36 VEGETAUX Petits pois 5583 200 Tomates 238 140 PRODUIT LAITIER Lait de vache 819 2 Parmesan 9847 1200 VIANDE Poulet 3309 44 Bœuf 2846 33 Le glutamate, additif alimentaire
Sous le code de E620, le glutamate est utilisé comme exhausteur de goût des aliments. Il est additionné aux préparations alimentaires pour renforcer leur goût.
Outre le glutamate[13], le Codex Alimentarius a reconnu aussi comme exhausteurs de goût ses sels de sodium (E621), de potassium (E622), calcium (E623), ammonium (E624) et magnésium (E625) :
Glutamates (daprès Jinap et al 2010) N° Descriptif E620 Acide glutamique E621 Glutamate monosodique, GMS E622 Glutamate monopotassique E623 Diglutamate de calcium E624 Glutamate d'ammonium E625 Diglutamate de magnésium Le glutamate monosodique (GMS), sel sodique de l'acide glutamique, apporte une saveur semblable à celle de cet acide aminé avec l'avantage de contenir 3 fois moins de sodium que le sel de table.
Le glutamate (ou ses sels) est souvent présent dans les plats tout prêt (soupes, sauces, chips, plats cuisinés). Il est aussi couramment utilisé en cuisine asiatique.
Il est actuellement fréquemment utilisé en combinaison avec des arômes dans les apéritifs (goût bacon, goût fromage). Cela permet de rehausser le goût de bacon, du fromage, etc. Il est rare de trouver un apéritif qui n'en contienne pas. On en trouve aussi dans certaines capsules de médicaments mais pas pour ses fonctions gustatives. Enfin, il est le composant majoritaire des auxiliaires de cuisine (bouillons cubes, fonds de sauces, sauces, etc). Il est à noter que les produits portant la mention Bio comportent rarement du glutamate monosodique.
Son utilisation est très controversée mais défendue par le lobby de l'agro-alimentaire puisque permettant de réduire de façon substantielle les coûts de production, en remplaçant les arômes naturels plus onéreux.
Amélioration du goût et régime diététique
Des études ont montré que l'addition de glutamate de sodium (GMS) augmentait la palatabilité[14] des aliments et stimulait l'appétit (Belliste[15] 1998, Yamaguchi et al[16] 2000). Cet effet est plus net chez les enfants et les adultes que chez les personnes âgées. En faisant goûter des aliments avec différentes concentrations de GMS, on observe que certains ne sont pas améliorés (comme les produits sucrés) et que les autres ont une concentration optimum préférée[17]. Et bien sûr, cette concentration optimum varie d'un individu à l'autre. D'après les études de F. Bellisle[17], la concentration préférée des Européens (qui se trouve dans la fourchette 0,6 à 1,2%), tend à être supérieure à celle des Asiatiques. En général, l'assaisonnement au glutamate convient bien aux produits salés et aigrelets. La dose à ajouter, recommandée par Jinap et Hajeb[13], est de 0,5 à 4 g pour 500 g de nourriture, soit très peu, car un excès gâche rapidement le goût.
L'addition de GMS à un aliment, en améliorant son goût, ne risque-t-elle pas d'induire une consommation excessive, pouvant éventuellement conduire à une prise de poids? Cette hypothèse a été testée chez un groupe de personnes âgées (de 84 ans en moyenne) vivant dans des maisons spécialisées durant un an[15]. L'addition de GMS à des aliments intéressants sur le plan nutritionnel (soupe, légumes, féculents) a induit une augmentation de leur consommation. Mais la taille totale de la prise alimentaire n'a pas variée, puisque l'accroissement de la consommation de produits assaisonnés au glutamate a été compensée par la diminution de la prise d'aliments servis plus tard, comme les desserts sucrés. La même observation a été faite auprès de patients diabétiques hospitalisés. A nouveau, les patients consommaient spontanément plus de produits recommandés assaisonnés de glutamate et moins d'autres produits, ce qui ne changeait pas l'apport énergétique global des repas. L'addition de GMS permet donc de réorienter les choix alimentaires.
Le syndrome du restaurant chinois
Quelques cas d'intolérance au glutamate monosodique ont été rapportés, mais aucune recherche n'a permis d'établir un rapport entre glutamate monosodique et des intolérances d'une façon concluante.
De manière générale, le glutamate monosodique a longtemps été décrit comme responsable du « syndrome du restaurant chinois » du fait que la cuisine asiatique est très riche en glutamate[13]. En cas de réaction, le premier symptôme est un flush (coloration rouge intense et passagère du visage, du cou et du torse). Peuvent apparaitre aussi : yeux injectés de sang, céphalées, sensation de brûlures survenant au niveau du tronc et irradiant vers la périphérie, impression d'oppression thoracique, brûlures de la tête, bouffées de chaleur, nausées et vomissements, sensation d'étouffement, difficultés à respirer, tendance à faire des malaises…
Cette affection survient un quart d'heure à une demi-heure après ingestion et peut durer jusqu'à deux heures. La puissance de cette réaction n'est pas forcément proportionnelle à la dose ingérée, quelques milligrammes suffisent. Son lien direct avec l'ingestion de glutamate n'est pas scientifiquement prouvée. À peu près une personne sur 5 000 serait sensible à son ingestion, mais curieusement une sur 50 000 dans les pays asiatiques.
Une étude randomisée, en double aveugle et contre placebo, menée en commun par quatre grandes universités américaines[18], publiée en 2000, reste à ce jour l'étude la plus complète et la mieux conduite, sur les effets potentiels de la consommation de glutamate (Geha et als[19], 2000). Les 130 sujets recrutés qui disaient avoir connu le syndrome du restaurant chinois ont été soumis à une série de tests sur une boisson contenant 5 g de GMS versus placebo. S'ils répondaient positivement au premier test, par au moins deux symptômes, ils étaient examinés suivant un second protocole, cherchant un effet de dépendance de dose et une reproduction des réactions. Or seulement la moitié de ceux qui avaient eu une réaction à 5 g de GMS et pas au placebo réagirent de manière similaire au protocole suivant, suggérant une inconsistence dans les réponses. Dans les protocoles suivants, les sujets qui avaient répondu positivement à 5 g de GMS et pas au placebo, devaient avaler des capsules (pour qu'ils n'aient aucun indice de goût), à jeun ou au cours d'un petit déjeuner. Seuls, deux sujets manifestèrent des réponses consistantes au glutamate. Au final, les résultats suggèrent que de fortes doses de GMS prises sans nourriture peuvent induire plus de symptômes que des placebos chez des individus qui pensent être sensibles au glutamate. Mais aucun effet persistant n'a été observé au cours des tests successifs.
De nombreux aliments utilisés dans la cuisine occidentale sont riches en glutamate monosodique comme l'asperge verte (106mg/100g), la coquille Saint-Jacques (140mg/100g) et la tomate (246 mg/100g), et malgré leur fort taux en glutamate, ils ne sont pas connus pour être des déclencheurs de migraines, mais des intolérances ou allergies à ces produits sont régulièrement rapportées.
Notes et références
- Masse molaire calculée d’après Atomic weights of the elements 2007 sur www.chem.qmul.ac.uk.
- « Acide glutamique (l-) » dans la base de données de produits chimiques Reptox de la CSST (organisme québécois responsable de la sécurité et de la santé au travail), consulté le 23 avril 2009
- Yves Landry, Jean-Pierre Gies, Pharmacologie. Des cibles vers l'indication thérapeutique, Dunod, 2009 (2{exp
- Dingledine R, Borges K, Bowie D, Traynelis SF, « The glutamate receptor ion channels », dans Pharmacol. Rev., vol. 51, no 1, mars 1999, p. 7–61 [lien PMID]
- Andersson O, Stenqvist A, Attersand A, von Euler G., « Nucleotide sequence, genomic organization, and chromosomal localization of genes encoding the human NMDA receptor subunits NR3A and NR3B », dans Genomics, vol. 78, no 3, décembre 2001, p. 178–84 [lien PMID, lien DOI]
- (en) Okumoto, S., et al., « Detection of glutamate release from neurons by genetically encoded surface-displayed FRET nanosensors », dans Proceedings of the National Academy of Sciences In Press: (2005), .
- (en) Rosana CT Mori, Regina B. Guimarães, Cláudia MO Nascimento et Eliane B. Ribeiro, « Lateral hypothalamic serotonergic responsiveness to food intake in rat obesity as measured by microdialysis », dans Can. J. Physiol. Pharmacol., vol. 77, no 4, 1999, p. 286–292 [lien DOI (page consultée le 24 octobre 2008)].
- Manev H, Favaron M, Guidotti A, Costa E, « Delayed increase of Ca2+ influx elicited by glutamate: role in neuronal death », dans Mol. Pharmacol., vol. 36, no 1, juillet 1989, p. 106–12 [texte intégral, lien PMID]
- B.S. Meldrum *, M.T. Akbar, A.G. Chapman, « Glutamate receptors and transporters in genetic and acquired models of epilepsy », dans Epilepsy Research, vol. 36, 1999, p. 189-204
- S. Jinap, P. Hajeb, « Glutamate. Its application in food and contribution to health », dans Appetite, vol. 55, 2010, p. 1-10
- Kumiko Ninomiya, « Natural occurrence », dans Food Reviews International, vol. 14, no 2-3, 1998 [texte intégral]
- Filer LJ, Garattini MR, Kare MR, Reynolds WA, Wurtman RJ,, Glutamic acid, advances in biochemistry and physiology, Raven Press, 1979
- S. Jinap, P. Hajeb, « Glutamate. Its application in food and contribution to health », dans Appetite, vol. 55, 2010, p. 1-10
- définie comme l'évaluation hédonique des caractéristiques orosensorielles des aliments dans des conditions standardisées (Sorensen et al, 2003)
- France Bellisle, « Effects of monosodium glutamate on human food palatability. », dans Ann N Y Acad Sci., vol. 855, 1998, p. 438-41
- Yamaguchi S, Ninomiya K., « Umami and food palatability. », dans J. Nutri., vol. 130, no 4S suppl, 2000
- France Bellisle, « Experimental studies of food choices and palatability responses in European subjects exposed to the Umami taste », dans Asia Pac J Clin Nutr., vol. 17, no suppl 1, 2008
- Boston University, Harvard University, Northwestern University et UCLA
- Raif S. Geha, MD,a Alexa Beiser, PhD,b Clement Ren, MD,a Roy Patterson, MD,c Paul A. Greenberger, MD,c Leslie C. Grammer, MD,c Anne M. Ditto, MD,c Kathleen E. Harris, BS,c Martha A. Shaughnessy, BS,c Paul R.Yarnold, PhD,c Jonathan Corren, MD,d and Andrew Saxon, MDd, « Multicenter, double-blind, placebocontrolled, multiple-challenge evaluation of reported reactions to monosodium glutamate », dans Journal of Allergy Clinical Immunology, vol. 106, no 5, 2000, p. 973-980
Voir aussi
Articles connexes
Liens externes
- (en) http://www.chemie.fu-berlin.de/chemistry/bio/aminoacid/glu_en.html
- (fr) Danger et description
- (fr) Texte législatif européen sur l'utilisation des additifs alimentaires 95/2 EC
 Portail de la biochimie
Portail de la biochimie Portail de la médecine
Portail de la médecine Portail de la chimie
Portail de la chimie Portail des neurosciences
Portail des neurosciences Portail de l’alimentation et de la gastronomie
Portail de l’alimentation et de la gastronomie


Wikimedia Foundation. 2010.