- Cycle De Krebs
-
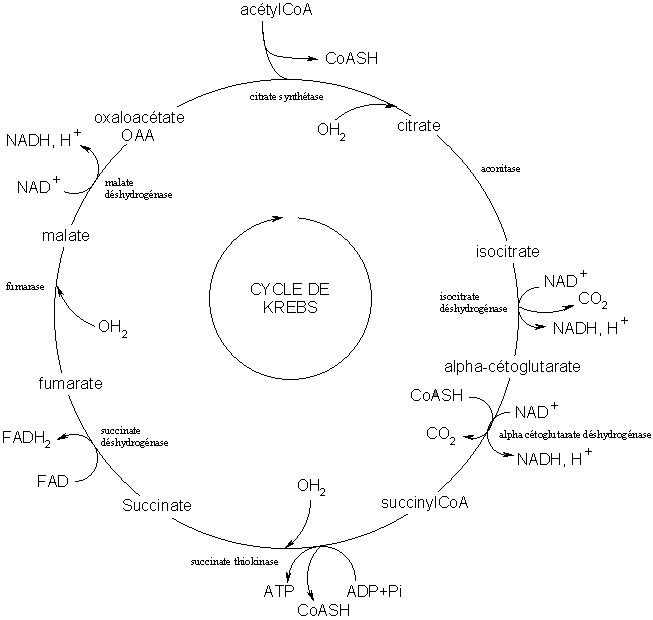
Cycle de Krebs
 Pour les articles homonymes, voir Krebs.
Pour les articles homonymes, voir Krebs.Le cycle de Krebs ou plus rarement mais plus justement cycle de Szent-Györgyi et Krebs ou cycle des acides tricarboxyliques ou encore cycle de l'acide citrique (citrate) est une série de réactions biochimiques dont la finalité est de produire des intermédiaires énergétiques qui serviront à la production d'ATP dans la chaîne respiratoire. Il s'agit d'un cycle car le dernier métabolite, l'acide oxaloacétique, est aussi impliqué dans la première réaction. Le cycle peut se résumer dans l'oxydation de 2 carbones en CO2 ; l'énergie dégagée par ces réactions génère du GTP (ou de l'ATP), des électrons, du NADH,H+ et du QH2, qui pourront être métabolisés par la chaine respiratoire pour former de l'ATP.
Point de convergence de plusieurs réactions de catabolismes du métabolisme cellulaire, il a été découvert par le biologiste Hans Adolf Krebs en 1937. Le cycle de Krebs peut aussi servir à la production d'intermédiaires utilisés pour certaines réactions d'anabolisme.
Il est le point final et commun du catabolisme des glucides (glycolyse, voie des pentoses phosphates ), lipides (hélice de Lynen) et acides aminés car tous ces catabolismes aboutissent à la formation d'acétyl-coenzyme A. L'acétyl-coenzyme A est une forme de transport des groupements acétyls qui proviennent du pyruvate. La première étape du cycle consiste à transférer ce groupement acétyl sur l'oxaloacétate pour former du citrate. Le reste du cycle consiste en des transformations catalysées. La dernière étape produit de l'oxaloacétate, qui peut ensuite réagir à nouveau dans la première étape un acétyl et recommencer le cycle. Il existe toutefois des réactions d'échappement au cycle de Krebs qui permettent d'utiliser certains intermédiaires pour d'autres fonctions cellulaires.
Le cycle de Krebs se déroule dans la matrice de la mitochondrie chez les eucaryotes ou dans le cytoplasme des bactéries, en conditions aérobies (présence d'oxygène). Les enzymes catalysant cette suite de réactions sont localisées dans la matrice mitochondriale (cytoplasme chez les bactéries) ou au niveau de la membrane interne mitochondriale (membrane interne chez les bactéries).
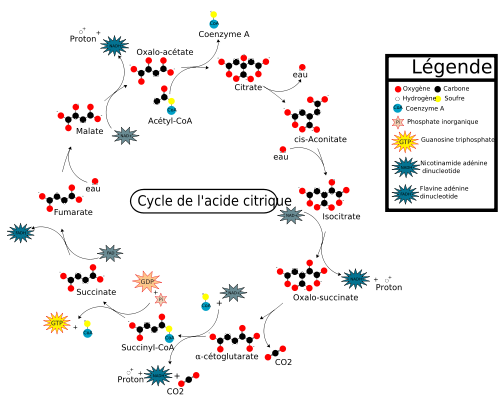 Cycle de Krebs ou cycle de l'acide citrique
Cycle de Krebs ou cycle de l'acide citrique
Avec la chaîne respiratoire qui réoxyde les coenzymes NADH et CoQH2 produits par le cycle, le cycle de Krebs est le processus ultime de dégradation des différents métabolites qui seront dégradés en dioxyde de carbone et en eau.
Étapes du cycle de Krebs
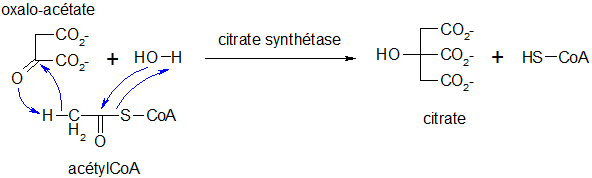
Synthèse du citrate
La réaction de condensation irréversible est catalysée par la citrate synthétase mais présente un intermédiaire transitoire : le cytroyl CoA.
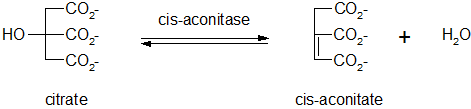
Déshydratation du citrate
Cette réaction de déshydratation réversible, catalysée par une lyase (cis-aconitase), produit du cis-aconitate (aussi appelé Z-aconitate). Bien que le citrate semble être symétrique, il a été prouvé que le départ d'eau a lieu entre les carbones de l'oxaloacétate.
Hydratation du cis-aconitate
Cette réaction est réversible et catalysée par la même enzyme qu'à l'étape précédente. L'addition d'eau sur la double liaison a lieu dans une position différente : c'est l'isocitrate.
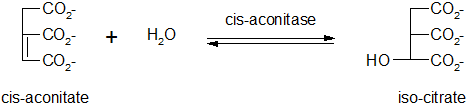
Oxydation de l’isocitrate
Cette réaction réversible est catalysée par une oxydoréductase : l’isocitrate déshydrogénase.
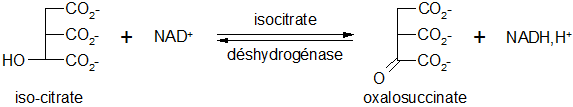
L’isocitrate déshydrogénase NAD+ dépendante exige également comme cofacteur des ions Mn2+ ou Mg2+.
Décarboxylation de l'oxalosuccinate
Il y a libération de dioxyde de carbone lors de cette réaction irréversible et spontanée, l'oxalosuccinate étant un composé instable.
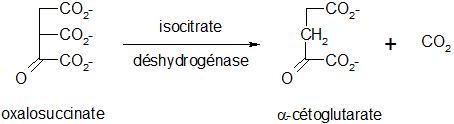
Décarboxylation oxydative de l'α-cétoglutarate
Cette réaction est la même que celle permettant le passage du pyruvate à l'acétylCoA. Le complexe enzymatique fait intervenir 5 coenzymes successifs : le thiamine pyrophosphate ou TPP), le lipoate, le NAD, le coenzyme A et le FAD. Cette réaction est irréversible.
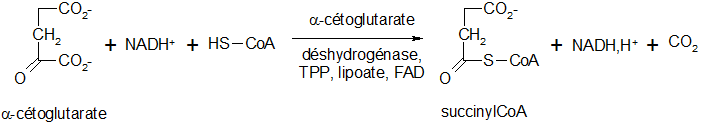
Formation du succinate
Lors de cette réaction, il y a transfert de l'énergie du succinylcoenzyme A (par sa liaison acylthioester) à la guanosine diphosphate. Cette réaction réversible est catalysée par une transférase, la succinate thiokinase. Formation d'une liaison P-O (GTP chez les animaux et ATP chez les végétaux)
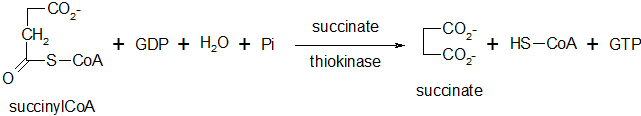
Oxydation du succinate
Cette réaction avec réduction de l'ubiquinone (Coenzyme Q10, CoQ) en ubiquinol (CoQH2) est catalysée par une enzyme flavoprotéique à FAD, inhibée par le malonate, la succinate déshydrogénase (oxydoréductase). Cette enzyme est en fait le complexe II de la chaîne respiratoire. Le FAD étant un groupe prosthétique lié de manière covalente dans l'enzyme, ne fait que transmettre les électrons et protons au vrai substrat CoQ.
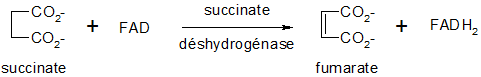
Hydratation du fumarate
Cette réaction d'addition est catalysée par une lyase, la fumarase.

Oxydation du malate : fermeture du cycle
Cette réaction referme le cycle. Il y a formation d'oxaloacétate, catalysée par le malate déshydrogénase (oxydoréductase).
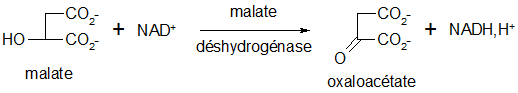
Moyen mnémotechnique
Si le citron isole l'acétone, le succinct succès fumera moins haut
(citrate, isocitrate, alphacétoglutarate, succinyl CoA, succinate, fumarate, malate, oxaloacétate)
Bilan du cycle de Krebs
Le cycle de Krebs est composé de 8 étapes, chacune étant catalysée par une enzyme spécifique. Au cours du cycle sont produites, à partir d'une mole de acide acétique et jusqu'au stade CO2 et H2O :
- 2 moles de CO2
- 3 moles de NADH,H+
- 1 mole de CoQH2
- 1 mole de GTP
On constate que le cycle de Krebs ne produit qu'un seul équivalent ATP (1 GTP), soit moins que la glycolyse (4 ATP pour une molécule de glucose dont 2 seront utilisés lors de la phase d'"activation" de la glycolyse [étape 1 et 3 qui correspondent à des étapes de phosphorylation]). L'essentiel de l'énergie chimique potentielle produite est sous forme de pouvoir réducteur(NADH,H+ et CoQH2). Ce pouvoir réducteur est ultérieurement utilisé dans la chaîne respiratoire mitochondriale pour produire 11 autres ATP (via un gradient protonique et une ATP synthase) que l'on attribue parfois, par erreur, au seul et unique cycle de Krebs.
Bilan du cycle de Krebs Consommation Production 1 Acétyl-CoA
3 NAD+
1 GDP
1 Pi
2 H2O
1 CoQ+1 CoA
3 (NADH, H+)
1 GTP
2 C02
CoQH2Ce qui correspond, au total, pour l'ensemble de la respiration aérobie (glycolyse, cycle de Krebs, réduction des coenzymes NAD et CoQ par la chaîne respiratoire) à 36 ou 38 ATP pour une molécule de glucose (cela dépend de la navette utilisée pour transporter le NAD de la glycolyse).
L’utilisation du glucose par respiration aérobie est plus énergétique que les fermentations.
En présence d'une grande quantité d'acétyl CoA, le cycle de Krebs peut être débordé en particulier chez les diabétiques ayant un déficit sévère en insuline ou après un jeûne prolongé, voir cétoacidose diabétique.
Régulation du cycle
Les étapes irréversibles du cycle de Krebs peuvent être régulées : étape de la citrate synthase, de l'isocitrate déshydrogénase et de α-cétoglutarate déshydrogénase.
- La citrate synthase est activée par l'ADP mais inhibée par le NADH, l'ATP et le citrate. Elle est donc respectivement inhibée par le pouvoir réducteur, la charge énergétique et le produit de la réaction qu'elle catalyse.
- L'isocitrate déshydrogénase est activée par le calcium, l'ADP et inhibée par le NADH et l'ATP.
- L'α-cétoglutarate déshydrogénase est activée par le calcium et inhibée par le NADH, l'ATP et son produit le succinyl-CoA.
Il y a donc une régulation selon la disponibilité du substrat, le pouvoir réducteur, la concentration en produit et la charge énergétique. On peut noter qu'il n'y a pas de régulation par covalence (phosphorylation des protéines).
Voir aussi
- Chaîne respiratoire : le devenir des NADH,H+
- Glycolyse
- Les principales voies du métabolisme (sur wikibooks)
Lien externe
- (en) Le cycle de Krebs
 Portail de la biologie
Portail de la biologie Portail de la biochimie
Portail de la biochimie Portail de la microbiologie
Portail de la microbiologie
Catégories : Métabolisme | Bioénergétique | Respiration cellulaire

Wikimedia Foundation. 2010.