- Fossiles
-
Fossile
 Ammonite fossilisée
Ammonite fossilisée
 Poisson fossilisé (Coelodus costai)
Poisson fossilisé (Coelodus costai)Un fossile (dérivé du substantif du verbe latin fodere : fossile, littéralement « qui est fouillé ») est le reste (coquille, os, dent, graine, feuilles...) ou le simple moulage d'un animal ou d'un végétal conservé dans une roche sédimentaire. Les fossiles et les processus de fossilisation sont étudiés principalement dans le cadre de la paléontologie.
Suivant les espèces et les périodes, les fossiles peuvent être de différentes qualités et plus ou moins abondants. Le processus de fossilisation est exceptionnel, et les témoignages que nous apportent les fossiles sur plus de trois milliards d'années d'évolution de la vie sur Terre sont encore lacunaires et le resteront certainement. Quand, pour les périodes récentes la fossilisation est inachevée, on parle de subfossilisation.
Sommaire
Éléments historiques
 Trilobite perminéralisé (Asaphus kowalewskii)Article détaillé : Histoire de la paléontologie.
Trilobite perminéralisé (Asaphus kowalewskii)Article détaillé : Histoire de la paléontologie.Depuis la préhistoire, l'homme a trouvé de nombreux fossiles, restes d'organismes pétrifiés par les minéraux qui les ont remplacés ou qui ont conservé leur enveloppe extérieure. L'homme primitif leur attachait une signification magique. Les auteurs de l'Antiquité, comme Aristote, les ont observés et, d'une façon générale, interprétés correctement. Toutefois, les deux idées essentielles à leur propos, i.e. leur origine organique et le fait qu'il s'agisse de témoignages que d'autres formes de vie ont existé avant l'Homme, n'ont pas été véritablement appréhendées avant le XVIIe siècle.
Le terme « fossile » est employé depuis Pline au Ier siècle[1], et son utilisation fut récupérée au XVIe siècle par Agricola, pour faire allusion à un corps enterré, que ce soient des restes d'organismes ou de minéraux intégrés dans les matériaux de la croûte terrestre. Cette situation curieuse a perduré jusqu'au début du siècle dernier. Lyell décrit les fossiles comme les restes d'organismes qui vivaient à une autre époque et actuellement intégrée au sein de roches sédimentaires. Cette définition reste valable, bien que désormais on accorde une plus grande ampleur au terme, en incluant les manifestations de l'activité de ces organismes tels que les excréments (coprolithes), les restes de constructions organiques, les traces d'empreintes, les impressions de parties du corps (ichnofossiles) ou même la dentelle, les squelettes ou les troncs, etc.
Les premiers progrès réels découlent d'une proposition explicitée au début du XVIIIe siècle : les terrains contenant des fossiles d'animaux ou végétaux marins devaient en toute logique avoir été recouverts par la mer, afin qu'ils s'y déposent sur le fond et s'enfoncent sur le lit sédimentaire. C'est la première fois que le fossile est envisagé comme indice stratigraphique. Toutefois, le poids de l'idée de génération spontanée, selon laquelle les espèces étaient apparues les unes après les autres et d'origine divine, empêcha une interprétation systématisée et approfondie des causes du renouvellement des espèces, tel que logiquement déduit de l'étude des fossiles.
À la suite de ces premiers progrès, l'idée d'une filiation entre les espèces fait son chemin, notamment par les écrits de Geoffroy Saint-Hilaire et Lamarck. S'opposent alors les visions créationniste, fixiste d'une part, transformiste, évolutionniste d'autre part. Le cœur de la controverse est atteint lorsqu'à la question des origines de la vie animale et végétale est mêlée celle des origines de l'Homme.
C'est également au XVIIIe siècle que trois grandes branches scindent la paléontologie — et subsistent encore à ce jour, sous la forme de spécialités disciplinaires : la paléontologie descriptive et comparative, de Cuvier ; la paléontologie évolutive, de Lamarck ; un peu plus tard, la paléontologie stratigraphique, d'Oppel et d'Orbigny. Suit la paléogéographie vers 1830.
De façon très nette, paléontologie et fossile se sont opposés de facto à une Église dogmatique, de la même manière que l'astronomie au Moyen Âge. Multidisciplinaire, organisée comme une enquête historique, l'étude des fossiles a également eu des implications importantes sur le rapport de l'Homme au temps, par exemple sur la question de l'âge de la Terre ou du vivant, ou encore sur la question des durées — l'unité temporelle de base d'un fossile est le million d'années, un laps de temps difficilement imaginable. Grâce à des progrès rapides et importants dans les techniques d'observation et d'investigation, la connaissance des fossiles et de la fossilisation au cours des temps géologiques a réalisé ses plus grandes avancées à partir du XIXe siècle.
Les fossiles sont examinés perpétuellement, à chaque fois qu'il est possible d'utiliser des techniques plus modernes. L'application de ces techniques implique parfois la modification des approches précédentes. Par exemple, à la suite d'un examen mené en 2006 avec des techniques de tomographie aux rayons X, il a été conclu que la famille qui contient les vers Markuelia avait une grande affinité avec les vers priapuliens, et est adjacent à la branche de l'évolution des Priapuliens, des Nématodes et des Arthropodes[2]. Le dernier fossile à avoir été découvert est celui du Lognkosauria, fossile découvert en 2007, son squelette était intact à 70% et est le 3e plus grand fossile au monde et aussi le plus complet d'entre eux.
Localisation des sites fossilifères
Certaines régions du globe sont particulièrement connues pour l'abondance de leurs fossiles. Ces sites fossilifères d'une qualité exceptionnelle portent le nom de Lagerstätten (littéralement lieu de repos ou d'emmagasinage, en allemand). Ces formations résultent probablement de l'enfouissement de carcasses dans un environnement anoxique avec très peu de bactéries, ce qui a ralenti le processus de décomposition. Sur l'échelle des temps géologiques, les lagerstätten s'étendent du Cambrien à nos jours.
Parmi ces sites, on trouve notamment les marnes jurassiques de la Voulte-sur-Rhône (conservation des parties molles de céphalopodes en trois dimensions), les schistes de Maotianshan en Chine et ceux de Burgess en Colombie-Britannique[3], le calcaire lithographique de Solnhofen en Bavière. Celui-ci détient, par exemple, un des magnifiques exemples d'Archéoptéryx. Ces gisements fossilifères sont tellement rares qu'à chacune de leur découverte ou redécouverte, ils ont bouleversé la vision de la progression de la vie.
Registre fossile
Le registre fossile correspond à l'ensemble des fossiles existant. Il s'agit d'un petit échantillon de la vie du passé, déformée et partiale[4]. Toutefois, il ne s'agit pas d'échantillons aléatoires. Toutes les investigations paléontologiques doivent tenir compte de ces aspects pour comprendre ce qui peut être obtenu grâce à l'utilisation de fossiles et ce qui ne peut pas l'être.
Rareté des fossiles

La fossilisation est un événement extrêmement rare. En effet, une grande partie de ce qui compose un être vivant a tendance à se décomposer relativement rapidement après la mort. Pour qu'un organisme soit fossilisé, les restes doivent normalement être recouverts par les sédiments dans les plus brefs délais. Cependant, il existe des exceptions à cette règle, comme pour un organisme congelé, desséché, ou immobilisé dans un environnement anoxique (sans oxygène). Il existe plusieurs types de fossiles et de fossilisation.
En raison de l'effet combiné des processus taphonomiques et du simple hasard mathématique, la fossilisation tend à favoriser les organismes composés de parties dures, celles qui sont particulièrement répandus sur le globe et ceux qui ont vécu pendant une longue période. D'autre part, il est très rare de trouver des fossiles de petits corps mous, d'organismes géographiquement limités ou éphémères géologiquement parlant, en raison de leur relative rareté et la faible probabilité de conservation. Les spécimens de grande taille (macrofossiles) sont plus souvent observés, déterrés et exposés, alors que les restes microscopiques (microfossiles) sont de loin les fossiles les plus courants.
Certains observateurs occasionnels furent perplexes devant la rareté des espèces transitionnelles dans le registre fossile. L'explication communément admise a été donnée par Darwin. Il a ainsi déclaré que « l'extrême imperfection du registre géologique », combiné à la courte durée et à l'aire de répartition géographique réduite des espèces de transition, conduisait à une faible probabilité de trouver beaucoup de ces fossiles. En d'autres termes, les conditions dans lesquelles se déroule la fossilisation sont assez rares et il est fort peu probable qu'un organisme donné se fossilise à sa mort. Eldredge et Gould ont développé une théorie de l'équilibre ponctué qui permet d'expliquer en partie le motif de stase et les apparitions soudaines dans le registre fossile.
Représentativité
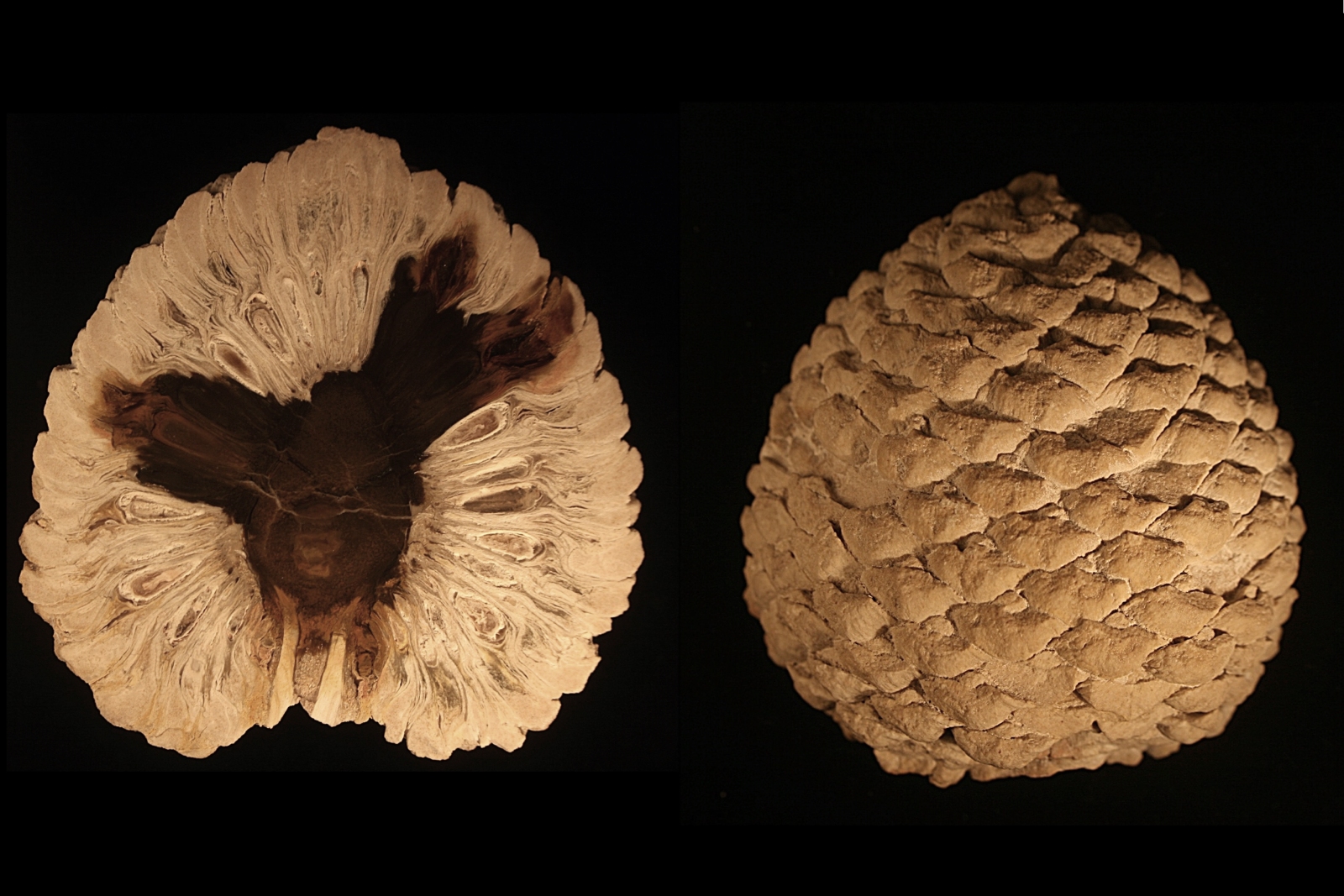
 Cône fossilisé de Araucaria sp., datant du Jurassique.
Cône fossilisé de Araucaria sp., datant du Jurassique. Fossile de triolobite Phacopide (Eldredgeops rana crassituberculata). Les yeux schizochroaux à facettes ont contribué à la formulation de la théorie de l'équilibre ponctué.
Fossile de triolobite Phacopide (Eldredgeops rana crassituberculata). Les yeux schizochroaux à facettes ont contribué à la formulation de la théorie de l'équilibre ponctué.Le nombre total d'espèces (y compris les plantes et les animaux) décrites et classées s'élève à 1,5 millions. Ce nombre continue d'augmenter, avec près de dix mille espèces d'insectes découvertes chaque année (il y a une grande diversité d'insectes avec 850 000 espèces connues). Les spécialistes estiment qu'il n'y a qu'une centaine d'espèces d'oiseaux connues à ce jour (il y a une faible diversité d'oiseaux avec seulement 8 600 espèces connues). Pour comparaison, on estime à près de 5 millions le nombre d'espèces vivantes possibles. On ne connaît environ que 300 000 espèces de fossiles, soit 20% du nombre d'espèces vivantes et moins de 6% du nombre probable. Le registre fossile s'étend d'il y a 3,5 milliards d'années jusqu'à aujourd'hui, mais 99% des fossiles ne remontent que jusqu'à 545 millions d'années. Ces chiffres sont énormes si l'on considère que le registre fossile correspond à une période correspondant à des centaines de millions d'années et que la faune et la flore vivant aujourd'hui ne représentent qu'un instantané à l'échelle des temps géologiques. Si la préservation des fossiles était bonne, on aurait d'avantage d'espèces fossiles que d'espèces vivantes à l'heure actuelle.
Il y a plusieurs explications possibles à la pauvreté relative des espèces fossiles. Dans un premier temps, il est possible que la biodiversité ait connue une forte croissance au fil du temps. Ceci conduit les experts à se demander s'il n'y aurait pas un manque de variété dans le passé géologique. Une autre explication peut provenir du fait que la diversité se mesure au nombre de taxons (espèces, genres, familles, ...) qui ont vécu au cours d'un intervalle de temps défini et que chaque ère géologique ne présente peut-être pas la même diversité. Il ne faut pas oublier par ailleurs que certaines des parties de la colonne géologique sont mieux connues que d'autres. Par exemple, le nombre de paléontologues travaillant sur le Précambrien et le Paléozoïque ne représente qu'un très faible pourcentage des chercheurs, alors que le travail sur ces périodes est considérable. Enfin, il est probable qu'il faille aussi tenir compte du fait que les roches récentes se trouvent dans les strates supérieures, ce qui expliquerait pourquoi on les retrouve plus facilement.
Tout donne à penser que la diversité actuelle peut ne pas être significativement plus élevée que la moyenne, pour les ères géologiques remontant jusqu'au Cambrien. Par conséquent, le faible nombre d'espèces fossiles ne peut être expliqué de façon satisfaisante par l'idée que la diversité croît avec les progrès de l'évolution. Les espèces disparaissent et sont remplacées par de nouvelles au cours des temps géologiques. Il a été suggéré qu'il faudrait un délai de 12 millions d'années pour opérer un remplacement complet de toutes les espèces. La durée de chaque biochrones se situe entre 0,5 et 5 millions d'années (2,75 Ma en moyenne). Enfin, en conclusion, la quantité d'espèces fossiles est estimé à :
Nb d'espèces par biochrone x Nb de biochrones = 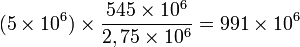
Les types de fossiles
 Tronc pétrifié d'Araucarioxylon arizonicum.
Tronc pétrifié d'Araucarioxylon arizonicum.Les fossiles les plus anciens sont les stromatolithes, qui sont composées de roches créé par la sédimentation de substances (telles que le carbonate de calcium) grâce à l'activité bactérienne[5]. Celle-ci a été découverte à travers l'étude des stromatolithes actuellement produit par des tapis microbiens. La formation Gunflint contient de nombreux microfossiles, largement acceptés comme étant des restes microbiens[6].
Il existe de nombreux types de fossiles. Les plus courants sont les restes d'escargots ou des os transformés en pierre. Beaucoup d'entre eux montrent tous les détails originaux de la coquille ou de l'os. Les pores et autres petits espaces de leur structure sont remplis de minéraux. Les minéraux, tels que la calcite (carbonate de calcium), sont des composés chimiques qui ont été dissous dans l'eau. Lorsque l'escargot (ou l'os) passe à travers le sable ou la boue, des minéraux se déposent dans les espaces de sa structure. C'est pourquoi les fossiles sont si lourds. D'autres fossiles ont pu perdre toutes les marques de sa structure originelle. Par exemple, un escargot dont la coquille était à l'origine composée de calcite peut se dissoudre complètement après avoir été enterré. L'impression laissée dans la roche peut alors se remplir par d'autres matériaux et forme une réplique exacte de l'escargot. Dans d'autres cas, l'escargot est dissous et il ne reste alors plus qu'un trou dans la pierre, une sorte de moule que les paléontologues peuvent remplir de plâtre pour découvrir à quoi ressemblait l'animal.
Généralement, les fossiles ne montrent seulement que les parties rigides de l'animal ou du végétal : le tronc d'un arbre, la coquille d'un escargot ou les os d'un dinosaure. Certains fossiles sont plus complets. Si une plante ou un animal reste enfoui dans un type spécial de boue, qui ne contient pas d'oxygène, certaines des parties molles peuvent également être préservées en fossiles.
Les plus spectaculaires « fossiles parfaits » sont ceux des mammouths laineux qui ont été retrouvés dans un sol gelé[7]. La viande était tellement gelée, qu'elle aurait pu être consommée, même après 20 000 ans. Par convention, on estime que les plus récents fossiles d'organismes vivaient à la fin de la dernière glaciation quaternaire (Würm), c'est-à-dire il y a quelques 13 000 ans environ. Le autres, qui datent d'une époque plus récente (néolithique, âge des métaux, etc.) sont généralement considérés comme des sous-fossiles.
Enfin, il faut aussi tenir compte des produits chimiques inclus dans les sédiments qui indiquent l'existence de certains organismes qui les sécrétaient ou en était fait. Ils représentent l'extrême limite de la notion de fossiles (marqueurs biologiques ou fossiles chimiques).
Ichnofossile
 Asteriacites, ichnogenre attribué à des étoiles de mer (groupe Cubichnia).
Asteriacites, ichnogenre attribué à des étoiles de mer (groupe Cubichnia).Les ichnofossiles sont les restes de dépôts, d'empreintes, d'œuf, de nids, de bioérosion ou de n'importe quel autre type d'impression. Ils sont l'objet d'étude de la Paléoichnologie. Les ichnofossiles présentent des caractéristiques qui les rendent facilement identifiables et permettent sa classification comme parataxons : ichnogenres et ichnoespèces. Les ichnotaxons sont des classes de pistes de fossiles regroupés suivant leurs propriétés communes : géométrie, structure, taille, type de substrat et fonctionnalité. Bien que parfois un diagnostic de l'espèce productrice de l'ichnofossile peut s'avérer ambigu, en général, il est possible de déduire au moins le groupe biologique ou le taxon supérieur auquel il appartenait.
Le terme ichnofaciès fait référence à l'association caractéristique des traces fossiles qui reflètent les conditions environnementales telles que la bathymétrie, la salinité et le type de substrat[8]. Les traces et les empreintes d'invertébrés marins constituent d'excellents indicateurs paléoécologiques. En effet, elles sont le résultat de l'activité de ces organismes, en liaison avec leur environnement spécifique (nature du substrat et conditions du milieu aquatique : salinité, température, bathymétrie). En particulier, la profondeur de la mer conditionne le type d'organismes qui vont s'y développer et, par conséquent, il n'est pas surprenant que l'on puisse distinguer une gamme d'ichnofaciès suivant la bathymétrie, dont la nomenclature due à Seilacher fait référence aux types de pistes les plus fréquentes et les plus caractéristiques[9].
Microfossile
 Microfossiles de sédiments marins, en provenance de l'Antarctique.
Microfossiles de sédiments marins, en provenance de l'Antarctique.Le microfossile désigne une plante ou un animal fossilisé dont la taille est trop petite pour être analysé à l'œil nu. On applique communément un seuil de taille pour distinguer les microfossiles des macrofossiles, 1 mm, mais il ne s'agit que d'un guide approximatif. Les microfossiles peuvent être soit des organismes complets (ou quasi-complets), comme le plancton foraminifère ou coccolithophoridé, soit des parties isolées de plus gros animaux ou de plantes, comme des petites dents ou des spores. Les microfossiles sont d'une grande importance pour les biostratigraphes qui les utilisent pour aider à la corrélation des roches critiques, car ce sont des réservoirs d'informations paléoclimatiques.
En règle générale, on utilise deux éléments pour différencier les microfossiles eucaryotes ou procaryotes : la taille (les eucaryotes sont généralement plus grand que les procaryotes) et la complexité des formes (les eucaryotes ont une structures plus complexes en raison de leur cytosquelette).
Résine fossile
La résine fossile (aussi appelé ambre) est un polymère naturel que l'on rencontre dans plusieurs types de strates différentes, partout dans le monde. Il s'agit de résine fossilisée provenant de la sève des arbres et datant pour la plupart du Tertiaire (2-5 millions d'années), voire du Trias (200 millions d'années). On le trouve généralement sous forme de pierres jaune-orangées.
On estime que la résine est une adaptation évolutive des arbres pour la protection contre les insectes et l'étanchéité des blessures causées par les éléments. La résine fossile contient souvent d'autres fossiles, appelés inclusions, qui ont été capturés par la résine collante. Il s'agit notamment de bactéries, de champignons, de plantes ou d'animaux. Les inclusions animales sont généralement de petits invertébrés, principalement les arthropodes comme les insectes et les araignées, et très rarement des vertébrés comme un petit lézard.
Fossile vivant
 Nautile.
Nautile.Un fossile vivant est un terme informel qui est utilisé pour qualifier une espèce vivante qui présente des ressemblances morphologiques avec des fossiles retrouvés. En règle générale, il s'agit d'espèces qui ont très peu évolué, du point de vue morphologique, au cours du temps. On parle ainsi plutôt de panchronisme. Les brachiopodes sont de parfaits exemples de fossiles vivants. On peut aussi citer les lingulata (dont des fossiles datant de 200 millions d'années ont été retrouvés) ou les cœlacanthes. Ce fut un grande surprise quand ces derniers ont été découverts le long des côtes africaines, en 1938, alors qu'on les pensait disparus depuis 70 millions d'années.
Ce peut donc être une espèce ou un taxon connu uniquement sous forme de fossiles avant que des représentants vivants ne soient découverts (cœlacanthe, monoplacophore primitif, ginkgo biloba, ...), une espèce vivante sans aucun proche parent (cagou de Nouvelle-Calédonie, caurale soleil, ...) ou un petit groupe d'espèces étroitement liées sans proche parent (stromatolithe, lingulata, nautile, psilotum, limule, sphénodon, ...).
Pseudo-fossile
Un pseudo-fossile est un motif que l'on peut observer sur une roche mais qui est le résultat d'un processus géologique, plus que biologique. Ils peuvent facilement être confondus avec de vrais fossiles. Certains pseudo-fossiles, tels que les dendrites, sont formés par des fissures qui se produisent naturellement dans la roche et qui se remplissent par percolation des minéraux. Parmi les autres types de pseudo-fossiles, on peut également citer les reins de minerai (formes rondes dans le minerai de fer) ou l'agate mousse, qui ressemblent à de la mousse ou des feuilles coincées dans une agate. Des concrétions, sphériques ou ovoïdes, en forme de nodules dans certaines couches sédimentaires ont déjà été considérées comme étant des œufs de dinosaures et sont également souvent confondus avec des fossiles.
Les erreurs d'interprétation dues aux pseudo-fossiles ont généré plusieurs controverses tout au long de l'histoire de la paléontologie. Ainsi, en 2003, un groupe de géologues espagnols a remis en question l'authenticité des fossiles de Warrawoona. Selon William Schopf, il s'agirait de cyanobactéries qui seraient les premières traces de vie sur la Terre, il y 3,5 milliard d'années. Le groupe espagnol affirme, pour sa part, qu'un sel de baryum et un silicate, placé dans un environnement alcalin, à température et pression ambiante, peut produire des structures filamenteuses similaires[10].
La fossilisation
La fossilisation des êtres vivants est en général un processus de minéralisation (remplacement des tissus vivants par des substances minérales) dans de la roche sédimentaire qui est la roche par excellence pour la conservation de fossiles. Dans des cas plutôt rares, on peut avoir une conservation de la matière organique (mammouth dans le pergélisol, momification dans du bitume, la diatomite (roche siliceuse), inclusion dans de l'ambre). Dans d'autres cas, ce ne sont que des traces d'activité biologique qui sont conservées (paléoichnologie).
Processus de décomposition

La capacité de conservation des fossiles est en grande partie due au processus de décomposition des organismes. Celui-ci explique pourquoi il est rare de retrouver des fossiles des parties molles organiques (60% des individus d'une communauté marine sont uniquement composés de parties molles). La présence des parties molles est alors le résultats de conditions sédimentologique et diagénétiques exceptionnelles.
La biodégradation s'effectue de manière rapide et efficace grâce à une décomposition aérobie. Ainsi, il est nécessaire d'avoir un environnement anoxique pour pouvoir préserver des organismes légèrement minéralisés et composés de parties molles. Ceci exige une concentration élevée d'oxygène (106 moles pour 1 mole de carbone organique) :
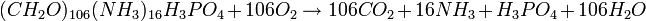
La décomposition est la source principale de perte de données dans le registre fossile et la minéralisation est le seul moyen de la freiner. Les tissus peuvent se conserver sous la forme de perminéralisations (déchets organiques altérés) ou, quand la détérioration est prolongée, sous la forme d'empreintes. Si la décomposition est plus importante que la minéralisation, les tissus sont détruits et seuls les matériaux réfractaires (chitine, lignine ou cellulose) sont conservés.
La décomposition dans le registre fossile se caractérise à trois niveaux. Dans un premier temps, il s'agit d'identifier la décomposition et la perte d'information sur la structure de l'organisme fossilisé. Ensuite, il faut reconnaître les minéraux particuliers et les marqueurs géochimiques associés aux régimes particuliers de décomposition. Enfin, il faut garantir la préservation des microbes fossiles impliqués dans le processus.
La majeure partie de la matière organique se recycle pour la majeure partie à l'intérieur de la colonne d'eau, en particulier dans la zone euphotique. Une petite proportion de la matière organique produite sert à la formation des sédiments adjacents et est affectée par les modifications du flux organique (biostratinomique) telles que la photo-oxydation, l'activité microbienne et les organismes détritivores.
Processus de diagenèse fossile
 Catellocaula vallata, organisme à corp mou (trous en forme d'étoile), préservé par bioimurration dans un squelette de bryozoaire, datant de l'Ordovicien Supérieur[11].
Catellocaula vallata, organisme à corp mou (trous en forme d'étoile), préservé par bioimurration dans un squelette de bryozoaire, datant de l'Ordovicien Supérieur[11].En plus des lipides, la matière organique comprend également des biopolymères, comme les glucides, les protéines, la lignine et la chitine, dont certains seront utilisés pour sa consommation ou modifié par les organismes benthiques et les micro-organismes. Ceux qui ne sont pas utilisés pourront subir une polycondensation qui conduira à la formation de géopolymères qui s'intègreront au proto-kérogène (précurseur du kérogène). Lors de l'enfouissement des sédiments, la condensation s'accroit et l'insolubilité produit la lente conversion diagénétique du kérogène, constituant principal de la matière organique dans les sédiments anciens.
On trouve de grandes quantités de molécules organiques dans les sédiments et les roches sédimentaires. On les qualifie de marqueurs biologiques ou de « biomarqueurs ». Leur étude et leur identification nécessitent des techniques avancées d'investigation et d'analyse. Ces marqueurs conservent un registre très détaillé de l'activité biologiques passée et ils sont liés aux molécules organiques actuelles. On trouve autant de sources possibles de marqueurs biologiques dans les échantillons que de molécules dans un organisme.
Une roche mère est un volume rocheux qui a généré et expulsé des hydrocarbures en quantité suffisante pour former une accumulation de pétrole et de gaz. La plupart des roches mères potentielles contiennent entre 0,8 et 2% de carbone organiques. Il est courramment admit, comme limite basse, un pourcentage de 0,4% en volume de carbone organique pour la production d'hydrocarbures. Toutefois, la génération est plus efficace avec un pourcentage supérieur à 5-10%. La nature des hydrocarbures générés dépend essentiellement de la composition du kérogène, qui peut être composé de deux types de matières organiques : les débris de plantes terrestres - les sédiments libèrent alors du gaz - ou d'organismes aquatiques, comme les algues, le phytoplancton, le zooplancton - auquel cas ils forment alors du pétrole (si la maturation est suffisante).
Processus de destruction physico-chimique
 Tigre à dents de sabre (Smilodon californicus) fossilisé.
Tigre à dents de sabre (Smilodon californicus) fossilisé.La durabilité des squelettes dépend de leur résistance à la rupture et à la destruction par des agents chimiques, physiques et biotiques. Ces processus destructeurs peuvent être divisées en cinq catégories qui suivent plus ou moins l'ordre séquentiel : la désarticulation, la fragmentation, l'abrasion, la bioérosion et la corrosion/dissolution.
La désarticulation correspond à la désintégration de squelettes composés de plusieurs éléments le long des jointures ou des articulations préexistantes. Ce phénomène peut également se produire avant même la mort, comme la mue ou l'exuvie chez les arthropodes). Cette décomposition détruit les ligaments reliant les ossicules d'échinodermes en quelques heures ou quelques jours après la mort. Les ligaments, comme ceux des moules, composés de conchyoline, sont plus résistantes et peuvent rester intacts pendant des mois, en dépit de la fragmentation de la coquille.
La fragmentation se produit lors d'un impact par des objets physiques et par des agents biotiques, tels que les prédateurs ou les nécrophages. Certaines formes de rupture permettent d'identifier le prédateur. Les coquilles ont tendance à se briser le long de lignes de faiblesse pré-existantes, telles que les lignes de croissance ou d'ornementation. La résistance à la fragmentation dépend de plusieurs facteurs : la morphologie du squelette, la composition et la microstructure (notamment épaisseur et pourcentage de matière organique).
L'abrasion est le résultat du polissage et du concassage des éléments du squelette, qui produit un arrondissement et une perte des détails de la surface. Il y a eu des études semi-quantitatives sur les proportions de l'abrasion, en introduisant des coquilles dans un tambour rotatif, rempli de gravier siliceux[12]. Le degré d'intensité est liée à plusieurs facteurs : l'énergie du milieu, le temps d'exposition, la taille de la particule abrasive et la microstructure du squelette.
 Fossile d'Eurypterus remipes.
Fossile d'Eurypterus remipes.La bioérosion ne peut se produire que si elle est associée à des fossiles reconnaissables, tels que les éponges Cliona ou les algues endolithiques. Son action destructrice est très importante dans les milieux marins peu profonds, où on peut observer une perte de masse allant de 16 à 20% dans les coquilles des mollusques actuels. Aucune étude ne montre toutefois si les proportions étaient les mêmes au Paléozoïque, quand les éponges cliona étaient moins abondantes.
La corrosion et la dissolution est le résultat de l'instabilité chimique des minéraux qui se trouvent dans la colonne d'eau et dans les pores des sédiments. La dissolution commence à l'interface sédiment-eau avant de continuer vers l'intérieur du sédiment. La bioturbation des sédiments favorise normalement la dissolution grâce à l'introduction d'eau de mer à l'intérieur du sédiment, ce qui permet également l'oxydation des sulfures.
Dans la pratique, il est difficile de distinguer les effets de l'abrasion mécanique, de la bioérosion et de la corrosion. Certains auteurs ont ainsi proposer le terme de corrasion pour indiquer l'état général des coquilles, comme le résultat d'une combinaison de ces processus. Le grade de corrasion est proportionnel à un indice général du temps durant lequel les restes ont été exposés à ces trois processus.
Diagenèse fossile
La compréhension des processus diagénétiques est essentielle pour l'interprétation correcte de la minéralogie originale, de la structure des squelettes et des coquilles, de leurs affinités taxonomiques et de la paléoécologie. L'un des problèmes auxquels nous sommes confrontés est très souvent de déduire ce qu'a été la minéralogie originale de groupes disparus (coraux bruts, archéocyathes, stromatopores...). La transition vers un état de fossile dépend surtout de la composition du squelette.
Nodule de carbonate et de calcaire lithographique
 Rusophycus, que l'on assimile généralement à des restes de trilobites.
Rusophycus, que l'on assimile généralement à des restes de trilobites.La préservation des parties molles est souvent associée à la précipitation des carbonates sous la forme de nodules stratifiés, comme pour le calcaire lithographique. Les nodules de carbonates sont composés de calcite ou de sidérite, et associés aux sédiments argileux riches en micro-organismes. Ils contiennent souvent des fossiles conservés dans leurs trois dimensions, et contiennent parfois même les restes fossilisés des parties molles. Leur taille varie entre 10 et 30 centimètres, même si certains atteignant les 10 mètres ont été retrouvés (dont un Plésiosaure complet). Le contenu de micro-organismes et leur décomposition sont les principaux facteurs qui contrôlent le degré d'anoxie, le potentiel d'oxydo-réduction et le pH. En présence d'oxygène, la respiration microbienne produit du CO2 qui s'accumule dans l'eau interstitielle des sédiments, favorisant la dissolution des carbonates :
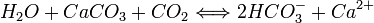
En l'absence d'oxygène, les bactéries utilisent une série d'oxydants alternatifs dans le processus de la respiration (Mn, NO3-, Fe ou SO42-). Une fois que tous les oxydants ont disparu, la fermentation devient la réaction dominante et la production de méthane augmente. Le calcaire lithographique se forme dans un environnement marin ou lacustre et présente sous formes de fines bandes à grain fin. On peut citer comme exemple, le célèbre calcaire de Solnhofen datant du Jurassique et contenant des fossiles d'Archaeopteryx. Les dépôts de carbonate peuvent provenir de sources biogéniques (comme les algues calcaires) ou d'un précipité chimique.
Squelette d'aragonite
Normalement, l'aragonite se transforme en calcite à travers un processus de dissolution ou de calcification. Si les eaux du gué ne sont pas saturés en carbonates, il se produit une dissolution totale du squelette et des chairs par la calcite. L'espace vide reproduit le moule d'une coquille vide et la structure de cette dernière n'est pas conservée. Il peut se former des druses avec des cristaux dirigés vers le centre. La durée de ce processus est variable. Dans le cas de la calcification, le squelette des coquilles conserve son ancienne structure (en couches ou lamelles). Il se peut Même que soient préservés les cristaux d'aragonite, ce qui nous donnent des renseignements très utiles. Ce remplacement se fait progressivement et respecte la structure d'origine.
Squelette de calcite
 Moulage calcitique d'une coquille de bivalve.
Moulage calcitique d'une coquille de bivalve.En général, les squelettes fossiles qui étaient constitués de calcite, conservent souvent leur composition originale (à moins qu'ils ne se silicifient ou ne se dolomitisent). La teneur en magnésium a tendance à diminuer, de sorte qu'il puisse y avoir une altération diagénique, soit à forte, soit à faible teneur en calcite. Il existe des techniques spéciales, telles que la cathodoluminescence, pour déterminer son contenu original à partir des zones qui ont conservé leur composition originale.
Squelette calcaire
Les squelettes de carbonate de calcium peuvent se transformer en apatite sans modification de la morphologie externe. Dans les milieux naturels, cette modification diagénique est associée à des dépôts de phosphate. La transformation bactérienne des organismes calcaires en apatite a été démontrée en laboratoire. Ces observations et ces expériences suggèrent, dans un premier temps, que le phosphore nécessaire pour remplacer le carbonate par de l'apatite provient des micro-organismes des sédiments. Par ailleurs, il semble que les micro-organismes (bactéries, algues, champignons) favorisent la décomposition, en libérant des ions phosphates et en acidifiant l'eau interstitielle des sédiments. Cette acidification, qui peut être très localisée, favorise la dissolution des carbonates. Le phosphate libéré se combine avec le calcium pour former de l'apatite, préférentiellement à l'interface entre le carbone et le micro-organisme remplaçant le carbonate dissous. Ce remplacement préserve l'apparence originale de la coquille et le fluor joue un rôle important en ce qui concerne la composition finale en carbonate-fluor-apatite.
Squelette de silice

Le phosphatisation de la silice primaire apparaît aussi sur certains squelettes de radiolaires, bien que ce processus ne soit pas encore bien connu à l'heure actuelle. L'examen microscopique d'échantillons de phosphorites montre que de nombreux micro-organismes sans carapace minérale (algues, champignons, bactéries) se minéralisent comme l'apatite, bien qu'ils n'aient aucun précurseur minéral. Un exemple bien connu est le coprolithe phosphaté, où la matière organique est elle-même remplacée par de l'apatite qui conserve la forme exacte de l'objet. La phosphatisation des parties molles est également fréquent, notamment chez de nombreux arthropodes (copépodes, ostracodes) où des nodules calcaires et phosphatés apparaissent au sein de calcaire nodulaire ou de coprolithes de grands vertébrés.
Des études sur les phosphorites et sur la synthèse expérimentale de l'apatite ont abouti à une estimation des conditions probables de fossilisation de l'apatite. En raison de son besoin de stabilité, l'apatite se forme de préférence dans un environnement déficient en oxygène, parfois même dans des conditions totalement réductrices, comme l'indique la présence fréquente de pyrite à proximité. Cet environnement est atteint facilement dans les milieux où l'on trouve beaucoup de matière organique qui est la principale source de phosphore.
La silice peut remplacer la calcite et l'aragonite des coques et perminéraliser le bois. Il peut également se former des nodules et des couches de silex, en remplaçant les sédiments carbonés, en précipitant directement ou en remplissant les fossiles ou les inclusions. La coque peut alors être remplacées par une croûte blanche granuleuse, par une couche finement granuleuse ou par des anneaux concentriques de silice.
Fossile pyritisé

La pyrite sédimentaire est une composante mineure des sédiments clastiques marins. Les études actuelles sur les sédiments ont montré que la formation de la pyrite authigénique a lieu au tout début de la diagenèse, à quelques centimètres au-dessous de l'interface eau-sédiments. Une augmentation du nombre de micro-organismes et/ou de la profondeur d'enfouissement empêche la diffusion de l'oxygène dans les sédiments et les micro-organismes sont obligés de respirer en anaérobie. La minéralisation empêche la perte d'information relative à la décomposition de macro-organismes et la précipitation de la pyrite, au début de la diagenèse, est un moyen important pour la préservation des fossiles. Dans les tissus mous, comme les muscles et la chitine, il peut se produire un pyritisation au début de la diagenèse. Lorsque la décomposition est plus avancée (mais avant que ne se produise la formation de la pyrite), les tissus mous seront détruits et seuls les composés biologiques résistants (appelés réfractaires), comme la cellulose et la lignine, sont préservés. Les parties biogéniques dures, telles que les coquilles (composées de carbonate de calcium et de magnésium) et les os (phosphate de calcium) sont quelques-unes des structures biologiques les plus résistantes à la décomposition. Sur les deux, le carbonate de calcium est le plus instable et il est donc plus probable qu'il soit remplacé par la pyrite.
La formation de la pyrite est contrôlée par la concentration en carbone organique, en sulfate et en minéraux détritiques ferreux. Dans un environnement marin normal, les minéraux ferreux et les sulfates sont présents en abondance et la formation de pyrite est contrôlée par l'approvisionnement en carbone organique. Toutefois, dans les milieux en eau douce, la formation de pyrite est très limitée par la faible concentration en sulfates.
Plante fossile

Les différentes parties des plantes (branches, racines, feuilles, pollen, fruits, graines) se détachent pour certaines au cours de leur vie, et pour les autres après leur mort. Une bonne compréhension des processus de dispersion qui affectent la dispersion de ces parties est très important pour interpréter correctement les associations paléofloristiques. Les études sur la dispersion des feuilles par le vent montrent qu'elle dépend de leur poids et de leur forme. Les débris végétaux se conservent soit par préservation du matériel original, soit pas carbonisation, soit par perminéralisation.
La perminéralisation se produit après l'enfouissement, quand les espaces vides situés à l'intérieur d'un organisme (espaces remplis de liquide ou de gaz, quand il est en vie) se remplissent d'eaux souterraines riches en minéraux et que ces minéraux précipitent en comblant les espaces vides. Ce processus ne peut se produire que dans de très petits espaces, notamment au sein de la paroi cellulaire d'une cellule végétale. Une perminéralisation à petite échelle peut produire des fossiles avec de très nombreux détails. Pour que la perminéralisation se produise, il faut que l'organisme soit recouvert par les sédiments peu de temps après la mort ou peu après le début du processus de décomposition. La vitesse de dégradation des restes, une fois recouverts, détermine les futurs détails du fossile. Ainsi, alors que certains fossiles ne comprennent que des ossements ou des dents, d'autres contiennent des traces fossiles de peau, de plumes ou même de tissus mous.
Importance scientifique
 Insecte (Sciaridae) piégé dans de l'ambre.
Insecte (Sciaridae) piégé dans de l'ambre.De tout temps, les fossiles ont intrigué les hommes qui, suivant les époques, leur ont donné différentes significations : talismans, restes de géants, objets maléfiques, animaux disparus lors du Déluge. Ce n'est qu'au XIXe siècle, avec les travaux de Charles Lyell, de Jean-Baptiste de Lamarck, puis de Charles Darwin et les théories de l'évolution, puis de la théorie de la tectonique des plaques, formulée par Alfred Wegener en 1915, que se met en place le cadre théorique moderne dans lequel sont étudiés les fossiles.
Pour le grand public, les fossiles sont surtout connus grâce à quelques familles caractéristiques comme les ammonites, sortes de céphalopodes marins, les trilobites de la famille des arthropodes, les oursins ou enfin les végétaux fossiles conservés dans le charbon (fougères, prêles, etc.).
L'extraction d'ADN fossile a récemment été développée grâce à l'amplification permise par la réaction en chaîne par polymérase. Depuis la fin des années 1990, les connaissances sur ces techniques se sont améliorées[13]. L'une des techniques proposée consiste à extraire de l'ADN de l'ambre. Bien que cette idée soit actuellement irréalisable, l'imagination populaire a été nourrie à travers le livre et le film « Jurassic Park ». Dans ce livre, on suggére que les moustiques piégés dans l'ambre pourraient avoir conservé intact l'ADN d'autres animaux, tels que les dinosaures. On a cru parvenir à de bons résultats grâce à cette méthode et plusieurs études font ainsi état d'ADN datant de plus de 100 millions d'années [14], mais des études plus récentes (quoique moins médiatisées) ont montré que ces résultats n'étaient absolument pas concluants et provenaient la plupart du temps de contaminations actuelles [15].
De l'ADN peut également être extrait de cristaux présents dans les os fossilisés. Les scientifiques ont montré que parfois des cristaux se formaient à l'intérieur des os, et que ces cristaux pouvaient contenir des traces d'ADN.
L'importance de l'étude de la formation des fossiles a conduit à la fondation d'une nouvelle discipline, la taphonomie.
Annexes
Notes et références
- (es) Cet article est partiellement ou en totalité issu d’une traduction de l’article de Wikipédia en espagnol intitulé « Fósil ».
- ↑ (es)Museo Geológico Virtual de Venezuela, « Glosario de Fósiles ». Consulté le 23 décembre 2007
- ↑ (en)PCJ Donoghue, et al., Synchrotron X-ray tomographic microscopy of fossil embryos, vol. 442, Nature, 2006, p. 680-683
- ↑ (en)The Burgess Shale, Universidad de Berkeley. Consulté le 22 décembre 2007
- ↑ (en)S.R. Fernández-López, La naturaleza del registro fósil y el análisis de las extinciones, vol. 51, Coloquios de Paleontología, 2000, p. 267-280
- ↑ (en)Stromatolites, the Oldest Fossils. Consulté le 4 mars 2007
- ↑ (en)A.H. Knoll, et al, New organisms from the Aphebian Gunflint Iron Formation, vol. 52, Journal of Paleontology, 1978, p. 1074-1082
- ↑ (es)10.000 años muy bien llevados. Consulté le 22 décembre 2007
- ↑ (en)R.G. Bromley, Trace fossils. Biology and Taphonomy, Unwin Hyman, Londres, 1990, 280 p.
- ↑ (de)A. Seilacher, Studien zur palichnologie. I. über die methoden der palichnologie. Neues Jahrb, vol. 96, Geologie Paläontologie Abhandlungen, 1953, p. 421-452
- ↑ (es)Un español replantea la forma de detectar la vida primitiva. Consulté le 27 décembre 2007
- ↑ (en)T.J. Palmer et M.A. Wilson, Parasitism of Ordovician bryozoans and the origin of pseudoborings, vol. 31, Palaeontology, 1988, p. 939-949
- ↑ (en)K.E. Chave, Skeletal durability and preservation, John Wiley and Sons, New York, 1964, p. 377-387
- ↑ (en)boffr D. Grimaldi, DNA sequences from a fossil termite in Oligo-Miocene amber and their phylogenetic implications, Science, 1992, p. 1993-1936
- ↑ (en)R.J. Cano et al., Amplification and sequencing of DNA from a 120–135-million-year-old weevil, vol. 363, Nature, 1993, p. 536-538
- ↑ (en)M. B. Hebsgaard et al., Geologically ancient DNA: fact or artefact?, vol. 13, Trends in Microbiology, 2005, p. 212-220
Articles connexes
Bibliographie
- (en)D.V. Ager, Principles of paleoecology, McGraw-Hill, 1963, 371 p.
- (en)P.A. Allison et D.E.G. Briggs, Exceptional fossil record: distribution of soft tissue preservation through the Phanerozoic, vol. 21, Geology, 1993, p. 527-530
- (es)Mauricio Antón, El secreto de los fósiles, Aguilar, 2006 (ISBN 978-84-03-09762-9)
- (it)P. Arduini et G. Teruzzi, Fossili, Arnaldo Mondadori Editore, Milan, 1986
- (en)R.L. Bates et J.A. Jackson, Dictionary of geological terms (Third Edition), Anchor Book (Prepared by The American Geological Institute), New York, 1984, 571 p.
- (en)R.L. Bates et J.A. Jackson, Glossary of Geology (Third Edition), American Geological Institute, 1987, 788 p.
- (en)D.E.G. Briggs, Extraordinary fossils, vol. 79, American Scientist, 1991, p. 130-141
- (en)R.G. Bromley, Trace fossils, biology and taphonomy, Unwin Hyman, Londres, 1990, 280 p.
- (es)R. Candel et al., Historia Natural: Vida de los animales, de las plantas y de la Tierra, vol. IV (geología), Instituto Gallach de Librería y Ediciones, 1963, p. 345-346
- (en)S. Conway-Morris, The community structure of the Middle Cambrian Phyllopod Bed, vol. 29, Paleontology, 1986, p. 423-467
- (en)M. Crichton, Jurassic Park, Alfred A. Knopf, New York, 1990 (ISBN 0-394-58816-9)
- (en)A.D. Greenwood et al., Nuclear DNA sequences from late pleistocene megafauna, vol. 16, Mol Biol Evol, 1999, p. 1466-1473
- (en)M. Höss, Neanderthal population genetics, vol. 404, Nature, 2000, p. 453-454
- (en)K.O. Kimberly et H.M. Robertson, Ancient DNA from amber fossil bees?, vol. 14, Mol Biol Evol, 1997, p. 1075-1077
- (es)M.J.S. Rudwick, El significado de los fósiles, Hermann Blume, 1987
- (en)K.S. Thomson, Living Fossil : The Story of the Coelacanth, Hutchinson Radius, 1991, p. 252
- (en)K.R. Walker et R.K. Bambach, The significance of fossil assemblages from fine-grained sediments: time-averaged communities, vol. 3, Geological Society of America Abstracts with Programs, 1971, p. 783-784
- (es)D.G.A. Whitten et J.R.V. Brooks, Diccionario de Geología, Alianza Editorial, Madrid, 1986, 300 p.
- (en)H.B. Whittington, The Burgess Shale, Yale University Press, New Haven, 1985, . . : .
- (en)E. Willerslev et al., Diversity of Holocene life forms in fossil glacier ice, vol. 96, Proceedings of National Academy of Science, USA, 1999, p. 8017-8021
Liens externes
- Paléontologie:eocène parisien - miocène du sud-ouest
- Fossiles du sud de la France
- La paléontologie et les fossiles
- la paléontologie, les poissons fossiles et les gisements de Provence
- Fossiles de Vertébrés et grands gisements paléontologique
- Site dédié à l'étude d'un gisement en région parisienne
- Portail sur la Paléontologie
- A la découverte des fossiles
- Association d'Etude et de Recherche Macrofaune du Cenozoique
- Images of Plant Fossils
- PP Collections, L'Ambre dominicain et inclusions, Collections d'ambres
 Portail de la Préhistoire
Portail de la Préhistoire Portail de la paléontologie
Portail de la paléontologie Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant Portail des sciences de la Terre et de l’Univers
Portail des sciences de la Terre et de l’Univers
Catégories : Fossile | Paléontologie






















Wikimedia Foundation. 2010.