- Histoire Évolutive Des Oiseaux
-
Histoire évolutive des oiseaux
Les oiseaux (Aves) sont des animaux bipèdes, ovipares, à sang chaud, en principe dotés de plumes. Très simplement reconnaissables, les oiseaux ont longtemps été placés dans le règne animal comme intermédiaires entre les reptiles et les mammifères. Cependant la paléontologie montre que les oiseaux sont des descendants de dinosaures tétrapodes et la génétique confirme que les Crocodilia forment le groupe actuel vivant le plus proche de celui des oiseaux. Plus surprenant, les oiseaux ne dérivent pas des reptiles volants tels que les Ptérosaures mais d'une autre branche. Un consensus définit ce groupe comme monophylétique.
 Le groupe des reptiles est en bleu.
Le groupe des reptiles est en bleu.
Sommaire
Origines
Généralités
Il a existé plusieurs théories quant à l'apparition des oiseaux. La plus communément admise fait des oiseaux les descendants de petits dinosaures théropodes. Une autre les présente comme descendants de reptiles du Permien, les Thecodontia.
Les études de Jacques Gauthier et d'autres paléontologues démontrent que de nombreux caractères qui étaient classiquement considérés comme spécifiques aux oiseaux sont apparus avant ceux-ci, chez les théropodes, voire plus tôt encore.
L'hypothèse des théropodes sous-tend que certains des caractères adaptatifs propres aux dinosaures terrestres, furent conservés chez les oiseaux. D'autres ont été modifiés comme les écailles appelées scutelles sur les pattes des gallinacés ou les plumes. Les modifications permirent le vol et un mode de vie arboricole. Naturellement, l'apparition des caractéristiques des théropodes qui précédèrent les oiseaux ne fut pas simultanée. De la même façon que l'œuf ne fait pas l'oiseau, la plume est présente chez de très nombreux théropodes. Bon nombre de caractéristiques propres aux dinosaures ne se retrouvent plus aujourd'hui que chez les oiseaux.Position des doigts et longueur des poignets aux doigts Rhamphorhynchus, un ptérosaure Archéoptéryx 

Le quatrième doigt porte l'aile, le poignet est long. Le second doigt est le plus long et porte l'aile,
pas de tarsométatarsien.L'anatomie comparée montre que les ptérosaures ne peuvent pas être les ancêtres des archéoptéryx pour des raisons morphologiques.
└─o Archosauria ├─o Crocodylotarsi (ie Crocodylia, les crocodiles) └─o Ornithodira ├─o │ └─o Dinosauria (les dinosaures, dont les théropodes, puis les oiseaux) └─o └─o Pterosauria (éteint) (les ptérosaures)
Les théropodes
L'ancêtre reptilien direct des dinosaures théropodes est un petit carnivore bipède qui marche comme un oiseau.
- Ses pattes avant sont libres pour saisir des objets ou des proies, et le plus long doigt est le deuxième, non le troisième comme chez les autres Sauropsides. Il a toutefois cinq doigts par main, alors que presque tous les théropodes primitifs et les oiseaux en ont trois. Chez cet animal, l'articulation de la cheville est déjà en charnière, et les métatarses sont allongés, relevés : l'animal marche, comme les dinosaures après lui. Les nombreuses modifications ultérieures du pied permettent probablement une foulée plus longue et plus rapide, qualité qui aidera ultérieurement les théropodes à développer le vol.
- Les premiers théropodes présentent des adaptations qui allègent leur squelette : des os creux et des cavités dans le crâne.

- Comme les oiseaux actuels, leur cou est long et leur dos horizontal.
Le quatrième doigt de la main des premiers dinosaures est déjà réduit, et le cinquième doigt a déjà presque disparu. Le quatrième doigt devient bientôt lui-aussi un moignon, puis ces deux doigts réduits disparaissent ensemble chez les théropodes tétanoures. Les trois doigts restants fusionnent au cours de l'évolution des oiseaux postérieurs à Archaeopteryx. - Les membres postérieurs des premiers théropodes se transforment aussi. Ils s'allongent : le fémur devient plus long que le tibia, et la fibula se réduit.

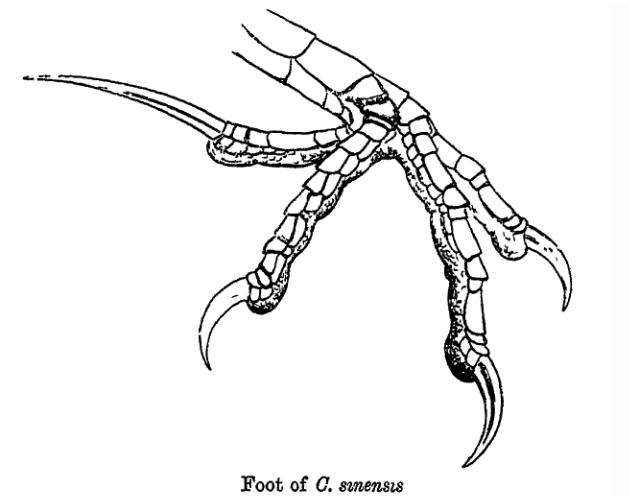
- Ces dinosaures marchent sur les trois doigts du milieu, ceux qu'utilisent toujours les oiseaux actuels. Le cinquième orteil, sans articulation, est raccourci et effilé. Le premier orteil, qui dépasse sur le côté du deuxième orteil, est formé d'un os raccourci, d'une articulation et d'une griffe. Il est placé au-dessus des autres orteils, tel un ergot.
- Les plumes (ou un protoplumage) sont déjà présentes chez de nombreux théropodes et devaient avoir d'autres utilités. Ainsi, déjà présentes chez les ancêtres des oiseaux, elles ont ensuite permis à ceux-ci de voler grâce à toute une autre série de caractéristiques.
o Tetanurae └─o Coelurosauria └─o ├─o Ornithomimosauria (éteint) └─o Maniraptora ├─o Ornitholestes, Compsognathidae, Alvarezsauridae (éteint) └─o ├─o Oviraptoriformes (éteint) │ ├─o Therizinosauria (éteint) │ └─o Oviraptorosauria (éteint) └─o Paraves
Apparition des oiseaux
- Puis au cours de l'évolution des théropodes, d'autres caractères qui semblaient être l'apanage des oiseaux apparaissent. Par exemple, le membre antérieur et la ceinture pectorale se modifient, favorisant d'abord la capture de proies, puis l'envol.
- Les bras s'allongent, sauf chez les carnivores géants, tels Carnotaurus, Allosaurus et Tyrannosaurus : chez les théropodes primitifs, le membre antérieur est deux fois plus court que le membre postérieur, mais, pour Archaeopteryx, le membre antérieur est plus long que le membre postérieur, et cette différence s'accentue encore chez les oiseaux plus récents, auxquels cet allongement confère un battement d'ailes plus puissant.

- Lorsque Archéoptéryx apparait, le premier orteil orienté situé plus haut a migré pour se placer derrière les autres.
Chez certains oiseaux plus récents, il sera plus bas sur la patte, opposable aux autres orteils, et il permettra aux pieds d'oiseaux arboricoles d'agripper fortement les branches ou de se transformer en serre.
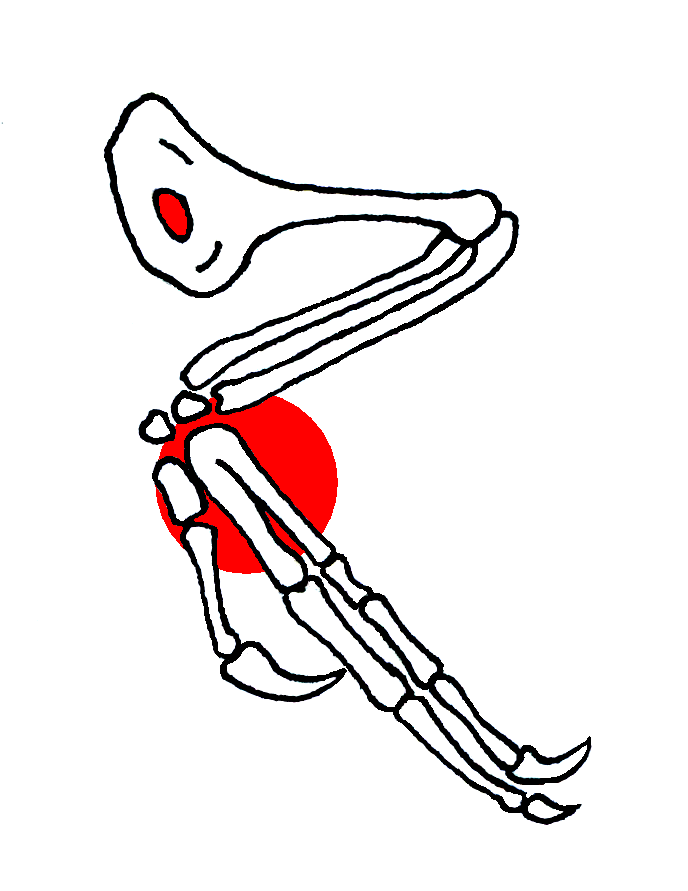
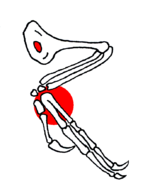 Aile de Confuciusornis. En rouge, l'ouverture de l'humérus et la soudure du Carpométacarpe, Chiappe et al. 1999).
Aile de Confuciusornis. En rouge, l'ouverture de l'humérus et la soudure du Carpométacarpe, Chiappe et al. 1999).- Les doigts des ailes aussi s'allongent, occupant progressivement une plus grande proportion du membre antérieur, et le poignet est remodelé. Les théropodes primitifs ont un os du poignet aplati, qui chevauche la base des premier et second os de la paume et les doigts. Chez les maniraptoriens, cet os a une forme de demi-lune le long de la surface de contact avec les os du bras. Cette forme en demi-lune est importante : elle permet à ces animaux de plier le poignet latéralement, et plus seulement verticalement. Ainsi, ils peuvent presque replier leur longue main comme le font les oiseaux actuels avec leurs ailes. Cette main allongée peut aussi être tournée et brusquement projetée vers l'avant pour la capture d'une proie. Seul les Hoazins ont gardés des griffes sur leurs ailes.
- Le sternum des tétanoures s'ossifie en deux plaques qui fusionnent, à l'origine il est cartilagineux et il devient le bréchet. L'ensemble de ces modifications consolident le squelette. Plus tard, ce renforcement soutient les muscles du vol ; la furcula devient un point d'ancrage pour les muscles du membre antérieur, permettant d'abord de saisir, puis de voler.
- Dans la région du bassin, au cours de l'évolution, des vertèbres s'ajoutent à la ceinture pelvienne, et l'orientation du pubis change, le pubis pointe vers l'avant et vers le bas ; puis il recule et, chez les oiseaux plus évolués qu'Archéoptéryx, devient parallèle à l'ischion. Nous ignorons ce qui a favorisé cette évolution, mais le partage de ces caractéristiques par les oiseaux et par d'autres Maniraptora témoignent de leur origine commune.
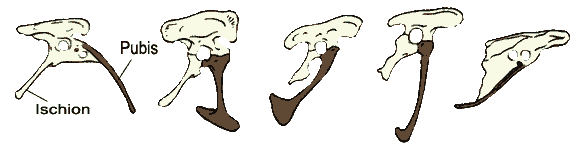
- Leur queue raccourcit et se rigidifie, devenant un balancier plus efficace pendant la course. Steven Gatesy, de l'Université Brown, a montré que cette transformation est associée à un changement de fonction : l'ancrage des muscles des membres postérieurs passe de la queue au bassin. Chez les maniraptoriens, les muscles qui tiraient les pattes vers l'arrière peuvent alors contrôler la queue. Chez les oiseaux qui évoluent après Archéoptéryx, ces muscles semblent être utilisés pour diriger la queue, couverte de plumes, pendant le vol.
La physiologie des oiseaux a dû s'adapter pour répondre aux besoins énergétiques nécessaires au vol :
- L'appareil respiratoire est devenu plus efficace et grâce à une disposition unique de plusieurs sacs aériens séparés des voies respiratoires, l'air circule de façon continue dans leurs poumons.
- Les oiseaux sont endothermes et leur niveau d'énergie ne varie pas avec la température ambiante, l'isolation thermique étant fournie par les plumes. Il est néanmoins probable que l'endothermie ait précédé la capacité de voler chez les théropodes.
Tous ces éléments physiologiques dont ne témoignent pas précisément les fossiles (organes mous internes non fossilisables et fragilité du squelette des oiseaux), la paléontologie des oiseaux ne fournit hélas pas autant de fossiles que pour d'autres créatures.
└─o Paraves ├─o Deinonychosauria (éteint) │ ├─o Troodontidae (éteint) │ └─o Dromaeosauridae (éteint) └─o Aves ├─o Archaeopterygidae (éteint) └─o Ornithothoraces
Caractéristiques anatomiques Théropodes ("non avaire")
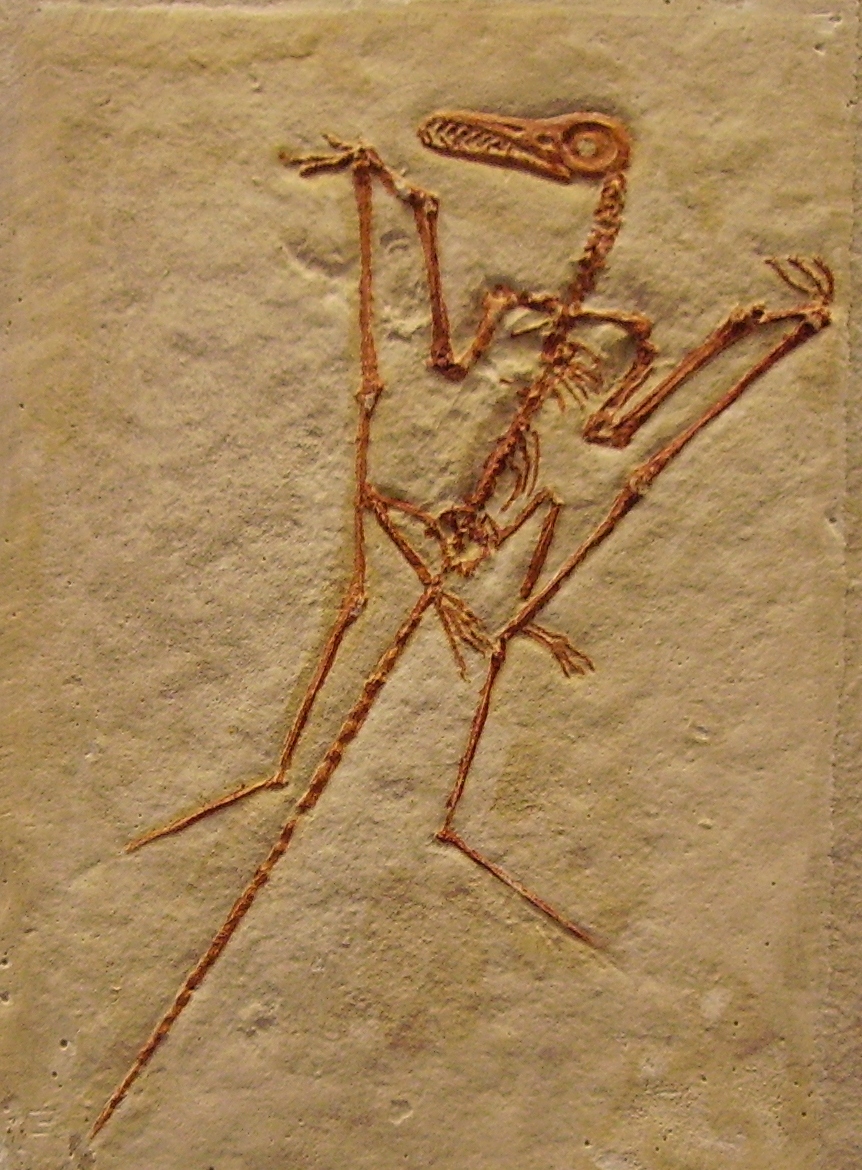
220 à 65 MaArchaeopteryx
156 à 150 MaConfuciusornis
125 et 110 Maoiseaux plus développés
Ornithothoraces à partir du Jurassique moyenFusion des deux fenêtres temporales avec l'orbite - + - + Disparition des dents -
nombreuses- + Beaucoup pour les
Oiseaux du CrétacéLa queue à Pygostyle raccourcie,
vertèbres de queue fusionnéesen petit nombre - + + Clavicules fusionnées (furcula)| -
Accollées
(forme de boomerang) |+
Forme de boomerang+
Forme de boomerang+
Forme en V,
à angle aigulongueur du Coracoïdes
long, en forme de ruban- - + + Sternum
avec bréchet- - -
Sternum plat avec quille trés réduite+
Seulement les Neognathae , disparition secondaire chez les paléognathesCôtes liées au sternum avec une - + + Disparition des Côtes abdominales
(Gastralia)- - - + côtes avec
Processus uncinéavec une côte supplémentaire ou présent chez les Maniraptora - + +
PlusieursHumérus dilaté
avec crête deltopectoral perforé (autoapomorphie)- - + - os de métacarpe des patte avant I, III
à Carpe-métacarpe fusionné- - + (fusion
après la base)+ os médian du pied II-IV fusionné
(Tarso-métatarse)un + + + os médian du pied IV present + + + - Plumes de vol actif, asymétriques quelques uns + + + Alula peu abondantes - - + Période d'affirmation
 Un arbre évolutif des oiseaux, depuis les théropodes.
Un arbre évolutif des oiseaux, depuis les théropodes.Certains auteurs défendent la thèse que les oiseaux pré-tertiaires appartiendraient à des groupes modernes de charadriiformes ou de gaviiformes. D’autres assurent que peu de lignées d’oiseaux ont survécu à la fin du Crétacé.
Les fossiles d’oiseaux sont rares car ils se conservent mal ; en outre, ils donnent peu d’indication sur les relations avec les oiseaux modernes. Les reconstitutions que l'on peut voir sont basées sur ce que nous connaissons des oiseaux d'aujourd'hui et de leur histoire. Certains points peuvent s'avérer faux.
Les fossiles d'oiseaux de mer, sont plus nombreux car ceux-ci vivaient dans un milieu ou des zones de sédimentation sont présentes[1].
Les plus anciens identifiés sont les Hesperornithiformes, comme Hesperornis regalis et vivaient au Crétacé. Ils avaient de petites dents pointues et ne pouvaient certainement pas voler. Les oiseaux marins d'aujourd'hui descendent plutôt d'un groupe apparenté aux Tytthostonyx glauconiticus
──o Pygostylia ├─o Confuciusornithidae └─o Ornithothoraces ├─o Enantiornithomorpha (éteint) └─o Ornithurae ├─o Yanornis (éteint) └─o ├─o Patagopteryx (éteint) └─o ├─o Hesperornithiformes (éteint) │ ├─o Enaliornithidae (éteint) │ ├─o Baptornithidae (éteint) │ └─o │ └─o Hesperornithidae (éteint) └─o Carinatae ├─o Ichthyornithiformes (éteint) └─o Neornithes
Durant le paléogène, les oiseaux de mer les plus fréquents sont classés dans le groupe des Procellariiformes parmi lequel on classe plusieurs espèces éteintes d'oiseaux de grande taille ressemblants à des pingouins[2].
C'est à la fin du Miocène et au cours du Pliocène que les genres modernes deviennent prépondérants, bien que le genre Puffinus remonte lui à l'Oligocène[1]. Leur diversité disparait soudainement, et, un peu plus tard, les mammifères marins semblent leur ravir la place. Ceci est peut être dû à un évènement astronomique qui a pu diminuer la biomasse des océans il y a 2 Ma[3], la nouvelle présence des Pinnipedia empêchant dès lors les oiseaux de retrouver leur diversité d'autrefois[4].
La radiation adaptative
Il existe aujourd'hui près de 10 000 espèces d'oiseaux.
 Les Pinsons de Darwin des Galápagos illustrent une radiation évolutive :
Les Pinsons de Darwin des Galápagos illustrent une radiation évolutive :
d'une espèce originale, il est apparu 4 types de bec pour 13 espèces au total.└─o Carinatae └─o Neornithes ├─o Paleognathae (autruche, émeu...) └─o Neognathae (les autres oiseaux.)
Les Paléognathes qui incluent les Tinamidae que l'on ne retrouve qu'en Amérique du Sud et Centrale et les Struthioniformes. Ils vivent aujourd'hui exclusivement sur l'hémisphère Sud. Ces oiseaux ne peuvent voler. Certains scientifiques pensent qu'ils représentent un groupement artificiel d'oiseaux qui ont perdu leur capacité de voler indépendamment ; en tout cas, les données disponibles quant à leur évolution ne sont pas très claires.
Les dates de différentiation entre les diverses espèces de Neognathae sont très disputées.
On observe cependant que la scapula s'allonge ensuite et s'affine, comme le coracoïde, qui s'étire ainsi vers le sternum. Les clavicules fusionnent sur leur ligne médiane et s'élargissent jusqu'à former une furcula en forme de boomerang.Les analyses génétiques, pour être précises, doivent être corroborées puis étalonnées par des fossiles et doivent porter sur des gènes significatifs qui témoignent de l'évolution.
De nombreuses discussions entre scientifiques portent sur cette question.Voir aussi: Liste d'oiseaux préhistoriques
Menace actuelle
L'évolution se produit généralement à une échelle trop lente pour être perçue par les humains. Cependant on note une forte diminution des espèces, ainsi actuellement 12 % des espèces sont reconnues en danger par l'UICN. Outre leur disparition par la prédation, l'empoisonnement par les pesticides, la destruction de leur environnement, on observe pour certaines espèces qui cherchent à s'adapter, une augmentation insoupçonnée de l'hybridation. En effet, certaines espèces proches, voyant leur habitat se morceler, se croisent en produisant même des hybrides stériles en quantité si importante que cela peut mettre ces espèces en péril. Par exemple, les nombreux oiseaux-mouches hybrides trouvés dans le nord-ouest de l'Amérique du Sud peuvent représenter une menace pour la conservation de ces espèces en espèces distinctes.
Par conséquent on observe donc une perte manifeste du capital génétique.
Genèse de cette théorie
Le premier fossile de ptérosaure a été décrit en 1784 et les paléontologues ne savaient s'il s'agissait d'une créature aquatique, d'une chauve-souris ou d'un oiseau. Georges Cuvier, en se basant sur les principes de l'anatomie comparée, annonce en 1801 qu'il s'agit d'un reptile volant et qu'ils ne sont pas les ancêtres des oiseaux.
Charles Darwin publie en 1859 L'Origine des espèces. La première découverte, qui allait être suivie de neuf autres, l'est sous la forme d’une plume fossilisée d'Archaeopteryx en 1861. Tous ces fossiles proviennent de la Bavière, conservés dans le calcaire de Solnhofen. La particularité de ce matériau est qu’il s’est formé dans un ancien lagon, sa composition ayant permis de protéger les moindres détails des êtres vivants (dont les plumes) qui s’y sont égarés lors de l’ère Mésozoïque. Des squelettes d’Archaeopteryx sont exposés au Muséum d'histoire naturelle de Londres et au Humboldt Museum de Berlin.
En 1862, est publié une étude d’Johann Andreas Wagner, un opposant à la théorie de Darwin, directeur du Musée de Paléontologie de Munich qui identifie un reptile, le Griphosaurus. En 1863, Richard Owen du British Museum y voit un oiseau. Le naturaliste britannique Thomas Huxley, est le premier à affirmer que c'est bien une forme transitoire. Il se demande également si les dinosaures n'auraient pas été des animaux à sang chaud (comme les oiseaux) plutôt qu'à sang froid (comme les reptiles) dans son ouvrage On the Animals which are Most Nearly Intermediate between Birds and Reptiles publié auprès de la Société géologique de Londres. En comparant l'anatomie de l'archéoptéryx et celle d'un autre fossile bavarois, le petit Compsognathus, il recense quatorze caractéristiques communes et voit entre eux un lien de parenté. Son hypothèse tomba dans l'oubli.
En 1870, le premier Hesperornithes est découvert par Othniel Charles Marsh et correctement décrit par lui-même en 1877. En 1887, Harry Govier Seeley publie sa théorie selon la laquelle les dinosaures se sont séparés en deux groupes qui se différencie selon leur ischion. Ironiquement, les oiseaux, qui sont, selon la théorie la plus admise, descendant des Saurischia (terme qui dérive de lézard à ischion) et non pas des Ornithischia (terme qui dérive du grec oiseau à ischion).
En 1924, la publication d'un dessin anatomique d'Oviraptor montrant une fourchette passe inaperçue, puis en 1936, à l'Université de Berkeley, Charles Camp découvre le squelette complet d'un petit théropode du Jurassique inférieur qui possède des clavicules. Des travaux plus récents démontrent, plus généralement, la présence de clavicules chez de nombreux théropodes apparentés aux oiseaux.
À la fin des années 1960, un siècle après la communication controversée de Huxley , John Ostrom, de l'Université Yale, décrit le squelette du théropode du groupe des maniraptoriens Deinonychus, prédateur aux griffes en faucille et grand comme un adolescent humain, qui rôdait dans le Montana il y a 115 Ma. Ostrom identifie un ensemble de caractères communs aux oiseaux, dont Archaeopteryx, Oviraptor et Deinonychus, et à d'autres théropodes, mais que ne possèdent pas d'autres Sauropsida. Il en déduit que les oiseaux descendent directement des théropodes.
En 1975, l'américain Robert Bakker jeta finalement le pavé dans la mare en affirmant que les dinosaures étaient les ancêtres directs des oiseaux contemporains. À l'Université de Berkeley, Jacques Gauthier entreprend une vaste étude cladistique des oiseaux, des dinosaures et de leurs parents reptiliens, en se fondant sur les comparaisons de J. Ostrom et sur bien d'autres caractères. Il confirme que les oiseaux descendent de petits dinosaures théropodes, et le maniraptorien aux griffes en faucille décrit par J. Ostrom, Deinonychus, est bien l'un des plus proches parents des oiseaux.
Vers la fin des années 1970 et durant toute la décennie suivante, Charles Gald Sibley et Jon Edward Ahlquist mènent des études basées sur des méthodes d'hybridation de l'ADN, ce qui modifie profondément les connaissances sur la phylogénie des oiseaux. Cependant les datations obtenues, en tenant compte du taux de mutations naturelles, laisse supposé que la diversification des oiseau est antérieur au Crétacé.
En 1981, une quarantaine de spécimens de Gansus, les plus anciens des Neornithes sont découverts en Chine. Des Confuciusornis découverts en 1993 bousculent un peu le schéma, sans le remettre en cause.
En 1993, la découverte de Vegavis, daté de 66 à 68 Ma, sont la première preuve de la coexistence entre thérapodes non-aviaires et oiseaux modernes, ce fossile pourrait remettre en cause la théorie de la lignée unique pour les oiseaux, offrant une données plus en phase avec la datation génétique[5].
En 2002, on a trouvé en Chine un spécimen de la taille d'un dindon, qui vivait aux alentours de 140 à 125 Ma. Le Jeholornis est un peu plus évolué que l'archéoptéryx. Plusieurs spécimens de Microraptor, des Dromaeosauridae trouvés en 2002, suggèrent que ceux-ci pouvaient voler. De plus, ces derniers possédaient certaines caractéristiques plus proches de celles des oiseaux qu'Archeopteryx qui leur est plus ancien mais considéré comme plus proche des oiseaux. La théorie la plus admise reste cependant celle de la lignée unique.
Classification classique
La vision traditionnelle de l'évolution des Neornithes (oiseaux modernes) est de placer à la base de l'arbre phylogénétique, après les ratites et tinamous, les groupes d'oiseaux marins tels les manchots, les grèbes, les plongeons et pélicans, etc.
Classification de Sibley
Articles détaillés : Dinosauria (classification phylogénétique), Aves (classification phylogénétique) et Taxinomie Sibley-Ahlquist.La nouvelle classification a montré que les canards et les gallinacés étaient les taxons les plus proches de la séparation d'avec les ratites (qui intègrent les tinamous) formant les paléognathes. Les Galloanserae constituant la lignée de base des Néognathes. Les groupes d'oiseaux marins traditionnellement considérés comme archaïques ont été placés dans un ordre des Ciconiiformes élargi, qui comprend aussi les rapaces diurnes (Accipitridae, Sagittariidae (Aves) et Falconidae) et les limicoles (Scolopacidae, Charadriidae, Laridae, etc.).
Cladogramme
Le cladogramme simplifié suivant suit la proposition de Norell et al. 2006, with clade names after the definitions by Sereno, 2005[6], [7].
└─o Sauria ├─o Lepidosauria (serpents, lézards...) └─o Archosauria ├─o Suchia (crocodiliens) └─o Dinosauria ├─o Ornithischia (éteint) └─o Saurischia └─o Theropoda └─o Coelurosauria └─o Maniraptora ├─o Oviraptoriformes (éteint) │ ├─o Therizinosauria (éteint) │ └─o Oviraptorosauria (éteint) └─o Paraves ├─o Deinonychosauria (éteint) │ ├─o Troodontidae (éteint) │ └─o Dromaeosauridae (éteint) (dont les microraptors) └─o Aves ├─o Archaeopterygidae (éteint) (les archéoptéryx) └─o Ornithothoraces ├─o Enantiornithomorpha (éteint) └─o Ornithuromorpha ├─o Hesperornithiformes (éteint) └─o Carinatae ├─o Ichthyornithiformes └─o Neornithes ├─o Paleognathae (autruches, émeux...) └─o Neognathae (les autres oiseaux)
Nomenclature des ordres d'Oiseaux modernes
Correspondances entre classifications récentesVoir aussi
Articles connexes
Sources
- Archaeopteryx, volatile controversé, Association FABULA
Liens externes
- gigadino, le site sur la paléontologie et les fossiles.. avec des dossiers et documentation
- Evolution, dinosaures et Oiseaux
- Archaeopteryx, volatile controversé, Association FABULA
- Dinosaures et oiseaux sur astronomie et d'astronautique.com
- (en)Aves: Overview sur Palæos
- (en)David Swanson, « Zoology 461/561, Ornithology, Summer 1999 (July 6-29) », sur l'origine et la morphologie des oiseaux.
- (en)Gary Ritchison, « Introduction to Birds », Department of Biological Sciences Eastern Kentucky University
- (en)Fossil records of avian taxa sur ornitaxa.com, Bird Taxonomy for Birders
- (fr)Une analyse moléculaire confirme le lien entre le Tyrannosaure et les oiseaux sur ornithomedia.com
- (fr)Une nouvelle étude bouleverse l'arbre phylogénétique des oiseaux sur ornithomedia.com, 27/6/2008
Notes
- ↑ a et b Schreiber, Elizabeth A. & Burger, Joanne., Biology of Marine Birds
- ↑ (en) Goedert, J., « Giant Late Eocene Marine Birds (Pelecaniformes: Pelagornithidae) from Northwestern Oregon" », dans Journal of Paleontology, vol. 63, no 6, 1989, p. 939–944 [résumé]
- ↑ (en)Comins, Niel F.; William J. Kaufmann III, , Discovering the Universe, 7th edition, New York, NY: Susan Finnemore Brennan, 2005, 359 p. (ISBN 0716775840.)
- ↑ (en) Olson, S. & Hasegawa, Y., « Fossil Counterparts of Giant Penguins from the North Pacific », dans Science magazine, vol. 206, no 4419, 1979, p. 688–689.
- ↑ (fr)Jean-Luc Goudet, « Après la chute de l'astéroïde, le canard était toujours vivant », Futura-Science
- ↑ (en) Norell, M.A., Clark, J.M., Turner, A.H., Makovicky, P.J., Barsbold, R., et Rowe, T., « A new dromaeosaurid theropod from Ukhaa Tolgod (Omnogov, Mongolia). », dans American Museum Novitates, vol. 3545, 2006, p. 1-51.
- ↑ (en)Sereno, P. C., McAllister, S., and Brusatte, S. L., « TaxonSearch: a relational database for suprageneric taxa and phylogenetic definitions. », PhyloInformatics
 Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant Portail de l’ornithologie
Portail de l’ornithologie
Catégories : Oiseau | Histoire évolutive






Wikimedia Foundation. 2010.