- Theorie synthetique de l'evolution
-
Théorie synthétique de l'évolution
La théorie synthétique de l'évolution constitue le cadre conceptuel le plus largement utilisé dans l'étude scientifique des processus d'évolution en biologie. Cette théorie est basée sur l'intégration de la théorie de l'hérédité mendélienne et de la génétique des populations à la théorie darwinienne[1]. Cette synthèse fut menée au cours des années 1930 et 1940 par R.A. Fisher, J.B.S Haldane, Sewall Wright, Julian Huxley, Ernst Mayr, Bernhard Rensch, George Gaylord Simpson et George Ledyard Stebbins. Baptisée ainsi par Julian Huxley en 1942, cette théorie est aussi appelée néodarwinisme ou synthèse néodarwinienne pour souligner le fait qu'elle constitue une extension de la théorie originale de Charles Darwin qui ignorait les mécanismes de l'hérédité génétique.
Histoire de la théorie synthétique de l'évolution
Définition
Aujourd'hui, l'évolution n'est plus envisagée comme la transformation d'individus isolés mais comme celle de groupements d'individus de même espèce, c'est-à-dire des populations. Mais le principe de base est le même : elle explique l'évolution par l'action de la sélection naturelle sur des populations.
Une population évolue quand la fréquence d'une version d'un gène appelée allèle (ou de plusieurs allèles) s'y modifie. On voit ainsi se répandre dans certaines espèces des caractères ayant acquis, en raison de changements du milieu, une valeur adaptative qu'ils ne possédaient pas auparavant ; les individus porteurs de ces caractères sont particulièrement favorisés dans le nouveau milieu, auquel ils se trouvaient en quelques sortes "préadaptés" ; ils constituent alors rapidement une grande partie de la population ou même toute la population de l'espèce. Les caractères qui se répandent alors correspondent à des allèles existant auparavant "silencieusement" au sein de l'espèce.
Lorsque l'ensemble des individus qui constituent une espèce forme plusieurs populations isolées, chacune de ces populations peut acquérir des caractères particuliers et donner naissance à des variétés différentes au sein de la même espèce. Si ces variations sont, par la suite, dans l'impossibilité de se croiser, elles divergent de plus en plus et finalement sont interstériles : elles constituent alors des espèces distinctes.
La "barrière" qui sépare les variétés d'une même espèce peut être de nature variée.
L'évolution, faits et théorie

Une observation des êtres vivants révèle l'existence de nombreux points communs dans l'organisation anatomique, physiologique et biochimique des organismes. Depuis l'Antiquité, l'Homme a tenté de trouver un ordre dans la nature[2] selon les ressemblances et les différences qu'il observait chez les animaux et les végétaux donnant naissance à diverses classifications des espèces. Si certaines espèces se ressemblent beaucoup et d'autres moins, l'unité du vivant n'a guère été remise en cause ; elle a même été renforcée d'abord au cours du XIXe siècle par la découverte de la cellule, unité vivante fondamentale de tous les organismes, puis au XXe siècle par la découverte de l'ADN comme support de l'hérédité commun à tout le vivant.
Le phénomène d'évolution permet d'expliquer ces ressemblances par l'existence de liens généalogiques entre toutes les formes de vie : les organismes se ressemblent parce qu'ils partagent des caractères hérités d'un ancêtre commun[3]. Des faits observés dans plusieurs disciplines scientifiques corroborent cette théorie.
Tous les êtres vivants fonctionnent sur les mêmes bases moléculaires (ADN, ARN, protéines…) et utilisent le même code génétique. Les différences entre les espèces et les variations entre individus au sein d'une même espèce sont dues essentiellement à des différences dans la séquence des gènes et la structure des chromosomes, différences provoquées par des mutations et des réarrangements chromosomiques occasionnels. Ces évènements aléatoires, qui produisent des individus dont les gènes sont légèrement différents de ceux de leurs parents, sont certes rares et - comme une coquille qui se glisse dans l'impression d'un texte - plus souvent nocifs qu'utiles, mais suffisent pour introduire la variabilité, source de l'évolution[4].
Les différentes espèces d'un embranchement, malgré des aspects extérieurs très différents, partagent un plan d'organisation invariable[5]. L'homologie des organes entre tous les membres d'un groupe biologique se retrouve à n'importe quel niveau de la classification. Par exemple, chez tous les mammifères, les membres sont organisés de la même façon : ceux d'une baleine, d'un lion, d'une chauve-souris ou d'un homme comportent les mêmes os, même si leur forme est très variable et qu'ils sont employés à des fonctions complètement différentes.
Par ailleurs, il existe une certaine variabilité des caractères au sein de chaque espèce, c'est-à-dire qu'il n'existe aucun individu complètement identique à un autre sauf cas exceptionnels. Cette variation peut être de plus ou moins grande ampleur, mais elle montre en tout cas que les êtres vivants ne sont pas fixes. L'éventail de variations que présente une population vivante est le matériau de base avec lequel l'évolution pourrait construire des organismes de plus en plus différents. On assiste directement à des transformations au sein de populations d'êtres vivants pouvant être interprétées comme des spéciations en cours[6]. L'acquisition de nouveaux caractères chez certains virus et bactéries a également été constatée pour être combattue, dans le cas de la résistance aux antibiotiques, ou au contraire utilisée, pour la capacité à digérer des composés synthétiques.
Les fossiles témoignent de la disparition de certaines espèces d'êtres vivants depuis l'apparition de la vie sur Terre il y a au moins 3,5 milliards d'années[7]. Certaines espèces sont appelées des formes « intermédiaires » entre différents groupes et permettent de mettre en évidence le phénomène d'évolution[8]. Par exemple, l'archéoptéryx, qui serait une forme de transition entre certains dinosaures et les oiseaux, ou Rodhocetus (ou Ambulocetus) dont l'anatomie semble se situer entre celle des mammifères terrestres et celle des baleines.
Les détracteurs de l'évolution soulignent qu'elle n'est « qu'une théorie ». En fait il ne s'agit pas d'une théorie au sens commun mais dans le sens scientifique du terme[9]. Cela signifie qu'il s'agit d'un ensemble de connaissances basé sur quelques suppositions de départ, et développé par des démonstrations rigoureuses à partir de ces principes. Le développement de la théorie est donc toujours susceptible d'être complété, ou même corrigé sur certains points. En revanche il n'est pas correct de considérer l'ensemble de l'édifice comme une simple hypothèse, sauf à disposer d'arguments forts contre les mécanismes de base de la théorie qui ont été observés en conditions expérimentales et naturelles.
Mécanismes de l'évolution
 Illustrations de cinq types de mutations chromosomiques.
Illustrations de cinq types de mutations chromosomiques.
L'évolution dépend de divers processus qui tendent à modifier la fréquence des allèles au sein de populations : mutation : dérive génétique, étranglement génétique ; flux de gènes, brassage génétique ; sélection naturelle : sélection stabilisante, directionnelle ou diversifiante ; sélection sexuelle : accouplement non aléatoire, ... ; pédomorphose ; pléiotropie : pléiotropie antagoniste ; ...
L'évolution des espèces est permise par les mutations que subissent les gènes portés par les chromosomes (constitués d'ADN). Tout être vivant possède de l'ADN (cela corrobore l'idée d'une origine commune des espèces) : cette molécule est constituée de deux hélices complémentaires au niveau de leurs bases azotées. Des mutations peuvent affecter cet ADN ; elles sont provoquées par des agents mutagènes tels que rayons X, alpha, UV, ou tout simplement par la défaillance des organites responsables de la réparation de l'ADN mal transcrit ou traduit. La théorie des monstres prometteurs met le stress provoqué par des facteurs externes au premier plan de ces facteurs de mutation. Ces mutations affectent la séquence d'un gène concerné (ordre des bases nucléotides d'un gène : adénine, thymine, guanine et cytosine).
Ces mutations sont à l'origine du polymorphisme des gènes, c'est-à-dire le fait que deux versions d'un même gène (deux allèles) par exemple, sont présentes chacune dans au moins un pour cent de la population de l'espèce considérée. Ces mutations créent donc de nouveaux allèles. Mais il ne faut pas oublier le phénomène des familles multigéniques, qui a un impact très important dans l'évolution : un gène peut être dupliqué et transposé sur un autre chromosome, ainsi tous les gènes issus de ce gène ancestral font partie d'une famille multigénique.L'environnement « encadre » ces mutations par le biais d'un phénomène appelé sélection naturelle : un gène présentant un avantage pour une espèce dans un environnement donné, permettant à ses représentants d'atteindre le mieux possible la maturité sexuelle, se répand chez les individus d'une même espèce, a contrario s'il est néfaste, il disparait. Quant aux gènes neutres, ils se répandent de façon aléatoire mais peuvent permettre de suivre l'évolution (mutation de gènes homéotiques). C'est donc l'environnement qui décide de l'évolution des espèces, celles-ci évoluant pour être toujours plus adaptées à celui-ci. Donc le caractère aléatoire des mutations de l'ADN est compensé par la sélection environnementale.
Il est donc très important de ne pas confondre évolution et innovation, ce qu'il faut retenir, c'est que l'espèce qui survit est l'espèce la mieux adaptée, pendant une certaine durée, à son environnement.
Par ailleurs, il ne faut surtout pas voir l'évolution d'un point de vue généalogique, mais phylogénétique, en effet les espèces ne descendent pas les unes des autres. Des phénomènes comme la dérive génétique font que deux populations d'une même espèce isolées pendant une très longue période de temps divergent et forment deux nouvelles espèces. Par exemple pour la lignée humaine, l'arbre phylogénétique est buissonnant : plusieurs espèces Homo et Australopithèque ont vécu simultanément. Il est aussi à noter que l'homme - contrairement aux idées reçues - ne descend pas "du singe", il a un ancêtre en commun avec lui. Et cet ancêtre n'était pas un singe. Notre patrimoine génétique est très proche de celui des chimpanzés, en effet les différences entre nous et celui-ci ne tiendraient qu'à la différence de quelques gènes déterminants : ils nous permettent de garder des caractères juvéniles toute notre vie, nous permettant de conserver la bipédie (le petit chimpanzé est quasi-bipède) et ils allongent la phase embryonnaire, permettant la mitose de beaucoup plus de neurones.
L'évolution résulte de l'action de plusieurs mécanismes évolutifs susceptibles de modifier la fréquence des génotypes dans une population, et par conséquent de modifier des caractères morphologiques, physiologiques ou comportementaux.
Le processus est dans tous les cas identique, dans le même ordre :- Apparition d'un nouveau caractère héréditaire chez un ou quelques individus.
- Ce nouveau caractère se répand dans les populations, soit parce qu'il est plus adapté à l'environnement, soit à cause d'un effet de hasard.
L'apparition de caractères nouveaux et héréditaires
Les mécanismes de
l'évolution biologiqueMécanismes non aléatoires:
Mécanismes aléatoires:
Dynamiques évolutionnaires :
A chaque génération, de nouveaux variants génétiques apparaissent qui peuvent eux-mêmes transmettre leur patrimoine génétique au moment de la reproduction. Ces sources de diversité génétique sont :
- les mutations ponctuelles dans la séquence d'ADN des gènes ;
- les réarrangements chromosomiques ;
- les recombinaisons ou brassages génétiques qui se produisent lors de la reproduction sexuée et, chez les bactéries, lors de transferts d'ADN entre cellules.
On peut ajouter à cette liste une autre source de diversité génétique que sont les migrations par lesquelles le stock génétique dans une population donnée se voit renouvelé par l'arrivée d'autres membres de l'espèce porteurs d'un pool génétique différent.
La mutation
Les mutations résultent d'erreurs lors de la réplication des gènes. Ces modifications accidentelles peuvent être transmises à la descendance, donnant naissance à des individus porteurs de gènes nouveaux, qu'aucun de leurs parents ne portait.
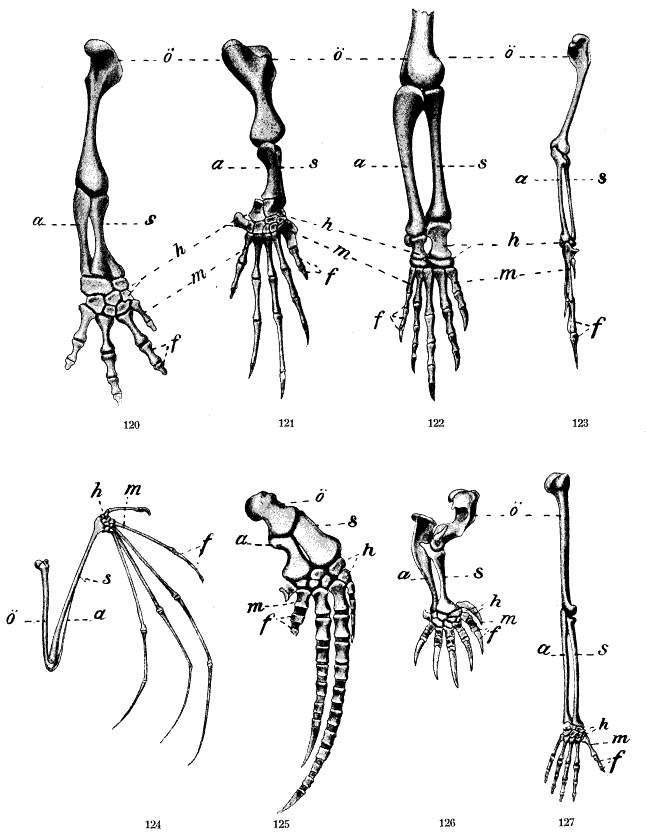
Il existe plusieurs types de mutations (mutations ponctuelles, duplications de gènes, cassures des chromosomes, insertions ou délétions de séquences). Ces différentes modifications suffisent à expliquer la diversité génétique observée dans la nature.
Il peut s'y ajouter des mutations par échange de matériel génétique entre espèces, par différents mécanismes naturels :
- Les bactéries sont capables d'intégrer et d'utiliser du matériel génétique (des plasmides) échangé entre deux bactéries, voire simplement présent dans le milieu. Le plus souvent cela ne conduit pas immédiatement à la définition d'une nouvelle espèce, mais le phénomène peut y contribuer et il contribue de façon importante à la sélection naturelle (par exemple, de nombreuses résistances aux antibiotiques se diffusent de cette façon).
- Les virus peuvent servir de vecteurs de transfert de matériel génétique d'une espèce vers une autre, même très différente ; le plus souvent, l'infection virale conduit à la mort de la cellule victime, mais ce n'est pas nécessairement le cas, et la cellule cible peut participer à la reproduction (notamment pour les plantes). Un des exemples les mieux connus (et les plus utilisés par la recherche) est celui du tabac et des mosaïques.
- Les plantes peuvent s'hybrider, conduisant à de nouvelles espèces (le blé étant un exemple typique).
Finalement, il faut mentionner les symbioses strictes, qui sont des créations de nouvelles espèces à partir d'anciennes, mais sans échange ni fusion du matériel génétique (au moins dans un premier temps). Les lichens et, selon l'hypothèse de l'endosymbiose, les organites dits « autonomes » à l'intérieur des cellules (chloroplastes, mitochondries) relèvent de ce type d'association.
Les mutations augmentent la variabilité génétique au sein d'une population. Cependant, elles sont rarement bénéfiques à l'individu qui les porte car elles peuvent détruire l'activité d'un gène essentiel.
La migration
La migration est une force de l'évolution qui augmente la variabilité génétique d'une population. De nouveaux arrivants en provenance d'une population éloignée amènent des allèles nouveaux, augmentant le pool génétique de la population d'accueil.
Le nouveau caractère se répand dans les populations
Les mécanismes de l'évolution supposent qu'un ensemble d'individus, regroupés en une espèce sur la base de caractères partagés, évoluent « conjointement. » Se pose donc le problème de la spéciation, c'est-à-dire de l'évolution des espèces au-delà des simples individus.
La dérive aléatoire
Lorsqu'une petite population est isolée du reste d'une population-mère (par exemple, par isolement sur une petite île), certains types d'allèles tendent à disparaître, du fait de faible nombre de copies distribuées dans le pool génétique de la population au départ. Au fil des générations, la population dérive alors, acquérant des caractéristiques qui lui sont propres (ex: petitesse (mammouth nain de Sicile), gigantisme (varan de Komodo) parmi les espèces insulaires). La dérive est une force de l'évolution qui diminue la variabilité génétique au sein de la population. La dérive influe sur la fréquence des allèles. Elle ne crée en aucun cas de nouveaux allèles. Certains allèles vont être surreprésentés et d'autres sous-représentés par rapport à la population d'origine. Cette modification de la fréquence s'explique par une erreur d'échantillonnage. (hasard)
La sélection naturelle
Elle diminue la variabilité génétique d'une population en enlevant certaines combinaisons génétiques défavorables dans le milieu où elles se produisent. Cependant elle peut aussi maintenir à une fréquence stable, des nouveaux variants génétiques, sans pour autant éliminer les anciennes versions. C'est le cas pour l'allèle de la drépanocytose. La sélection naturelle participe donc à l'augmentation de la diversité génétique.
Une image donnée par Richard Dawkins
Pour se trouver mille ancêtres différents, il faut remonter en arrière d'une dizaine de générations, ce qui représente quelques siècles. Or, avant le XIXe siècle, la moitié au moins des enfants mouraient en bas âge : on peut donc se demander combien de nos mille derniers ancêtres sont morts en bas âge, et la réponse n'est pas « au moins la moitié », mais bien entendu zéro, par définition. Nous ne sommes donc nullement représentatifs de l'humanité passée, car descendants d'une longue lignée de gens qui ont tous eu la chance d'amener une progéniture à l'âge de la procréation (c'est-à-dire survivre et trouver un partenaire, entre autres). Une génération peut avoir de la chance. Quand une dizaine en a coup sur coup, on peut supposer que cette « chance » correspond en fait à un ensemble de facteurs favorables qui se retrouvent de l'une à l'autre (facteurs qui ne sont pas tous génétiques : ils peuvent être culturels, religieux, économiques, etc.) Cette considération à elle seule montre que même à notre échelle, nous avons participé un tout petit peu, sur les quelques derniers siècles, à l'évolution. D'ailleurs notre simple choix d'un conjoint se révèle, comme le montre le biologiste Geoffrey Miller (The Mating Mind) obéir à des choix pas toujours conscients qu'on peut rapprocher de l'eugénisme.
Richard Dawkins pousse plus avant ses spéculations en considérant l'être vivant comme un conteneur à gènes (ce serait en tout cas le point de vue des gènes, s'ils en avaient un !). Pour lui les produits des gènes - cellules, tissus, organes, organismes, sociétés - servent aux gènes à se répliquer et à survivre. En quelque sorte, à la question : « De l'œuf ou la poule, qui est le premier ? », il répond : « La poule est le moyen trouvé par l'œuf pour faire d'autres œufs. ». Ce renversement de perspective considère l'être vivant comme la marionnette (Dawkins utilise le terme de véhicule-robot) de ses gènes.
Tout en la reconnaissant ingénieuse, Stephen Jay Gould a mis en garde contre une prise trop à la lettre de cette vision. Les gènes ne possèdent ni intention, ni projet au sens que nous donnons à ces termes. Dawkins n'a utilisé le terme qu'en tant que métaphore parlante, mais ses lecteurs n'ont pas toujours saisi la nuance.
En outre, il convient de se rappeler que les facteurs de survie ne sont pas seulement génétiques, ce qui est bien évident chez l'homme, mais ce qui est aussi le cas chez de nombreux animaux (par exemple, le chant des oiseaux, essentiel dans leur reproduction, dépend d'un apprentissage).
Devenir des mutations et caractères hérités
- Les mutations se font toujours au hasard, et sont le plus souvent létales. La plupart du temps, elles perturbent gravement le fonctionnement de l'organisme mutant. Bien sûr, il arrive aussi qu'elles soient neutres (en modifiant un caractère sans pour autant le rendre moins fonctionnel) ou avantageuses; le cas est beaucoup plus rare, mais les mutations portent sur des millions d'individus pendant des dizaines de milliers de générations ou davantage, ce qui assure un facteur multiplicatif de 1010 à chacun de ces évènements aléatoires.
- C'est la sélection naturelle, mise en évidence par Darwin, qui se charge de « faire le tri » entre les différentes variations. Il s'agit d'un processus purement mécanique : les mutants défavorisés auront tendance soit à mourir plus jeunes que les autres, soit à trouver plus difficilement un compagnon de reproduction. Dans un cas comme dans l'autre, ils laisseront moins (voire pas du tout) de descendants. Les mutants neutres mourront en moyenne au même âge que les autres, auront globalement autant d'enfants et pourront donc répandre leurs nouveaux caractères dans une population sous l'effet du hasard. Quant aux mutants avantagés, ils seront plus compétitifs et auront donc en moyenne plus de descendants. Une mutation qui permet de mieux échapper à un prédateur, de mieux s'orienter, de mieux séduire le sexe opposé, d'avoir plus de descendants, de mieux résister aux maladies, de mieux tirer profit d'une vie en société aura tendance à faire baisser le taux de mortalité chez les mutants ou à améliorer leur succès reproductif.
- Génération après génération, la sélection naturelle favorise les mutations qui se transmettent le plus, dans les populations (la dérive génétique, qui se fait au hasard, permet également à quelques mutations neutres de se fixer dans le génome de l'espèce). L'adaptation des populations tend donc progressivement à s'améliorer ; l'émergence d'organes aussi complexes et aussi fonctionnels que l'œil humain ou le radar de la chauve-souris s'explique par un tel processus de sélection cumulative. En revanche, l'existence de caractères sans utilité adaptative apparente peut s'expliquer par la fixation au hasard de mutations neutres. L'hypothèse dite de « l'auto-stop », ou « hitch hiking » en anglais, explique la fixation des mutations neutres par la mutation sur le même chromosome de deux gênes, l'une étant neutre et l'autre étant positive qui se transmettront simultanément.
Conséquences de l'évolution
Apparition d'espèces nouvelles
La spéciation désigne l'apparition de nouvelles espèces à partir d'une espèce ancestrale. Une espèce est un ensemble d'individus interféconds (interfertiles), isolés du point de vue reproductif des autres êtres vivants. L'apparition d'une barrière reproductrice (arrêt des échanges génétiques entre elles) au sein de cet ensemble homogène entraine l'apparition de deux groupes isolés (isolement reproductif). Les deux groupes vont ensuite évoluer indépendamment l'un de l'autre, et vont progressivement accumuler des différences génétiques, jusqu'à ce que l'isolement devienne irréversible.
- La spéciation dite allopatrique concerne deux ou plusieurs populations isolées géographiquement, séparées les unes des autres par des barrières géographiques (océan, montagne…). Un petit isolat peut aussi se former en périphérie de l'aire globale de distribution de l'espèce. Chaque population va alors pouvoir évoluer indépendamment, d'autant plus rapidement qu'elle est petite (les mutations se fixent plus facilement, pour des raisons mathématiques, dans les petites populations) et ainsi accumuler des remaniements chromosomiques pour finir par former une nouvelle espèce.
- La spéciation sympatrique : il arrive aussi qu'un isolement reproducteur apparaisse au sein d'une population qui ne sera jamais séparée géographiquement. Il suffit pour cela que quelques individus changent de période de reproduction ou de signaux pour la parade nuptiale pour que les autres représentants de l'espèce cessent rapidement de s'accoupler avec eux ; il y a alors formation de deux communautés se partageant le même espace, mais s'isolant sur le plan de la reproduction. Les deux populations, en évoluant chacune de leur côté, pourront finir par se transformer en deux espèces complètement différentes.
Comme les spéciations sont des phénomènes relativement courts, qui se produisent dans de petites populations isolées, on observe souvent une certaine discontinuité entre les différentes espèces dans les archives fossiles. Une espèce peut ne pas changer pendant très longtemps, puis être rapidement (à l'échelle géologique) remplacée par une autre.
Les mutations à l'origine des grandes modifications évolutives
Certaines mutations minimes permettent à l'évolution d'agir très rapidement. On connait des gènes régulateurs, dits gènes homéotiques, qui déterminent les grandes lignes du développement et du plan d'organisation de chaque partie du corps. Lorsque ces gènes (qui régulent eux-mêmes quelques milliers d'autres gènes) mutent, l'individu obtenu est souvent porteur de caractères nettement différents de ceux de ses parents. Certains « sauts évolutifs » s'expliquent par ce type de « macro-mutations » (que Goldschmidt avait découvert en proposant sa théorie du « monstre prometteur »). Par exemple, l'apparition de doigts et la disparition des rayons au bout de la nageoire des poissons crossoptérygiens (c'est-à-dire la formation des premières ébauches de pattes) s'expliquent par un simple décalage de l'activité de deux gènes homéotiques : Hox-a et Hox-b. Cette étape évolutive-là n'a pas nécessité des milliers, ni même des dizaines de mutations différentes : les gènes homéotiques ont permis l'émergence rapide de caractères radicalement nouveaux. Même si les « macro-mutations » donnent le plus souvent des « monstres » incapables de survivre et même si le processus, lent et sûr, de sélection cumulative de milliers de mutations minuscules a apparemment joué un rôle plus important dans l'évolution, il est de fait que certains « monstres prometteurs » ont permis à l'évolution de faire de grands bonds en avant.
D'autres mutations modifient la chronologie du développement : on parle d'hétérochronies. Elles peuvent être à l'origine de l'apparition d'une espèce plus juvénile (paedomorphique) ou, au contraire, plus adulte (hypermorphique) que son ancêtre. On connait plusieurs exemples d'histoires évolutives qui ont fait intervenir les hétérochronies. L'axolotl, un urodèle mexicain qui passe sa vie entière à l'état larvaire sans jamais se métamorphoser, est paedomorphique par rapport à son ancêtre l'ambystome. Une simple hétérochronie a permis l'apparition d'une espèce complètement différente. De même, il semblerait que l'homme soit paedomorphique par rapport à ses ancêtres simiens, car il garde toute sa vie des caractères juvéniles que le chimpanzé et le gorille perdent. De plus, les phases générales de son développement sont ralenties par rapport à celles des grands singes.
L'apparition de nouveaux gènes s'explique principalement par la duplication de gènes préexistants. On connaît plusieurs « familles de gènes » dont les différents membres sont apparemment issus de plusieurs duplications d'un même gène ancestral. Par la suite, chacun des exemplaires « dupliqués » a pu muter et évoluer indépendamment des autres. Plusieurs familles multigéniques sont de toute évidence apparues grâce à ce processus.
De même, des duplications de l'ensemble du génome (polyploïdisations) ont également pu jouer. Par exemple, on estime que deux épisodes de polyploïdisations se sont produits depuis la séparation des deutérostomiens (soient essentiellement les échinodermes et les chordés dont les vertébrés) et des protostomiens (les invertébrés) sur la branche des vertébrés. On retrouve ainsi que beaucoup de gènes, comme HedgeHog impliqué dans le développement des polarités chez la drosophile, existent en trois ou quatre exemplaires chez les vertébrés (Desert HedgeHog, Indian HedgeHog et Sonic HedgeHog) pour des fonctions similaires.
Approche évo-dévo : évolution et développement
Depuis les années 1980, les travaux visant à relier la génétique et l'embryologie apportent un nouvel éclairage sur les mécanismes de l'évolution biologique. Ces nouvelles approches réactualisent certains aspects de théories plus anciennes comme la théorie du monstre prometteur de Goldschmidt.
Ces travaux montrent en effet que les homologies observées au niveau des organismes se retrouvent aussi au niveau du développement. L'existence de gènes "chefs d'orchestre" quasi-identiques chez des espèces aussi diverses que les mouches, les souris et les humains, change la conception du rôle des gènes dans la construction de l'individu (l'ontogenèse) et dans les réorganisations anatomiques qui se produisent au cours de l'histoire évolutive (la phylogenèse). En plus de confirmer l'ascendance commune entre les espèces vivantes, ces découvertes montrent que de petites modifications dans la séquence génétique au niveau de ces gènes voire une simple différence dans la période pendant laquelle de tels gènes sont exprimés au cours de l'embryogenèse, peuvent avoir des effets très importants sur la morphologie de l'organisme. Cela montre que l'évolution ne consiste pas seulement à créer de nouveaux gènes par mutation mais aussi à changer la façon dont les gènes sont exprimés.
Parmi les autres découvertes de la biologie moléculaire, les mécanismes épigénétiques constituent une autre voie par laquelle de l'information peut être transmise "à côté" du seul code génétique. Ce type de transmission peut impliquer l'ADN, l'ARN ou les protéines ou les organites du cytoplasme cellulaire et peut être réversible ou non. L'un des processus épigénétiques les plus étudiés est la méthylation de l'ADN et son lien avec les modifications de la chromatine : ce mécanisme consiste en l'inactivation d'un gène par changement de la conformation de la chromatine du chromosome qui le porte. Ce changement rend le gène silencieux sans en affecter la séquence nucléotidique. Par conséquent, ce gène sera transmis "intact" mais silencieux aux générations suivantes. On pourra donc observer la résurgence d'un caractère ancestral plus tard dans la lignée simplement par réactivation du gène (via une reconformation de la chromatine, déclenchée par un évènement stressant dans l'environnement). L'importance de ces mécanismes épigénétiques est encore difficile à mesurer mais ils peuvent avoir des conséquences importantes dans l'explication de phénomènes qui ne respectent pas les lois de l'hérédité mendélienne.
Controverses actuelles
Adaptationnisme et Neutralisme
Au sein de la communauté scientifique, le débat concernant les théories modernes de l'évolution se porte donc sur le rôle que joue le "hasard" (la dérive génétique (?!), produisant la diversité des caractères) par rapport à la sélection naturelle. Les théories neutralistes ont ainsi été critiquées par des biologistes tenant d'un vision adaptationniste de l'évolution pour lesquels les pressions de sélection (naturelle et sexuelle) sont les principales forces dans l'évolution des espèces, le "hasard" ne jouant finalement que peu de rôle.
- Argumentation : Les adaptationnistes s'appuient notamment sur les phénomènes de convergence évolutive par lesquels différentes espèces aboutissent à la même solution évolutive lorsqu'elles sont soumises au même problème adaptatif[10].
La forme fuselée de l'ichthyosaure, de la baleine ou du manchot qui ont la forme des poissons témoignent de la force de la pression de la sélection naturelle qui plaide en faveur des adaptationnistes.
Le hareng est un poisson osseux. Les ichthyosaures étaient des reptiles. Les manchots à jugulaire sont
des oiseaux aquatiques.Les cétacés (dauphin) sont
en fait proches des hippopotames.Les siréniens (dugong) sont
en fait proches des éléphants.




La Théorie Synthétique de l'Evolution s'impose dans le monde scientifique comme théorie de référence. Dans les années 1970, elle connait d'importants développements théoriques, parfois iconoclastes. Par exemple, la théorie neutraliste de l'évolution moléculaire proposée par le généticien japonais Motoo Kimura en 1970, repose sur l'idée que l'évolution, au niveau des gènes, doit au moins autant sinon plus au hasard (c'est-à-dire à une dérive) qu'à la sélection naturelle.
- Argumentation : La plupart des mutations ne sont pas adaptatives. Une énorme partie des mutations est neutre ne procurant ni avantage, ni handicap, et une autre partie importante est handicapante. Les mutations procurant un avantage sont quasiment inexistantes. Cela est l'argument fort de la théorie neutraliste qui veut que le hasard ou la dérive génétique soit presque l'unique clé de l'évolution. La sélection naturelle ne jouerait qu'un rôle marginal selon la théorie neutraliste.
- Il est tout aussi dangereux d'adopter le seul point de vue « adaptationniste », selon lequel chaque caractère d'un être vivant pris isolément aurait été « façonné » par la sélection naturelle. Dans cette optique, tout devrait avoir une utilité adaptative, y compris la couleur des yeux, le chiasme optique et l'appendice.
L'expérience montre en fait qu'il existe beaucoup de caractères neutres en ce qui concerne la descendance. Certains gènes sont par ailleurs pléiotropes - c'est-à-dire qu'ils commandent plusieurs caractères à la fois. Une mutation avantageuse entraîne donc avec elle d'autres caractères ; quand la mutation se répandra dans la population, ses « effets secondaires » se répandront en même temps, sans avoir d'utilité en eux-mêmes... en tout cas à ce moment-là. Un exemple flagrant est celui de la drépanocytose, une maladie héréditaire, qui perdure parce qu'elle confère une certaine résistance au paludisme. La mutation ayant entrainé son apparition n'est pas neutre, mais la grave maladie qu'elle entraine est en quelque sorte compensée par un avantage sélectif, vis-à-vis d'une autre maladie.
Equilibres ponctués
 Podarcis sicula. Des lézards de l'espèce p. Sicula déposés sur l'ile de Prod Mrcaru en 1971 ont évolué en 36 ans de sorte à disposer d'un nouvel organe de digestion absent chez l'espèce d'origine : les valves sécales.
Podarcis sicula. Des lézards de l'espèce p. Sicula déposés sur l'ile de Prod Mrcaru en 1971 ont évolué en 36 ans de sorte à disposer d'un nouvel organe de digestion absent chez l'espèce d'origine : les valves sécales.En 1972, S. J. Gould et Niles Eldredge publient leur théorie des équilibres ponctués destinée à combler ce qui est pour eux une faille de la TSE : les paléontologues (comme Simpson) affirmaient en effet que l'on devrait voir les espèces se transformer toujours très graduellement, et que l'on devrait donc trouver des fossiles correspondant à toutes les étapes de la spéciation[11].
Cette thèse baptisée gradualisme phylétique peut se résumer sous l'expression répétée à plusieurs reprises par Darwin dans son Origine des espèces : "Natura non facit saltum" (La Nature ne fait pas de saut). A l'inverse, certains biologistes (comme Ernst Mayr) pensaient que les apparitions de nouvelles espèces correspondaient à des évènements rares et ponctuels, que l'on peut représenter sous forme d'un escalier, les paliers étant pour les périodes où une population ne subit aucun changement et les sauts correspondant à une modification rapide des caractéristiques qui aboutissent sur quelques générations à une nouvelle espèce.
Pour Gould et Eldredge, ce sont les biologistes qui ont raison ; les paléontologues auraient en fait mal interprété les fossiles. Le débat sera néanmoins ravivé avec la critique des paléontologues Simon Conway-Morris, Derek Briggs and Richard Fortey qui contredisent les interprétations de Gould sur les schistes de Burgess censés illustrer la thèse des équilibres ponctués. Leur analyse plus précise de ces fossiles montre certes une grande diversité anatomiques mais pas de quoi conclure qu'il y aurait là les traces d'un "saut" au moment de l'explosion cambrienne avec de nombreuses lignées qui auraient brutalement disparu du monde vivant tel que le prétendait Gould.
- Argumentation : Récemment, un cas d'évolution ultra-rapide a été enregisté[12].
Néo-Lamarckisme
- Lamarck et Charles Darwin décrivaient l'hérédité des caractères acquis (bien qu'ils n'emploient pas cette expression, la distinction entre caractères acquis et innés date de la fin du XIXe siècle). Darwin mentionne même dans la première édition de L'origine des espèces les « effets cumulatifs du dressage » de génération en génération chez des chiens d'arrêt (pointers). Darwin est en réalité le premier à proposer une théorie des caractères acquis, sous le nom de "théorie de la pangenèse", dans son ouvrage De la variation des animaux et des plantes sous l'action de la domestication. C'est August Weismann qui fera la démonstration, non de l'impossibilité de la transmission des caractères acquis, mais de l'impossibilité de la transmission des mutilations, soutenue à la fin du XIXe siècle par les néo-lamarckiens. La théorie de Weismann exclut la transmission des caractères acquis de manière purement théorique. La théorie synthétique postule aujourd'hui que les gènes ne peuvent être modifiés naturellement que par des mutations aléatoires, mais il s'agit, de l'aveu même de Francis Crick, d'un "dogme" de la biologie moléculaire.[réf. nécessaire]
- On croyait en conséquence, jusqu'à la fin du XXe siècle, que l'adaptation individuelle ne se transmettait pas. Toutefois, au début du XXIe siècle, de nouvelles expériences et observations ont rouvert la porte à l'hypothèse d'une transmission de l'adaptation individuelle dans certains cas (notamment la taille, par rapport aux conditions d'alimentation), non par la modification des gènes, mais par la modification de leurs conditions d'expression, et, par là, de leur niveau d'activité, avec toutes les conséquences. Ces phénomènes sont qualifiés d'épigénétique. En outre, on a découvert que si les mutations sont bien aléatoires, les probabilités qu'elles soient réparées sont dépendantes des conditions extérieures : les mécanismes d'auto-réparation et d'entretien de l'intégrité du génome sont sous contrôle de l'état des cellules, ce qui fait que les cellules laissent passer plus ou moins de mutations selon leur adaptation à leur milieu. Au final, l'adaptation acquise apparait bien comme un élément de l'hérédité.
Une autre approche de l'évolution est le néo-lamarckisme. L’action en faveur de Lamarck se traduit par l’organisation d’un congrès international à Paris en 1947 sous l’égide du CNRS sous le thème de « paléontologie et transformisme » dont les actes paraissent en 1950 chez Albin Michel. Il réunit nombre des plus grandes autorités françaises ; outre Grassé, y participèrent Lucien Cuénot (1866-1951), Pierre Teilhard de Chardin (1881-1955), Maurice Caullery...[13] Ceux-ci s’étaient alors opposés à des tenants du néo-darwinisme, brillants biologistes également, comme John Burdon Sanderson Haldane (1892-1964) et George Gaylord Simpson (1902-1984)[14]. Grassé fait l’éloge de Lamarck d’autres façons comme dans un article de l’Encyclopædia Universalis en affirmant que Lamarck a été injustement calomnié et qu’il doit être réhabilité[15].
- Agrumentation : La transmission de caractères acquis à la descendance, pourrait passer par un support épigénétique selon de récentes études[16].
Accroissement de la complexité au cours de l'évolution
Il reste difficile de mesurer la complexité dans le monde vivant. Elle est souvent estimée par le nombre de types cellulaires identifiables chez un organisme, mais cette estimation n'est pas, ou peu, corrélée à la taille du génome ou au nombre de gènes (complexité du génome) par exemple et ne saurait en aucun cas être une mesure indiscutable.[réf. nécessaire] Ainsi, le génome humain comprend 3 milliards de bases, le blé 12 milliards. Richard Dawkins signale également que les animaux à sang froid ont un génome plus long que des animaux de taille comparable à sang chaud.
Rapporté à la quantité d'êtres vivants, l'accroissement de la complexité semble être un phénomène marginal dans l'évolution. En effet, la plupart des espèces sont encore au stade unicellulaire (plus de 99 % des êtres vivants sont des bactéries[17]), seules quelques lignées ont évolué vers une complexification.
La sélection naturelle a tout de même favorisé, dans de nombreux cas, des organismes complexes. Chez les mammifères, on observe une tendance à l'accroissement de la taille du cerveau, et des comportements. On l'explique généralement par une « course aux armements » entre proies et prédateurs, ou à une co-évolution antagoniste, à cause d'une compétition perpétuelle dans les populations (voir la théorie de la reine rouge). Un système nerveux plus élaboré peut sauver la vie de l'animal. De même, des « fédérations de cellules », ébauches d'organismes pluricellulaires, ont pu dans certaines circonstances être plus efficaces que les bactéries isolées pour leur permettre de survivre. Cependant, il est plausible que l'accroissement de la complexité ait pu, dans certains cas, correspondre à un caractère neutre et être fixé par la dérive génétique[réf. nécessaire].
Si on parcourt l'histoire évolutive de l'ensemble du monde vivant on assiste à un accroissement de la complexité dans de nombreuses lignées: insecte, mammifères. Cette complexité semble liée à une meilleure adaptation des populations à un environnement fluctuant. En effet dans les espèces simples (bactéries) les effectifs des populations varient au gré des variations des conditions, ce qui est moins le cas pour les espèces plus complexes (humains, insectes sociaux). Par exemple , un organisme volumineux résiste mieux aux changements de température. Un comportement élaboré permet d'effectuer des réserves de nourriture, de prévoir les saisons.
Cependant, comme le rappelle Stephen Jay Gould là encore (op.cit.), le phénomène est biaisé en raison de ce qu'il nomme le mur de complexité minimale en dessous duquel on ne peut plus concevoir d'organisme viable et pouvant se reproduire. Le génome peut donc se complexifier autant que le lui permettent le temps et les ressources de l'environnement, mais n'a pas la latitude de se simplifier en deça de ce "mur". On est dans le phénomène bien connu en probabilités de marche au hasard avec une barrière absorbante pour les individus, et de ce fait réfléchissante pour l'espèce : ne sera préservé que ce qui ne la franchit pas.
Remises en question de la théorie
Les paléoanthropologues mettent en doute certains des aspects de cette théorie. Yves Coppens et Pascal Picq vont, entre autres, tenter de démontrer que l'évolution biologique a précédé l'évolution culturelle, dans le cas de notre espèce, et qu'en ce sens "Le développement technique et culturel dépasse le développement biologique"; c'est-à-dire que les explications biologiques des comportement culturels humains sont moins significatives que les explications sociales. Ils parleront aussi de "sélection culturelle", en opposition à la sélection naturelle supposément s'opérant. Une des thèses les plus importantes sur le sujet de l'explication biologique des comportements sociaux humains provient de E.O. Wilson, fondateur de la sociobiologie.
Notes et références
- ↑ Source David, P. & Samadi, S. (2000) La Théorie de l'évolution, Flammarion.
- ↑ Par exemple Aristote dans son Histoire des animaux
- ↑ Guillaume Lecointre, Hervé Le Guyader, Classification phylogénétique du vivant, éd. Belin, 2001, p. 43.
- ↑ Génomes et évolution - Innovations génétiques, CNRS, (page consultée le 2 juillet 2008).
- ↑ Parenté et diversité des organismes, Université Pierre-et-Marie-Curie, Paris, (page consultée le 2 juillet 2008).
- ↑ Neil Campbell & al., Biologie, éd. De Boeck, 2006, p. 516.
- ↑ Apparition de la vie, École Supérieure de Physique et de Chimie Industrielle, (page consultée le 2 juillet 2008).
- ↑ Cyril Langlois, L'Évolution montrée par les fossiles, École normale supérieure de Lyon, (page consultée le 2 juillet 2008).
- ↑ Charles Sullivan, Cameron Mcpherson Smith, Darwin : ce n'est pas une histoire de singe, Association française pour l'information scientifique, (page consultée le 26 avril 2008).
- ↑ Par exemple, les yeux des vertébrés et des céphalopodes sont très similaires alors même que l'ancêtre commun de ces taxons ne possédait pas de tels yeux. Les yeux de ces deux taxons sont donc des adaptations analogues au problème de la vision montrant, pour les adaptationnistes, que, malgré la diversité des espèces, les contraintes adaptatives sont suffisamment fortes pour contraindre la place laissée au "hasard". De même le degré de ressemblance entre le Thylacine qui est un marsupial, et le chien qui est un placentaire est tel qu'il faut être un spécialiste pour différencier leurs squelettes respectifs.
- ↑ Plusieurs scientifiques considèrent la théorie de la mutation rapide des espèces d'Hugo De Vries comme la base de cette théorie saltationniste.
- ↑ En 2004, une équipe scientifique dirigée par Duncan Irschick et Anthony Herrel soit allée sur l'ile de Prod Mrcaru en mer Adriatique afin d'observer un lézard que l'on connait bien puisque très présent en France sous le nom de lézard des ruines. Ce lézard avait en effet été introduit sur l'ile en 1971 par l'équipe du professeur Eviatar Nevo, un spécialiste en évolution. Celui-ci n'ayant pas pu suivre son projet personnellement du fait des conflits yougoslaves ayant entrainé une interdiction à l'expérience. Et c'est 36 années plus tard que, Duncan Irschick et Anthony Herrel découvrirent comme Podarcis sicula avait évolué de façon fulgurante, confirmant de façon frappante l'hypothèse des équilibres ponctués. Le lézard avait grandi, sa mâchoire devenue plus puissante, et il avait changé de régime alimentaire, d'insectivore il était devenu herbivore, et des valves sécales étaient apparues au niveau des intestins pour l'aider à digérer les herbes...
Voir site : [pdf](en)PNAS, vol. 105, n°12, pages 4792-4795 (25 mars 2008). - ↑ cf. Blanc (1990) : 238.
- ↑ cf. Blanc (1990) : 11.
- ↑ cf. Lestienne et Lapidus (2000) : 45.
- ↑ Par exemple, L’obésité serait non pas uniquement un effet direct touchant les individus atteints eux-mêmes mais également un effet transgénérationnel. Des données chez l'homme et chez l'animal semblent montrer que les effets d'une sous-alimentation subies par des individus pourraient en effet être transmises aux descendants. Des modifications épigénétiques (n’altérant donc pas l'ADN) n’étant pas effacées lors du passage par la lignée germinale sont désormais décrites chez plusieurs espèces. En fait, chez les plantes il existe une corrélation entre le niveau d'expression d'un gène et sa méthylation. Pareillement, chez les mammifères nous témoignons de la méthylation d'une séquence transposable qui est insérée à proximité d'un gène particulier. Le degré de méthylation d'un transposon pouvant enfin moduler l'expression du gène dans lequel il s'est inséré. (Revue : Cahiers de nutrition et de diététique ISSN 0007-9960 CODEN CNDQA8 ; Source : 2002, vol. 37, no4, pp. 261-272 ; Editeur : Masson, Paris, FRANCE (1966) ; Site : CAT.INIST du CNRS).
- ↑ Stephen, Jay Gould rappelle d'ailleurs, dans La vie est belle, que la biomasse bactérienne sur Terre est supérieure à celle des mammifères
 Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant
Catégories : Paléoanthropologie | Théorie sur l'évolution







Wikimedia Foundation. 2010.