- Selection de parentele
-
Sélection de parentèle
Les mécanismes de
l'évolution biologiqueMécanismes non aléatoires:
Mécanismes aléatoires:
Dynamiques évolutionnaires :
La sélection de parentèle est une généralisation de la théorie de l'évolution par voie de sélection naturelle de Darwin, soit une modification de l'équation générale de la valeur sélective ; la théorie de Darwin constitue un cas particulier de la sélection de parentèle où le coefficient de parentèle est nul. La sélection de parentèle, à la différence de la théorie darwinienne classique, permet d'expliquer l'existence de l'instinct grégaire chez les animaux.
Cette théorie fut développée en 1964 par le biologiste anglais William Donald Hamilton et le premier résultat théorique d'importance fut produit par le biologiste américain George Price en 1970 et publié dans Nature. La sélection de parentèle fut popularisée par le biologiste américain Edward Osborne Wilson dans son livre Sociobiology: The New Synthesis paru en 1975 et la première confirmation expérimentale de la théorie fut réalisée en 1976 par les biologistes américains Robert Trivers et Hope Hare et publiée dans Science. C'est John Maynard Smith qui donna pour la première fois[1] le nom de sélection de parentèle (kinship) à la théorie d'Hamilton.
La sélection de parentèle constitue la théorie standard de la biologie évolutive en ce qui concerne l'apparition des sociétés animales.
Sommaire
Histoire de la sélection de parentèle
Le darwinisme
Le mécanisme d'apparition des sociétés animales posa depuis le début de grandes difficultés à la théorie de l'évolution par voie de sélection naturelle de Darwin. Dans l'origine des espèces chap.7 p.236, Darwin déclare que les sociétés d'insectes sont : « Une difficulté particulière qui d'abord m'apparut insurmontable, et réellement fatale à ma théorie dans son ensemble ». Il contourne le problème en introduisant le mécanisme de sélection familiale et déclare p.237 : « Cette difficulté, bien qu'apparaissant insurmontable, est amoindrie ou, comme je le pense, disparaît quand il est rappelé que la sélection peut s'appliquer à la famille, aussi bien qu'à l'individu ».
La première guerre d'idée sur le sujet de l'altruisme dans la nature apparut entre le naturaliste anglais Thomas Henry Huxley et l'anarchiste russe Pierre Kropotkine. Huxley, en 1888, affirma dans The struggle for existence que « du point de vue du moraliste, la vie est au même niveau qu'un spectacle de gladiateurs...la vie est une guerre ouverte et au delà de la relation familiale, temporaire et limitée, la guerre hobbesienne des uns contre les autres est l'état normal de l'existence ». En 1899, Kropotkine répliqua dans L'entr'aide : « Pas de compétition ! [...] C’est le mot d’ordre que nous donnent le buisson, la forêt, la rivière, l’océan. « Unissez-vous ! Pratiquez l’entraide ![2] » [3] Cette pseudo-science, aux implications morales et politiques, allait ouvrir la voie à la justification du capitalisme et du colonialisme sauvage et d'autres positions politiques extrêmes par le darwinisme social d'un côté et au rejet de cette science capitaliste et bourgeoise de l'autre, favorisant l'apparition du lyssenkisme et entraînant un retard considérable de la Russie dans le développement de la théorie de l'évolution.
Le néodarwinisme
La mathématisation subséquente de la théorie de Darwin par la théorie synthétique de l'évolution allait poser plusieurs difficultés à la sélection de groupe (sélection familiale). En effet, introduire un mécanisme évolutif particulier, non basé sur la sélection des individus, phénotypiques ou génotypiques, semblait totalement artificiel. De plus, les quelques tentatives théoriques ne permirent aucunement d'expliquer clairement la formation des sociétés ni de réaliser des prédictions pouvant être vérifiées par des expériences, premier critère de la scientificité d'une théorie.
Haldane, Fisher et Wright, les pères fondateurs de la théorie synthétique, n'ont pas inséré l'altruisme au sein des équations de la sélection naturelle mais ils sont passés proche.
Dans son livre maintenant un classique The Causes of Evolution[4] et dans l'article Population genetics[5] Haldane s'intéresse à la relation entre l'altruisme et la parentèle dans le mécanisme de l'évolution naturelle. Il tente de trouver les fondements de l'apparition des comportements abaissant la valeur sélective propre d'un individu tout en augmentant celle des autres. Il arrive à la conclusion que ce type de comportement ne peut être sélectionné dans une grande population mais le pourrait dans une petite si les individus de ce groupe sont apparentés. Il établit également une relation entre l'application différentielle d'un comportement en fonction de la distance génétique et la probabilité que ce comportement soit sélectionné.
Fisher, après avoir réalisé plusieurs travaux sur l'évaluation de la proximité génétique entre les individus, présente dans son œuvre maîtresse The Genetical Theory of Natural Selection[6] un chapitre (the evolution of distastefulness) sur le phénomène du mauvais goût des larves d'insectes. En effet, comment l'évolution naturelle a t-elle pu sélectionner ce mauvais goût puisqu'il n'empêche aucunement le porteur de cette caractéristique de se faire dévorer. La seule explication qu'il trouve est que ce mauvais goût empêche ses frères et sœurs de se faire dévorer à leur tour. Une mutation peut donc être sélectionnée si elle favorise non pas directement un porteur mais ses apparentés.
Wright, ayant travaillé sur la mesure de la proximité génétique a également développé une théorie de la sélection de groupe. Malheureusement, il n'a jamais relié ces deux travaux d'une manière quelconque.
De la mathématique à la métaphore
Un des reproches habituels, formulé par les non-biologistes, envers la sélection de parentèle est que cette théorie prétend que la préservation du gène, plutôt que de l'individu, est la clé de l'évolution. Ce reproche est directement la conséquence de la métaphore de Richard Dawkins développée dans son best-seller Le Gène égoïste. Pourtant, le remplacement de la sélection des individus par les gènes est largement antérieur à la sélection de parentèle et est, en fait, la conception maintenant classique du néo-darwinisme.
Si une interprétation grossière de la théorie de Darwin pouvait laisser croire que la sélection des individus dans leur ensemble était la clé de l'évolution, le néo-darwinisme s'intéresse formellement (mathématiquement) au taux de reproduction différentiel produit par une mutation. Ainsi, une mutation est conservée (sélectionnée) si le taux de reproduction du porteur de la mutation est augmenté par celle-ci et elle est rejetée dans le cas contraire. Ici, c'est bel et bien la sélection des mutations qui est le moteur de l'évolution et non pas la sélection des individus.
Puisque, dans un environnement identique, toute mutation phénotypique (caractéristique physique ou comportementale) est causée par une mutation génétique, l'usage du terme mutation ou gène est habituellement utilisé dans le même sens sans ambiguïté. Notons que si nous voulions être plus précis, il faudrait parler de mutation génétique puisque le sens du terme gène est maintenant trop restrictif ; en particulier car l'épigénétique moderne révèle que des caractéristiques phénotypiques peuvent être véhiculées par des séquences d'ADN non associées à des gènes (introns, microARN, séquence cis et autres ADN non codant).
Si dans le néodarwinisme, la mutation n'était propagée que par l'individu, la sélection de parentèle attribue cette propagation à l'individu et aux apparentés. La sélection naturelle, au sens darwinien, se rétablit donc en considérant que la sélection s'effectue, simultanément, sur les individus (sélection individuelle) et sa famille (sélection familiale).
Le postdarwinisme
Hamilton commença sa licence de biologie à Cambridge vers la fin des années 1950. Passionné d'histoire naturelle, d'évolution des comportements et de génétique, il fut grandement déçu de ce qu'il constata à l'université. Malgré le fait que la synthèse moderne était en cours, «beaucoup de biologistes de Cambridge semblaient à peine croire en l'évolution naturelle ou restaient sceptiques sur l'efficacité de la sélection naturelle»[7]. Un jour, en étudiant à la bibliothèque St. Johns de Cambridge, il tomba sur le livre de Fisher The Genetical Theory of Natural Selection, il comprit immédiatement qu'il s'agissait de la clé pour comprendre l'évolution. Il devint un disciple de Fisher et se lança corps et âme dans la lecture de ce livre, oubliant tout, même ses cours[8].
Hamilton était inscrit à une majeure en génétique et le programme exigeait des cours facultatifs dans d'autres disciplines. Pour satisfaire son intérêt sur la question de l'altruisme social, il prit un cours d'anthropologie sociale avec le professeur Edmund Leach très attaché au modèle de la tabula rasa ; celui-ci vit d'un très mauvais œil qu'un étudiant s'intéresse à l'évolution génétique de l'altruisme. Les professeurs du département de génétique n'étaient pas plus chaud à cette idée et Hamilton fut vivement critiqué pour ce choix. Cet épisode le troubla profondément, jusqu'à reconsidérer une carrière de scientifique. Dans une lettre qu'il envoya à sa sœur en novembre 1959, il écrit : « Je commence à trouver Cambridge intolérablement oppressant... je pense que je renoncerai à l'espoir de réaliser un progrès malgré tout cela... » ; heureusement, ce ne fut pas le cas.
En 1963 et 1964, il publie dans l'indifférence totale (ses professeurs estimèrent qu'il ne méritait pas le titre de docteur qu'ils lui refusèrent jusqu'en 1968), les deux articles fondateurs[9] [10] de la sélection de parentèle dans lequel il expose sa fameuse équation et l'explication, qui en découle, de l'apparition de sociétés organisées chez les insectes. Son équation, toute simple, ne nécessite aucun recours à une quelconque forme de sélection de groupe.
C'est en 1965, au cours d'un voyage en train entre Boston et Miami, que le professeur Edward Osborne Wilson, grand spécialiste des insectes sociaux, découvre le travail d'Hamilton. Il le consulte avec admiration et incrédulité, cherchant des heures durant une faille dans le raisonnement ; arrivé à Miami il était convaincu : « Je dus admettre qu'Hamilton, qui connaissait infiniment moins de choses que moi sur les insectes sociaux, avait réalisé sur eux l'unique grande découverte de ce siècle.» [11]. Durant les années suivantes, Wilson exposa la théorie d'Hamilton au cours de nombreuses conférences.
En 1968, le généticien des populations George Price prit connaissance des travaux d'Hamilton et sombra en dépression ; sa fibre religieuse fut profondément ébranlée. Il remarqua, par contre, la puissance théorique de ce nouvel axiome. Il réutilisa des outils mathématiques développés par la biologie évolutive (étude de la covariance d'Alan Robertson) pour démontrer que l'équation d'Hamilton pouvait être déduite de la sélection de groupe et impliquait que des comportements malveillants intraspécifiques étaient naturellement sélectionnés dans les grandes populations. Ce fait théorique confirmait ce que l'on voyait partout dans le monde animal ; l'altruisme décroit de la famille immédiate au groupe (pour les animaux sociaux) pour devenir de l'agression systématique entre individus de groupes différents. Il publia cette conclusion mathématique remarquable dans Nature en 1970 [12] cet article fut immédiatement suivi, dans le numéro subséquent, d'un article d'Hamilton tirant toute la puissance explicative de cet apport fondamental[13]. Peu de temps après, Price se convertit au christianisme et, en 1975, après avoir donné toutes ses possessions aux pauvres, il se suicida. Peut-être voulait-il laisser un dernier message : l'homme peut transcender sa nature.
En 1971, la communauté scientifique internationale découvrait la théorie d'Hamilton grâce à la parution de The Insect Societies, qui est le premier grand ouvrage de Wilson. En 1975, en digne successeur de Darwin, voulant généraliser l'application de la théorie à l'ensemble du règne animal, y compris l'homme, il publie Sociobiology: The New Synthesis. Le dernier des vingt-sept chapitres, portant sur la formation des sociétés humaines, provoqua des remous comparables à ceux de L'origine des espèces ou La descendance de l’homme de Darwin, en particulier aux États-Unis et en France.
Le succès de la théorie est considérable, Hamilton est maintenant l'auteur dans le domaine de l'évolution des comportements le plus cité et il fut récompensé des prix les plus prestigieux de la biologie dont la médaille Darwin en 1988, la médaille linnéenne en 1989 et le Prix Crafoord en 1993. À sa mort le Guardian en dira : « Le plus grand innovateur de la biologie darwinienne moderne responsable de la forme de ce sujet aujourd'hui », le New York Times : « Une des plus grandes figures de la biologie moderne » et The Independent : « Un bon candidat au titre du darwinien le plus remarquable depuis Darwin ».
Wilson fut également récompensé par de nombreux prix entre autres la National Medal of Science, le Prix Crafoord, et deux fois le Prix Pulitzer. Il fut également élu membre étranger de la Royal Society en 1990. Le grand philosophe américain Charles Frankel dit à son propos : « Il appartient donc à la catégorie des grands visionnaires ; mais plutôt que dans la tradition de Marx ou de Spencer, il se situe dans celle de Descartes. [...] les visionnaires et les prophètes du premier jour s'acquittent d'une tâche indispensable. Même si les possibilités qu'ils envisagent se trouvent dans leur ensemble hors de portée, ils entrevoient des objectifs invisibles pour leurs contemporains. » [14]
La sélection de parentèle, une naissance trouble
En 1976, la revue anglaise Animal Behaviour demanda une analyse critique indépendante du livre Sociobiology: The New Synthesis de Wilson à quatorze sommités internationales ; seulement trois s'avérèrent négatives, toutes américaines [15]. Au cours des années suivantes, Wilson comprendra tout le sens du proverbe nul n'est prophète dans son pays. Alors que les différents cercles intellectuels mondiaux acceptaient volontiers la sélection de parentèle et que la première revue internationale de sociobiologie (Behavioral Ecology and Sociobiology, Springler-Verlag) était créé en Allemagne, le débat faisait rage au pays.
Le livre de Wilson était arrivé à un bien mauvais moment, juste vers la fin de la polémique sur l'héritabilité du quotient intellectuel. Il venait, tout à fait innocemment, de jeter de l'huile sur le feu. Ces débats de la côte est, où se mélangeaient politiques et sciences, avaient comme enjeux finaux la séparation des élèves du primaire et du secondaire en fonction de leur QI. Dans ce contexte, le dernier chapitre du livre de Wilson n'était pas le bienvenu d'autant plus qu'il fut lancé sur la place publique par un article élogieux, en première page, du New York Times. Le succès littéraire et scientifique du livre déclencha une guerre ouverte contre Wilson. Ses pires ennemis furent deux collègues de Harvard, le généticien Richard Lewontin et le paléontologue Steven Jay Gould, membres de Science for the People, un groupe de gauche qui défendaient bec et ongles la non-héritabilité du QI et tout ce qui aurait pu, de proche ou de loin, favoriser l'acceptation de l'idée contraire.
Une campagne de salissage fut menée contre Wilson, on déforma délibérément ses propos, on l'accusa de faire de la « pseudo-science » et de vouloir promouvoir les inégalités sociales par de fausses théories scientifiques. Ces attaques allaient tellement loin qu'en mars 1976, la revue Science publia un article spécial pour démentir les détracteurs et rétablir les faits[16]. Ces attaques continuèrent malgré tout jusqu'en février 1978 lors d'un symposium sur la sociobiologie organisé par l'association américaine pour l'avancement de la science. Lors de sa conférence, une extrémiste versa un récipient d'eau sur la tête de Wilson. L'assistance scandalisée rabroua les extrémistes vers la sortie et ovationna Wilson. Gould, au début de sa conférence, déclara désapprouver les méthodes mais partager le point de vue des extrémistes, il fut hué et l'audience quitta la salle. Ce fut le dernier épisode américain de cette saga. Malheureusement, le feu reprit quelques mois plus tard de l'autre côté de l'Atlantique.
Le peuple français découvrait la sociobiologie par les écrits de la Nouvelle droite et leur sociobiologisme prônant l'élaboration d'une morale sociale fondée, en grande partie, sur la biologie et, bien sûr, la sociobiologie. Ce courant promu par le journal populaire le Figaro (voir article détaillé) et dénoncé par Le Nouvel Observateur en 1979 allait déclencher un débat social houleux où la sociobiologie fut parfois amalgamée assez hâtivement à des thèses politiques.
En 1991, en partie en réponse à ces événements, la Société européenne de sociobiologie adopta une charte éthique mentionnant : « En accord avec ses statuts, la société s'abstiendra de tout us ou abus de ces études à des fins politiques. »
Le miracle de Harvard
Le conflit politique entre Wilson et Gould ne serait pas si confondant si, du point de vue scientifique, ils n'étaient pas également défenseurs de deux modifications distinctes de la théorie de Darwin ; les seules sérieuses à survenir depuis un siècle. Nous ferons remarquer que ces théories ne sont absolument pas contradictoires et se complètent parfaitement. C'est Gould qui inventera un schisme, de la même manière qu'il inventa de toute pièce le conflit politique avec Wilson.
En 1972, Gould expose avec Niles Eldredge la théorie de l'équilibre ponctué. Cette théorie postule que l'évolution comprend de longues périodes d'équilibre ponctuées de brèves périodes de changements importants comme la spéciation ou les extinctions.
En 1979, en se fondant sur la théorie neutraliste de l’évolution, il développe avec Richard Lewontin la théorie des trompes (spandrels). Cette théorie permet d'expliquer les radiations évolutives (Cladogénèse) décrites dans la théorie de l'équilibre ponctué. Cette théorie explique les périodes extrêmement rapides de spéciation par l'évolution neutre, c'est-à-dire, par la propagation de mutations à valeurs adaptatives nulles. Ainsi, pour Gould, la plupart des caractéristiques morphologiques et comportementales sont des décorations sans valeur sélective. La sempiternelle question de l'avantage évolutif perdrait donc, en général, tout son sens.
Cette théorie appuie le modèle de Konrad Lorenz de la spéciation par modification de la parade nuptiale. Si la parade nuptiale ou les mécanismes d'appariement sexuel entre un mâle et une femelle deviennent non-fonctionnels, mâle et femelle ne se reconnaissent pas comme appartenant à la même espèce. La parade nuptiale étant fortement décorative, il est fort probable que l'évolution de ce comportement se produise par un mécanisme de trompe.
Le cycle complet de l'apparition de nouvelles espèces animales comprendrait donc une extinction, suivie de la colonisation de niches écologiques maintenant libres par une espèce ayant résisté à l'extinction, puis une propagation de mutation décorative de la parade nuptiale dans cette population. Il se créerait donc ainsi naturellement des clines de la parade nuptiale entrainant rapidement la création de nouvelles espèces.
Gould croyait que l'évolution par trompe était un mécanisme fondamental et qu'un très grand nombre de comportements en découlait. Dans les faits, si elle permet d'expliquer la cladogénèse, elle est incapable de produire des prédictions expérimentales quantitatives vérifiables, ceci la laissant au simple statut de théorie explicative. La théorie de Gould explique bien les variations décoratives d'un même comportement mais elle est très faible en ce qui concerne la création de comportements nouveaux et d'intérêt nul en ce qui concerne l'instinct grégaire.
La théorie des trompes, utilisée en anthropologie, est en pratique inexistante en biologie et dans le cas qui nous intéresse, en éthologie. La lutte féroce qu'il mena, à l'encontre de la pensée largement dominante, contre l'application de la théorie de la sélection naturelle à l'homme était clairement teintée d'idéologie politique.
Gould, sans complètement nier la théorie de la sélection intraspécifique croit à «un type de sélection à un niveau supérieur sur des espèces elles-mêmes essentiellement statiques»[17]. Cette position est absente de la biologie contemporaine. Le biologiste Richard Dawkin en dit : «Une théorie de l'évolution (doit) expliquer des mécanismes complexes et bien conçus comme le cœur, la main, l'œil et l'écholocalisation. Personne, même le plus ardent partisan de la sélection interespèces ne pense que (celle-ci) puisse le faire (...) (Elle) n'est pas une force significative dans l'évolution de cette machine complexe qu'est la vie.»[18].
L'équation d'Hamilton
- Avant Hamilton, la valeur (adéquation ou fitness) sélective d'un individu était strictement équivalente à son taux de reproduction. Selon Hamilton, la valeur sélective d'un individu n'est pas seulement proportionnel à son succès personnel en matière de reproduction mais également à celui de ses tiers apparentés (génétiquement proches). La somme de la valeur sélective et de la valeur sélective indirecte est baptisé valeur sélective globale (inclusive fitness) par Hamilton.
- Un comportement sera qualifié d'altruiste envers un tiers s'il permet l'augmentation de la valeur sélective propre de ce dernier. Par conséquent, d'après le postulat (1), un comportement altruiste envers un tiers apparenté permet d'augmenter la valeur sélective globale.
À partir de ces postulats, Hamilton déduit l'équation suivante :
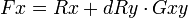
avec :
- Fx : la valeur sélective globale de l'individu x.
- Rx: la valeur sélective propre de l'individu x.
- dRy: l'effet de l'altruisme de x envers y sur la valeur sélective propre de y.
- Gxy: le coefficient de proximité génétique entre x et y.
Nous remarquerons que lorsque dRy = 0 nous obtenons la théorie darwinienne classique Fx = Rx ; soit que la valeur sélective globale est strictement égale à la valeur sélective propre, elle-même strictement équivalente au taux de reproduction (taux de reproduction différentiel dans le cas de la valeur sélective d'une mutation).
Il découle de cette équation que la sélection naturelle d'une mutation prédisposant à l'altruisme est favorisée (Fx > 0) si le coût de l'altruisme (perte de valeur sélective propre) est inférieur au gain que procure celle-ci via la valeur sélective indirecte. Cette contrainte est baptisée règle d'Hamilton et est formalisée par l'inéquation suivante :
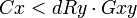
avec :
- Cx : la perte de valeur sélective propre de l'individu x causée par la mutation.
Il est possible de dégager les cas de figure favorisant la sélection d'une mutation prédisposant à l'altruisme :
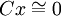
Si le coût de la mutation est extrêmement faible, la mutation sera sélectionnée dès qu'elle favorise le moindrement un apparenté, même très éloigné génétiquement.
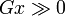
Si la proximité génétique est très grande, la mutation sera sélectionnée malgré un coût élevé et/ou un faible effet de l'altruisme. Le coefficient de parenté maximum étant 1, le cas le plus favorable est le clone (jumeau identique). Les organismes multicellulaires sont des sociétés cellulaires de clones.
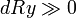
Si la mutation possède un effet altruiste très élevé, la mutation sera sélectionnée malgré un coût élevé et/ou un faible coefficient de parenté.
À la limite, une mutation pourrait conduire à la stérilisation des individus au profit d'une augmentation maximale de l'effet altruiste. Il serait également possible d'envisager l'apparition d'un comportement de sacrifice permettant un effet altruiste très grand.
N.B. Nous remarquerons que l'apparition d'une mutation prédisposant à l'altruisme implique, en général, la coexistence d'un mécanisme de reconnaissance de la proximité génétique (parentèle) entre les individus. En effet, il est implicitement contenu dans l'équation que l'effet (dRx) associé au coût (Cx) est réalisé par un comportement de x envers y. Pour un coût fixe, si le comportement était réalisé envers un individu possédant une distance génétique trop grande, celui-ci ne serait pas rentable.Groupe, individu et ADN
L'équation d'Hamilton induit un changement de perspective important. Il est maintenant impossible de conserver l'interprétation darwinienne de la valeur sélective. En effet, dans le cas de la stérilisation des individus comme ce qui se produit avec les hyménoptères sociaux, le taux de reproduction des porteurs d'une mutation de stérilisation est nul, ce qui entraine une contradiction. Par conséquent, il faut absolument reformuler la valeur sélective qui ne peut plus être le taux de reproduction de son porteur via la sexualité directe. Hamilton, pour résoudre le problème, introduit donc le concept de valeur sélective globale comme étant le taux de reproduction du porteur de la mutation par sexualité directe et indirecte via les apparentés.
Il serait également possible de considérer que la valeur sélective est simplement le taux de reproduction de la mutation elle-même, indépendamment du mécanisme utilisé pour réaliser la reproduction. Nous remarquerons que le taux de reproduction du porteur d'une mutation, par sexualité directe ou indirecte, est directement proportionnel au taux de reproduction de son ADN et de sa biomasse. La reproduction globale des individus, de la biomasse ou de l'ADN sont donc simplement trois façons distinctes d'interpréter le même phénomène naturel qu'est la reproduction du vivant. Par contre, du point de vue moléculaire, c'est bel et bien l'information contenue dans l'ADN qui a traversée les âges et non les individus ou leur masse.
Mais le plus remarquable est l'absence de la notion de groupe dans cette théorie. Le groupe émerge par l'application de comportements interindividuels eux-mêmes le résultat de la reproduction aveugle de mutations. La notion de sélection naturelle par « survie du groupe » popularisée par les premiers éthologues, en particulier Konrad Lorenz, était désormais tombée en disgrâce ; sa seule mention dans les publications spécialisées discréditait systématiquement leurs auteurs.
La reformulation de la théorie par Hamilton assisté de George Price au début des années 1970 allait donner la base théorique d'une lente réhabilitation de la sélection de groupe. Il est maintenant admis que la pression de sélection se réalise à tous les niveaux d'organisation du vivant, des cellules de l'organisme pluricellulaire (darwinisme cellulaire) aux écosystèmes, en passant pas les individus et les groupes.
Le paradoxe dégagé par l'analyse de Price est que l'altruisme n'évolue à l'intérieur d'un groupe que si celui-ci est en compétition avec d'autres groupes (sélection entre groupes) et ceci même s'il est composé exclusivement de parents. Si l'équation d'Hamilton continue de prédire dans quelles situations un comportement altruiste est sélectionné dans une population, elle n'est plus une alternative à la sélection de groupe qui, au contraire, lui est indispensable.
Une théorie, plusieurs modèles
Le modèle original d'Hamilton
Ce modèle est celui proposé par Hamilton[10] pour expliquer l'apparition de l'eusocialité chez les hyménoptères. Dans la nature, l'eusocialité ne se retrouve que chez les insectes sociaux, elle est caractérisée par :
- Superposition, dans une même société, de plusieurs générations d'adultes
- Forte cohésion des membres (échange d'information et de matière entre les individus)
- Division des rôles avec spécialisation des membres, certains pouvant être dédiés à la reproduction
- Élevage coopératif de la progéniture
Sur les 30 ordres d'insectes, seulement deux possèdent des espèces eusociales, les isoptères et les hyménoptères. De plus, les hyménoptères ont réinventé de façon indépendante (évolution analogue) l'eusocialité à 12 reprises au cours de l'évolution[19].
Le modèle d'Hamilton explique cette propension à la socialisation chez les hyménoptères par leur mécanisme sexuel hors norme : l'haplodiploïdie. En effet, de façon exceptionnelle, les mâles sont haploïdes alors que les femelles sont diploïdes. Par conséquent, la méïose ne se réalise que dans les gamètes femelles (ovules). Cette forme de sexualité implique des coefficients de proximité génétique très particuliers :
Mère Père Sœur Frère Fille 1/2 1/2 3/4 1/4 Fils 1/2 0 1/4 1/4 Le fait que deux sœurs possèdent une proximité génétique plus grande qu'avec leur propre descendance entraîne, selon l'équation d'Hamilton, que toute mutation favorisant la production de sœurs par une femelle, même au détriment de sa propre reproduction (fils ou fille), sera sélectionnée. Par conséquent, toute mutation altruiste envers la mère et l'empêchant de produire des fils sera sélectionnée. Nous remarquerons qu'une augmentation du ratio fille / fils produit par la reine ne diminue en rien sa valeur sélective globale mais augmente significativement celle de ses filles.
Ce modèle ne fut confirmé que treize ans plus tard par Trivers et Hare dans une publication de la prestigieuse revue Science (voir évolution par sexisme).
La manipulation parentale
Hamilton et Wilson croyaient initialement que les sociétés animales ne pouvaient apparaître que par des mutations altruistes. Pourtant, l'analyse mathématique de l'équation permet de prédire le contraire. Si Cx est négatif (le coût est en fait un gain), une mutation malveillante (dRx négatif) pourrait très bien être sélectionnée.
Cette simple constatation permet d'intégrer un modèle jugé initialement contradictoire, celui de la manipulation parentale de Richard Alexander et de Charles Michener qui l'on présenté, de façon indépendante, en 1974, pour expliquer respectivement l'évolution des sociétés de rat-taupes et des abeilles hypogées (ou halictes).
Dans ces sociétés, la femelle reproductrice unique domine et exclut de la reproduction les autres membres de la colonie par de fréquents comportements d'agression (malveillance de la reine envers les membres). Par contre, cette malveillance est largement compensée par les tâches de protection, recherche de nourriture et élevage de la progéniture effectuées par les membres au profit de la reproduction de la reine (altruisme des membres envers la reine).
Chez les rats-taupes il est démontré que les comportements d'agression sont inversement proportionnel au lien de parenté, ceci étant en parfait accord avec l'équation d'Hamilton.
L'évolution des sociétés animales peut donc utiliser des mutations altruistes, malveillantes ou encore un mélange des deux types.
Évolution par fratricide
La possibilité d'évolution sociale par apparition de comportements malveillants permet de revisiter la production de filles au détriment de fils par les reines d'hyménoptères sociaux. En effet, il est possible d'imaginer que la socialisation chez les hyménoptères ait évolué par fratricides de la part des sœurs. Il est en effet connu que les ouvrières se montrent souvent agressives envers les mâles et que les larves de mâles étaient les premières à être dévorées. De plus, en cas de situation critique, les ouvrières placent à l'abri en priorité les cocons femelles[20].
Évolution par sexisme
L'hypothèse radicale du fratricide n'est pas nécessaire, un investissement différentiel dans la production de femelles aux dépens des mâles pourrait très bien générer l'avantage sélectif global nécessaire. Trivers et Hare calculèrent[21] à l'aide de la théorie d'Hamilton, que le ratio minimum d'altruisme (soins parentaux) des ouvrières envers les femelles versus les mâles devrait être de trois (3).
Ils étudièrent ce ratio chez dix-neuf espèces de fourmis monogynes (possédant une seule reine par colonie). Seize espèces sur les dix-neuf avaient un ratio supérieur à 2,50 ; les valeurs se distribuant de 1,57 à 8,88 avec une moyenne de 4,36. Ce résultat est encore plus convaincant en sachant que la méthode utilisée ne permettait pas de distinguer entre les mâles produits par la reine de ceux produits par les ouvrières. En effet, dans plusieurs colonies, les ouvrières produisent une proportion importante des mâles.
Ce résultat constitua une première démonstration expérimentale de la sélection de parentèle. Ces prédictions furent largement confirmées dix ans plus tard[22].
La méthode expérimentale est très intéressante. Pour déterminer le ratio d'altruisme des ouvrières envers les mâles et les femelles, ils comparèrent la proportion d'aliments fournie aux mâles à celle fournie aux femelles. Pour ce faire, il s'agit simplement de calculer le rapport de biomasse femelle à la biomasse mâle, le poids des individus étant proportionnel à la quantité de nourriture ingérée.
Cette méthode utilise donc, indirectement, comme unité de l'altruisme la masse (Kg) impliquant une valeur sélective exprimée en variation de la masse par unité de temps. Ceci permet de rappeler que l'évolution de la vie est une évolution de la biomasse, soit une certaine masse de vivant produisant une autre masse de vivant, plus grande ou plus petite en fonction de la valeur sélective.
Le succès sélectif peut, en écologie, se mesurer en biomasse. La biomasse des fourmis est environ quatre fois supérieure à celle de l'ensemble des vertébrés terrestres[23].
La sexualité tardive
La théorie d'Hamilton permet de prédire que chez les animaux à maturité sexuelle tardive, une mutation poussant les individus sexuellement immatures à rester avec leurs parents pour les aider à élever leurs frères et sœurs serait sélectionnée.
Ce modèle fut présenté par le chercheur indien Raghavendra Gadagkar pour expliquer le comportement des guêpes Ropalida[24].
Le même raisonnement s'applique aux espèces où les individus peuvent atteindre la sénescence. Des mutations poussant les individus sénescents à participer à l'élevage de leurs descendances seraient sélectionnées.
Nous noterons que ces modèles pourraient très bien s'appliquer à Homo sapiens sapiens.
L'instinct de troupeau
La formation de troupeaux de mammifères, de bancs de poissons ou de colonies d'oiseaux chez les proies s'explique facilement par la théorie darwinienne classique. En effet, il est démontré que la fuite d'un groupe trouble les mécanismes cognitifs des prédateurs qui ont plus de difficulté à attraper une proie que si elle était seule. Une mutation favorisant le regroupement serait alors sélectionnée par simple augmentation de la valeur sélective propre.
Par contre, un autre comportement fréquent chez les proies, celui du déclenchement de l'alarme est plus difficile à expliquer à l'aide de la théorie classique. Ce comportement augmente de beaucoup la survie des membres en multipliant les organes des sens aux aguets ; des centaines de yeux, oreilles et nez en action permettent de détecter plus efficacement les prédateurs et donc, de fuir plus rapidement. De plus, la proportion de temps qu'un membre doit sacrifier à la surveillance au détriment d'autres activités comme l'alimentation est significativement diminuée. Ce comportement ne pouvait être expliqué, dans la théorie classique, que par la sélection de groupe. En effet, celui qui donne le signal se met délibérément en danger en se faisant remarquer. Par contre, en sachant que tous les apparentés du signaleur se trouvent dans le groupe, nous comprendrons aisément comment l'équation d'Hamilton résout le problème.
Un autre comportement de troupeau est la mobilisation du groupe en attaque collective contre un prédateur, autre comportement fréquent chez les proies, qui ne pouvait être expliqué dans la théorie classique que par la sélection de groupe. En effet, comment apparaît le premier mutant rentable ? Celui-ci doit affronter seul le prédateur avec une chance de survie pratiquement nulle. Ici encore, l'équation d'Hamilton permet de trouver une solution. Le sacrifice de l'individu X pour sauver un individu moyen Y (d'apparentement moyen) ne peut se réaliser que si Y possède un apparentement supérieur à 1 (ce qui est impossible) ou que le taux de reproduction propre de X soit nettement inférieur à celui de Y. Si la mutation se manifeste chez les individus trop vieux pour se reproduire, ce comportement n'affecte en rien leur taux de reproduction propre mais favorise les apparentés ; sous cette condition, l'équation d'Hamilton garantit la sélection de la mutation. Dans une société où les anciens se mobilisent, le risque d'être blessé ou tué en participant à la mobilisation diminue de façon importante ; le coût d'une mutation de mobilisation à tout âge devient alors rentable et elle peut maintenant être sélectionnée.
Confirmations expérimentales de la théorie
À la fin de la décennie 70-80, mise à part la confirmation expérimentale de Trivers et Hare, seule l'extraordinaire capacité explicative de la théorie permettait d'y adhérer. Pourtant, la méthode de validation empirique avait déjà été comprise par quelques chercheurs et exposée par Wilson dans Sociobiology: The New Synthesis. Cette méthode, toute simple, peut se résumer ainsi :
L'existence d'un mécanisme de reconnaissance des apparentés permettant à un animal de moduler un comportement de type altruisme-malveillance en fonction du degré d'apparentement ne peut s'expliquer avec la théorie darwinienne classique et nécessite la généralisation d'Hamilton. Autrement dit, un tel comportement ne pourrait être sélectionné dans le cadre de la théorie classique.
Avant la publication d'Hamilton (1964) aucun éthologue n'avait jamais étudié les mécanismes de reconnaissance des apparentés et c'est seulement en 1979 que fut publié, dans Science, le premier article associant reconnaissance de l'apparentement et altruisme[25] ; en 1980 on comptait moins de dix articles sur ce sujet[26]. De nos jours on en dénombre plusieurs centaines et aucun éthologue ne s'investirait dans l'étude d'une espèce sans s'intéresser aux mécanismes de reconnaissance des apparentés en relation aux comportements altruistes et malveillants.
La confirmation expérimentale est époustouflante, l'existence du mécanisme est démontré pour presque tous les groupes zoologiques des amibes aux mammifères en passant par les cœlentérés, insectes, poissons, amphibiens, oiseaux, etc. De plus, pour plusieurs espèces, la précision des mécanismes atteint parfois un niveau inespéré allant jusqu'à la distinction des frères des demi-frères, des demi-frères des neveux et ces derniers des cousins germains et ceci sans rencontre préalable des sujets. Plusieurs ouvrages spécialisés sont maintenant dédiés à la reconnaissance de l'apparentement[27].
Cas de l'amibe acrasiale Dictyostelium discoideum
Dans Sociobiology: The New Synthesis p.128 Wilson expose une méthode de validation expérimentale chez les mycétozoaires dont l'intérêt théorique « n'a rien à envier aux vertébrés ou aux insectes ». En effet, ces organismes, au carrefour évolutif des protistes, plantes, animaux et champignons possèdent plusieurs caractéristiques remarquables.
Les amibes acrasiales, bien que passant normalement leur vie sous une forme d'amibe bien ordinaire peuvent, en cas de dégradation de leur milieu, s'unir et former un organisme pluricellulaire : la limace amibiale mesurant deux millimètres. Ce pseudo-organisme se déplace en rampant vers la lumière. Une fois à la surface du substrat, la colonie va former un sporocarpe ressemblant à ceux des champignons myxomycètes, soit un pédicelle surmonté d'une capsule contenant des amibes enkystées appelées spores. Seuls les spores pourront survivre, les amibes composant le pédicelle sont condamnées à mourir.
Wilson, dans un élan prophétique, suppose en se basant sur la théorie d'Hamilton que la longueur du pédicelle devrait être proportionnelle à la proximité génétique moyenne des amibes formant la colonie. En effet, plus la proximité génétique est grande, plus qu'il devrait y avoir d'amibes prêt à se sacrifier, par acte altruiste, pour les membres de la capsule. Ainsi, le rapport longueur du pédicelle / diamètre de la capsule est une mesure du degré d'altruisme moyen de la colonie et devrait augmenter pour les colonies dont les membres sont génétiquement proches.
C'est seulement en 1990, quinze ans plus tard, que fut réalisé la confirmation expérimentale de la prédiction de Wilson[28]. Kolmes et ses collègues mélangèrent deux cultures d'amibes acrasiales pour concevoir une colonie mixte. Comme prévu, celle-ci développa des sporocarpes à grandes capsules et à petits pédicelles laissant entrevoir que même les amibes sont capables de distinguer leur apparentement.
Cas de la plante Cakile edentula
Si le phénomène de la reconnaissance entre apparentés est connu depuis maintenant longtemps chez les animaux, c'est seulement en 2007 qu'un tel mécanisme fut découvert chez une plante, le Caquillier édentulé (Cakile edentula) ou fusée des mers. Dudley et son étudiante comparèrent la masse des racines de plans voisins apparentés ou non et mirent en évidence que les plans voisins non-apparentés entrent en compétition et produisent plus de racine que les plans apparentés [29].
Critiques à l'encontre de la sociobiologie humaine
Le terme sociobiologie est souvent utilisé à tort pour exprimer l'application de l'ethologie à l'étude de l'homme ou plus simplement toute affirmation de l'importance de l'instinct dans le comportement de l'homme civilisé. Différentes formes de critique ont vu le jour :
Douance et déterminisme génétique
Le phénomène de la douance cognitive et la volonté de séparer les enfants doués de ceux qui le sont moins fut un champ de bataille politique important des années 1970. L'argument fort des pro-séparations était que le QI est une caractéristique génétique et que l'éducation ne peut rien faire pour l'améliorer. Les anti-séparations ont donc tenté par tous les moyens de démontrer que les facultés cognitives ne sont pas innées, pas plus que le moindre comportement humain.
Cette attitude était extrêmement risquée ; elle ne prévoyait aucune mesure si le facteur génétique du QI s'avèrait scientifiquement non-négligeable. Ainsi, au lieu de défendre le droit inaliénable à la non discrimination génétique, elle lui laissait la porte grande ouverte. De nos jours, bien que la charte universelle des droits de l'homme protège, théoriquement, contre la discrimination de l'état envers les détenteurs d'attributs « de naissance », peu de pays ont mis en place des mesures de protection contre la discrimination d'organismes privés envers le public. Présentement, les tests de dépistage de maladies génétiques en ce qui concerne les assurances vies sont sujets à une importante polémique et l'arrivée des puces à ADN dans le domaine du marché du divertissement est des plus troublants. Actuellement s'il est possible, pour une femme, de choisir l'avortement après une amniosynthèse si les tests génétiques révèlent une maladie que dire de son extension potentielle à des critères désavantageux comme des gènes révélant une possibilité de QI trop faible (comme le CHRM2 [30]). Il en est de même en ce qui concerne la sélection d'embryons par diagnostic préimplantatoire qui constitue, pour l'heure, la forme la plus poussée d'eugénisme.
La perte de la liberté ?
Le premier obstacle de la sociobiologie humaine est le sous-entendu du déterminisme génétique qui laisse croire que l'humain perd toute liberté de choix et que la culture est nécessairement assujettie à la nature (l'instinct). Les tenants du tout culturel défendent habituellement leur position par l'affirmation que les sociobiologistes croient à une conception du tout instinctif chez l'homme, ce qui est parfaitement faux. Ils oublient également souvent une vérité scientifique fondamentale : dans la nature, les cas de transmission culturelle sont des exceptions pratiquement miraculeuses, très étudiées et documentées ; même chez nos plus proches cousins les grands singes. Seul l'homme est un animal culturel et celui-ci n'intéresse pas particulièrement les biologistes.
Pour un tenant du tout culturel, le comportement humain est entièrement déterminé par l'expérience personnelle : la socio-culture, l'éducation, etc. La distinction qu'il est possible d'établir entre le déterminisme culturel et le déterminisme génétique est que le premier n'en est pas vraiment un puisqu'il peut changer rapidement, contrairement à l'instinct. Mais refuser l'existence de l'instinct chez l'homme par croyance qu'il implique un déterminisme rigide, outrepassant le déterminisme culturel est absolument absurde. Pour Konrad Lorenz, seul la transcendance de l'instinct par la culture pourra sauver l'homme de l'inadéquation de son instinct pour la survie de l'espèce. S'il considère que l'humain possède des tendances naturelles, celles-ci ne sont certainement pas nécessairement bonnes ni adaptées à la situation actuelle[31].
Mais dans tous les cas une vérité philosophique demeure : il est impossible de transcender ses déterminismes si nous ne les connaissons pas, notre ignorance de ceux-ci implique que nous nous laissons aveuglément conduire par eux. Sans connaissance, point de liberté.
L'idéalisation de l'homme (ou de soi-même) ?
Comme le souligne si bien Konad Lorenz : « Beaucoup d'entre nous ont quelque difficulté à reconnaître que l'évolution des civilisations humaines vers un niveau supérieur n'est en aucune façon commandée exclusivement par les jugements de valeur, l'intelligence et la bonne volonté de l'homme. »[31] p.268. En effet, pour certains, la distinction entre l'homme et les autres animaux est tellement profonde que le simple fait de rappeler que celui-ci en est un et qu'il possède des instincts comme tous les autres est blasphématoire. Si le darwinisme en plus de rabaisser le corps de l'homme à celui d'un singe s'attaque également à son esprit, il est naturel, par simple mécanisme de protection de l'égo, que l'on essaye d'y échapper.
Par contre, même si épistémologiquement, il est facile d'établir un parallèle entre ce phénomène de rejet et celui du copernicanisme ou du darwinisme, il est en fait beaucoup plus profond car il ébranle les bases fondamentales de la conception religieuse de l'esprit humain. La sociobiologie n'est pas qu'une des nombreuses facettes de la science matérialiste athée, il en est une affirmation triomphante.
La sociobiologie : politique de droite ?
La transmission génétique des avoirs (la transmission à sa descendance de ses richesses par héritage) est un fait connu et habituel de l'humanité depuis l'aube de la civilisation. Par contre, affirmer qu'il s'agit d'une conséquence évidente et directe de l'évolution génétique de l'homme est une autre histoire. En effet, le contrôle sévère de la transmission génétique des avoirs est ce qui distingue le simple socialisme du communisme. L'affirmation sociobiologique laisse penser que le communisme est contre nature et donc simplement voué à l'échec. Dans les années 1970, le marxisme léniniste et maoïste étaient des idéologies populaires dans les milieux des sciences humaines, il est facilement compréhensible que les affirmations sociobiologiques en aient troublés plusieurs. Par contre, malgré l'existence d'une tendance naturelle, il est rationnellement possible de considérer que de laisser le contrôle de la destiné de l'humanité à quelques familles de privilégié n'est pas une alternative viable et que le peuple doit posséder tous les leviers économiques pour un devenir meilleur.
Le simple socialisme par redistribution de la richesse n'est pas plus favorisé par l'équation d'Hamilton. En effet, celle-ci prévoit un altruisme nul entre les individus des pays civilisés. Pour les sociobiologistes, il s'agit simplement d'une preuve qu'il existe, ici, d'autres mécanismes instinctifs qui entrent en jeu comme le modèle de l'altruisme réciproque[32] de Robert Triver qui ne nécessite aucune affinité génétique entre le donneur et le receveur. Mais la réalité évolutive est probablement révélée par le cinquième principe de la psychologie évolutionniste, le mismatch, qui rappelle que les instincts de l'homme furent sélectionnés dans des conditions écologiques bien antérieures à la civilisation. Notre instinct grégaire serait en fait conçu pour de petits groupes, possédant une grande proximité génétique.
Si la gauche prône une redistribution de la richesse (altruisme social) et la droite une autonomie financière des individus, il est connu, qu'en général, seule l'extrême droite est xénophobe. Il semble donc exister un lien entre volonté de partage et l'impression psychologique de la disparité génétique de la population. Cette corrélation existe-t-elle réellement? Serait-il possible qu'il existe une relation entre la part de l'impôt utilisé en redistribution de la richesse et la disparité génétique des populations? Serait-il possible que les pays sans immigration comme la Suisse soient socialement beaucoup plus altruistes qu'un melting pot comme les États-Unis? Nos politiques seraient-elles influencées par de bas instincts? Rejeter ces questions du revers de la main n'est certainement pas une attitude scientifique. Par contre, ces questions n'appartiennent pas à la biologie mais à la sociologie et à la science politique ; répondre de façon scientifique est nécessaire pour l'avancement de la connaissance et ne concerne en rien l'idéologie politique. Si les héritiers de Gaston Bachelard veulent réussir à être pris au sérieux sur cette question par les biologistes, ils doivent impérativement abandonner l'épistémologie au profit du vulgaire empirisme comme le fait si bien toute science respectable.
La sociobiologie : science raciste ?
L'histoire de l'humanité est pavée de guerres, de massacres et de génocides. Ici, le comportement de l'homme civilisé avec son gros cerveau de plus de cent milliards de neurones ne se distingue guère de celui des cloportes réaumur n'en possédant que dix mille. Pour les sociobiologistes, il est clair que l'instinct d'agression inter-groupe chez l'humain fut sélectionné par les mêmes mécanismes évolutifs (équations de Price) que pour tous les autres animaux. Cette conséquence mathématique fut malheureusement confondue, au début de la sociobiologie, avec une tendance naturelle au racisme, soit au rejet voire à l'agression naturelle entre les membres des différentes races. Remarquons que les tensions sociales en Amérique, à l'époque, étayaient cette thèse. Heureusement, la réalité biologique est toute autre.
Chez tous les animaux sociaux, les mécanismes de reconnaissance de l'appartenance au groupe sont, en général, indépendants de marqueurs génétiques. En fait, l'évolution a simplement sélectionné tout mécanisme permettant que des individus cohabitant sur un territoire restreint se reconnaissent comme membres du même groupe. Nous remarquerons que dans la nature cette condition est toujours remplie par des apparentés. Il est donc possible, pour un expérimentateur, de recréer des groupes de cloportes, de fourmis ou de rats qui ne possèdent aucun apparentement, il en est de même chez l'homme.
L'étude des street corner societies (gangs de rue) a permis à la sociologie de dégager certains des mécanismes nécessaires permettant l'identification des membres des différents groupes entre eux. Il existe un parallèle surprenant entre ces mécanismes et ceux des sociétés primitives étudiées par l'ethnologie. Nous remarquerons que le critère de l'habillement et de la parure vestimentaire est fondamental et si la plupart des sociétés ont adopté l'habit occidental, parfois imposé par la force comme au japon, il existe encore une grande diversité vestimentaire sur la planète. Il est également significatif que l'imposition du communisme à une nation de plus d'un milliard d'habitants comprenait un vêtement unique pour toute la population. De même, l'habit imposé dans certains corps de métier ou dans les écoles privées a également cette fonction de construction artificielle d'un groupe d'appartenance.
Il existe plusieurs marqueurs de l'appartenance au groupe, interagissant, et créant par synergie un sentiment d'appartenance plus fort. Ces stimuli doivent également obéir à la règle de l'hyperstimulation de Konrad Lorenz et la connaissance de ces mécanismes est fondamentale pour la compréhension de la société et les applications sociales qu'ils suggèrent.
Sociobiologistes célèbres
- William Donald Hamilton
- George Price
- Robert Trivers
- Edward Osborne Wilson
- Richard Dawkins
- John Maynard Smith
Bibliographie
- Bibliographie de base
- Pierre Jaisson, La Fourmi et le Sociobiologiste, Paris, Odile Jacob, 1993. (épuisé)
- Michel Veuille, La Sociobiologie (Que sais-je ? N° 2284), Paris, P.U.F. (édition 1986 et 1997 épuisés)
- Richard Dawkins, Le Gène égoïste, Paris, Odile Jacob, 2003.
- Edward Osborne Wilson, L'Humaine Nature, Paris, Stock, 1979. (Prix Pulitzer)
- Edward Osborne Wilson, La Sociobiologie, Monaco, Rocher, 1987.
- Yves Christen, L'Heure de la sociobiologie, Paris, Albin Michel, 1979.
- Thèse de David Chavalarias (C.R.E.A, novembre 2004) Métadynamique en cognition sociale. Quelle définition de meilleur est la meilleure ?
- Bibliographie critique de la sociobiologie
- Marshall Sahlins, Critique de la sociobiologie : Aspects anthropologiques, Paris, Gallimard, 1980.
- Patrick Tort (dir.), Misère de la sociobiologie, Paris, PUF, 1985.
- Richard C Lewontin, Steven Rose, Leon J Kamin, Nous ne sommes pas programmés, Paris, La Découverte, 1985.
Notes et références
- ↑ Maynard Smith, J. (1964) Group selection and kin selection Nature 201:1145–1147
- ↑ Pierre Kropotkine, L'Entr’aide, Un facteur de l’évolution.
- ↑ Mentionné dans Nice guys finish first, document vidéo avec Richard Dawkins.
- ↑ Haldane J.B.S., The Causes of Evolution (1932), Longmans Green, London.
- ↑ Haldane J.B.S., Population genetics (1955), New Biology, 18: 34–51.
- ↑ Fisher, R.A., The Genetical Theory of Natural Selection (1930), Ed. 1. Dover, New York.
- ↑ Hamilton, W. D., Narrow Roads of Gene Land: The Collected Papers of W. D. Hamilton, Volume 1: Evolution of Social Behavior. (1996), W. H. Freeman, Oxford
- ↑ Hamilton, W. D., Narrow Roads of Gene Land: The Collected Papers of W. D. Hamilton, Volume 2: The Evolution of Sex (2001), W. H. Freeman, Oxford
- ↑ Hamilton , W. D.,The evolution of altruistic behavior (1963), American Naturalist, no.97 p.354–356
- ↑ a et b Hamilton W.D., The genetical evolution of social behaviour (1964), Journal of Theorical Biology, no.7, p.1-52
- ↑ E.O. Wilson, In the Queendom of the Ants : A Brief Autobiography (1985), Bucknell University Press, Cranbury, NJ, États-Unis
- ↑ Price G. R., Selection and covariance (1970), Nature, no.227, p.520–521
- ↑ Hamilton W., Selfish and spiteful behaviour in an evolutionary model (1970), Nature, no.228, p.1218–1219
- ↑ C. Frankel, La sociobiologie et ses critiques (1979), Sciences et tension sociales, Bulletin du Groupe de recherche sur l'histoire du racisme (CNRS), no.1, p.7-1
- ↑ Multiple review of Wilson's sociobiology (1976), Animal Behaviour, no.24 p.698-718.
- ↑ N. Wade, Sociobiology : troubled birth for new discipline (1976)
- ↑ Stephen Jay Gould, Le Pouce du panda (1982), Les grandes énigmes de l'évolution, Grasset, p.13
- ↑ Richard Dawkins, L'Horloger aveugle, pp. 308-313
- ↑ E.O. Wilson, The Insect Societies (1971), Harvard University Press, Cambridge
- ↑ P. Nonacs, N. Carlin, When can ants discriminate the sex of brood? A new aspect of queen-worker conflict (1990), Proceedings of the National Academy of Sciences of the USA, no. 87, p.9670-9673
- ↑ R.L. Trivers, H. Hare, Haplodiploidy and the Evolution of the Social Insects (1976), Science, no.191, p.249-263
- ↑ P. Nonacs, Ant reproductive strategies and sex allocation theory (1986), Quarterly Review of Biology, no.61, p.1-21
- ↑ E.O Wilson, Success and Dominance in Ecosystems : The Case of the Social Insects (1990), Ecology Institute, Oldendorf
- ↑ Gadagkar R., Demograhic predisposition to the evolution of eusociality : a hierarchy of models (1991), Proceedings of the National Academy of Science of the USA, no.88, p.10993-10997
- ↑ Greenberg, « Genetic component of bee odor recognition », Science, no.206, 1979
- ↑ Waldman B., Frumhoff P.C., Sherman P.W., «Problems of kin recognition», Trends in Ecology & Evolution, no.3, 1988
- ↑ « Kin Recognition », Edited by Peter G. Hepper, The Queen's University of Belfast, 1991
- ↑ M.J. De Angelo, V.M Kish, S.A. Kolmes, Altruism, selfishness, and heterocytosis in cellular slime molds, Ethology, Ecology & Evolution, no.2, p.439-443, 1990
- ↑ Susan A. Dudley, Amanda L, File, Yes, kin recognition in plants!, Biology Letters, février 23, 2008
- ↑ Dick DM, et al. Association of CHRM2 with IQ: Converging Evidence for a Gene Influencing Intelligence, Behavioral Genetics, vol.37, no.2, mars 2007, pp. 265-272
- ↑ a et b Konrad Lorenz, L'home en péril (1985), Flammarion.
- ↑ Trivers R.L., The evolution of reciprocal altruims (1971), Quaterly Review of Biology, no.46, p.35-47.
Voir aussi
Articles connexes
- Biologie, génétique, neurosciences
- Évolution
- Conflit sexuel
- Mémétique
- Psychologie évolutionniste
- Dilemme du prisonnier
- Phytosociologie
Liens externes
- www.sociobiologie.com : l'origine biologique du comportement social, Barhouf, 2003
- The Sociobiology of Sociopathy, Mealey, 1995 (www.bbsonline.org)
 Portail de la sociologie
Portail de la sociologie
Catégories : Écologie comportementale | Champ connexe à la psychologie | Comportement de l'insecte | Théorie sur l'évolution
Wikimedia Foundation. 2010.