- Acide Désoxyribonucléique
-
Acide désoxyribonucléique

« ADN » redirige ici. Pour les autres significations, voir ADN (homonymie).  Structure de la molécule d'ADN.
Structure de la molécule d'ADN.
L’acide désoxyribonucléique ou ADN[1] est une molécule, retrouvée dans toutes les cellules vivantes, qui renferme l'ensemble des informations nécessaires au développement et au fonctionnement d'un organisme. C'est aussi le support de l'hérédité car il est transmis lors de la reproduction, de manière intégrale ou non. Il porte donc l'information génétique, il constitue le génome des êtres vivants.
L'ADN détermine la synthèse des protéines.
Dans les cellules eucaryotes, l'ADN est contenu dans le noyau et une petite partie dans la matrice des mitochondries ainsi que dans les chloroplastes. Dans les cellules procaryotes, l'ADN est contenu dans le cytoplasme. Certains virus possèdent également de l'ADN dans leur capside.
Fonctions
L'ADN est une macromolécule, c'est-à-dire une très grosse molécule, dont la structure et les propriétés chimiques lui permettent de remplir toutes ces fonctions:
- Sa fonction principale, bien connue du grand public, est de stocker l'information génétique, information qui conditionne le développement et le fonctionnement d'un organisme. Cette information est contenue dans l'enchaînement non-aléatoire de nucléotides.
- Une autre fonction essentielle de l'ADN est la transmission de cette information de génération en génération. Cela permet l'hérédité.
- L'information portée par l'ADN peut se modifier au cours du temps. Cela aboutit à une diversité des individus et à une évolution possible des espèces. Cela est dû à des mutations dues principalement à des erreurs lors de la réplication des séquences de l'ADN (ajout, délétion ou substitution de nucléotides), ou bien à des recombinaisons génétiques.
L'ADN est donc le support de l'information génétique mais aussi le support de ses variations. Théoriquement, en subissant les effets de la sélection naturelle, l'ADN autorise l'évolution biologique des espèces.
Historique de sa découverte
 Modèle métallique utilisé par les découvreurs, en 1953
Modèle métallique utilisé par les découvreurs, en 1953La caractérisation et la découverte de la structure chimique de l'ADN se sont faites en plusieurs étapes.[2]
En 1869, le Suisse Friedrich Miescher isole une substance riche en phosphore dans le noyau des cellules, qu'il nomme nucléine (du latin nucleus, le noyau)
En 1889, l'Allemand Altmann sépare à partir de la nucléine, des protéines et une substance acide, l'acide nucléique.
En 1896, l'Allemand Kossel découvre dans l'acide nucléique les 4 bases azotées A, C, T, G.
En 1928, Levene et Jacobs (USA) identifient le désoxyribose. En 1935, on parle alors d'acide désoxyribonucléique.
En 1944, l'américain Avery découvre que l'ADN est responsable de la transformation génétique des bactéries, et que ce serait bien le support de l'hérédité[3]. Mais, certains scientifiques restent sceptiques, et n'abandonnent pas l'idée que les protéines puissent porter l'information génétique. En 1952, l'expérience de Hershey et Chase invalide définitivement cette dernière hypothèse.
- Découverte de la structure[4]
C'est au laboratoire Cavendish de Cambridge, qu'a été établie la structure en double hélice de l'ADN, grâce à la technique de diffraction des rayons X [5], qui est publiée dans Nature, le 25 avril 1953. On doit cette découverte à James Watson, alors âgé de 25 ans, Francis Crick, physicien de formation, et Maurice Wilkins qui reçurent le prix Nobel de physiologie et de médecine, le 31 octobre 1962. Mais leur découverte a été aussi rendue possible par le travail de Rosalind Franklin, qui mourut avant l'attribution du prix Nobel.
Ils s'appuyèrent sur un fait déjà établi : pour une espèce donnée les quantités de A et T sont sensiblement égales, ainsi que pour les quantités de C et G . Exemple chez l'homme : A=30,4 % & T=30,1 % ; C=19,6 % & G=19,9 %. Ce sont les règles d'équivalence de Chargaff (1949). Cela leur a suggéré la complémentarité des bases.
Rosalind Franklin obtint des clichés par diffraction aux rayons X de cristaux d'ADN, qui indiqua la structure en double hélice, ainsi que la distance entre les bases azotées.
En combinant ces données, James Watson et Francis Crick ont construit avec des tiges métalliques, le premier modèle en double hélice de l'ADN.
En 1959, le prix Nobel de physiologie ou de médecine est décerné à Severo Ochoa de Albornoz et à Arthur Kornberg pour la découverte du mécanisme biologique de la synthèse de l'acide désoxyribonucléique.
Structure
Aspect général et localisation
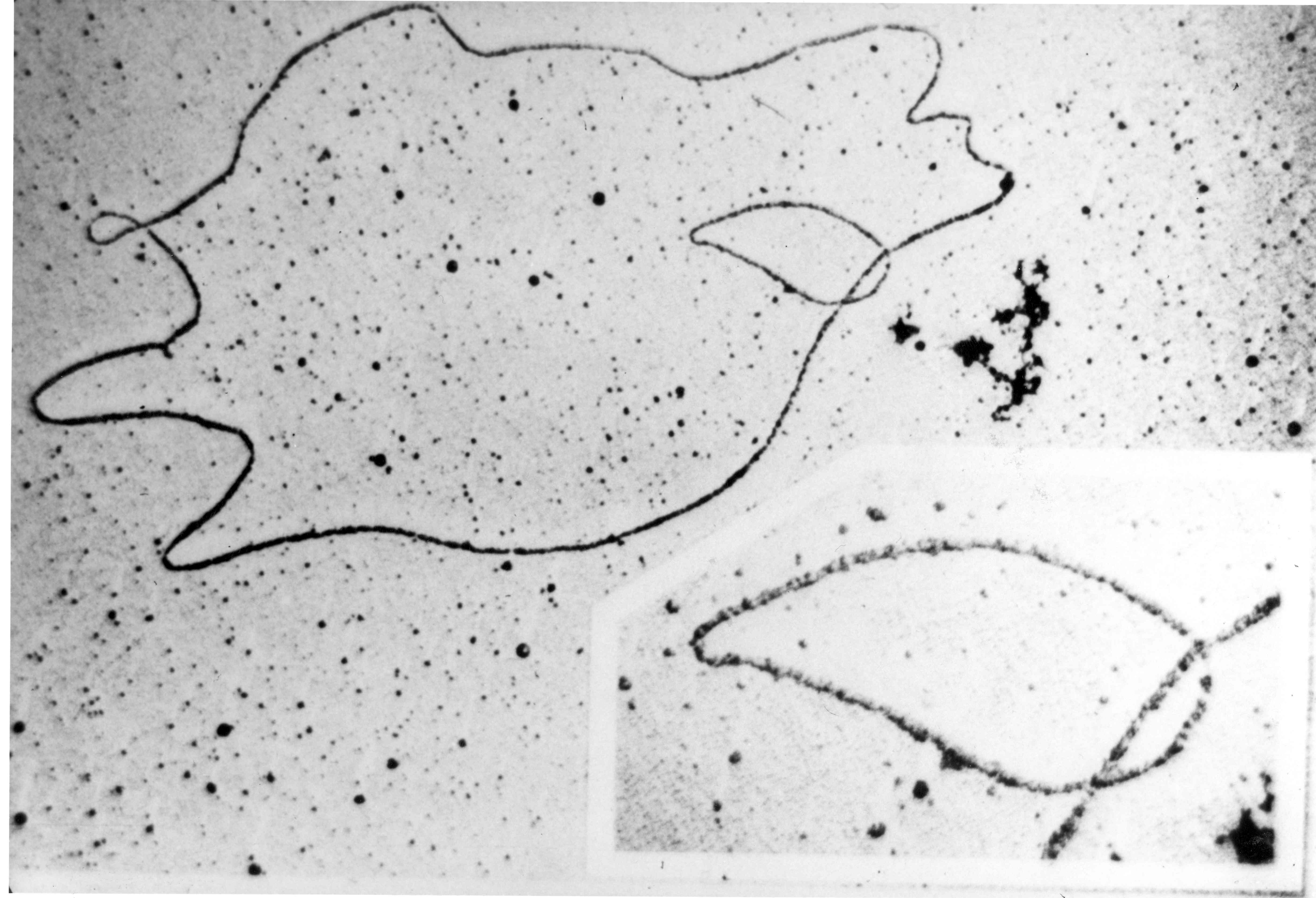
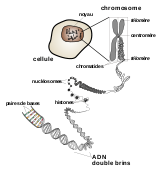 La structure et localisation d’un chromosome eucaryote.
La structure et localisation d’un chromosome eucaryote.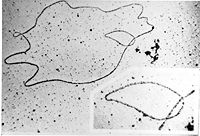 Deux images d'ADN circulaires bactériens observés au microscope électronique.
Deux images d'ADN circulaires bactériens observés au microscope électronique.L'ADN est une molécule allongée, pouvant mesurer plusieurs centimètres de long[6]. L'ADN peut être soit linéaire, soit circulaire:
Chez les procaryotes (organismes unicellulaires sans noyau), tels que les bactéries, l’ADN est en général présent sous la forme d’un seul chromosome circulaire superenroulé (à la manière d'un cordon téléphonique). Cet ADN circulaire peut se compacter encore plus en faisant des super-hélices et ceci va donner une structure dite hélicoïdale. En plus du chromosome circulaire principal, certaines bactéries, comme Vibrio cholerae, possèdent parfois une partie de leur génome déportée sur un ou plusieurs mégaplasmides. Enfin, quelques rares bactéries comme les Borrelia ont un chromosome linéaire.
Chez les eucaryotes, l’ADN est présent dans le noyau cellulaire principalement, mais aussi dans les mitochondries et les chloroplastes. Dans le noyau, il est linéaire et est scindé en plusieurs ADN formant des chromosomes. Il est plus ou moins compacté et associé à des protéines comme les histones. Dans les mitochondries et les chloroplastes, l'ADN peut prendre de nombreuses formes différentes, circulaires, linéaires ou encore ramifiés.
Séquence de nucléotides
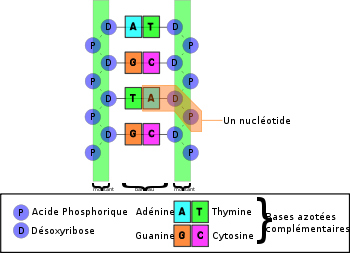 Structure de l'ADN
Structure de l'ADNL'ADN est composé de séquences de nucléotides; on parle de polymère de nucléotides ou encore de polynucléotide. Chaque nucléotide est constitué de trois éléments liés entre eux:
- un groupe phosphate lié à:
- un sucre, le désoxyribose, lui-même lié à :
- une base azotée.
Il existe quatre bases azotées différentes: l'adénine (notée A), la thymine (notée T), la cytosine (notée C) et la guanine (notée G). Chaque base est fixée sur un désoxyribose pour former un nucléoside. Lorsqu'un nucléoside est lié à un ou plusieurs phosphates, on dit qu'il s'agit d'un nucléotide. Dans l'ADN, les nucléotides sont reliés entre eux selon une certaine séquence grâce à des liaisons impliquant un groupe phosphate, qu'on appelle des liaisons 5'-3' phosphodiester. Pour fabriquer un brin d'ADN, il suffit donc d'enchaîner des nucléotides en les reliant par ce type de liaisons, appelées liaisons fortes.
Bases azotées
Ce sont les quatre bases azotées qui assurent la variabilité de la molécule d'ADN, ainsi que la complémentarité des deux brins. En effet, il n'existe que deux types complémentaires de bases : une pyrimidique sera toujours en face d'une purique.
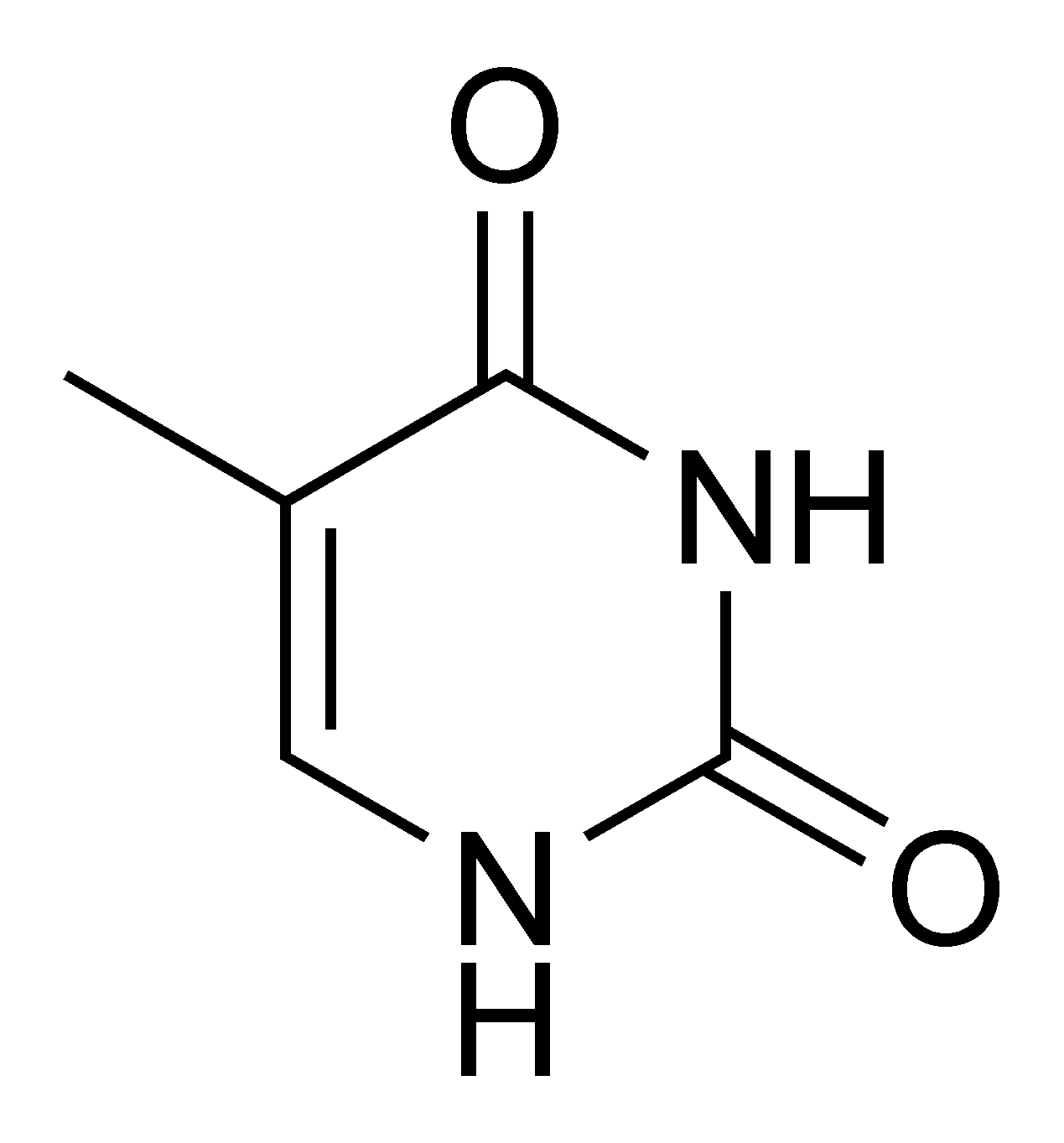
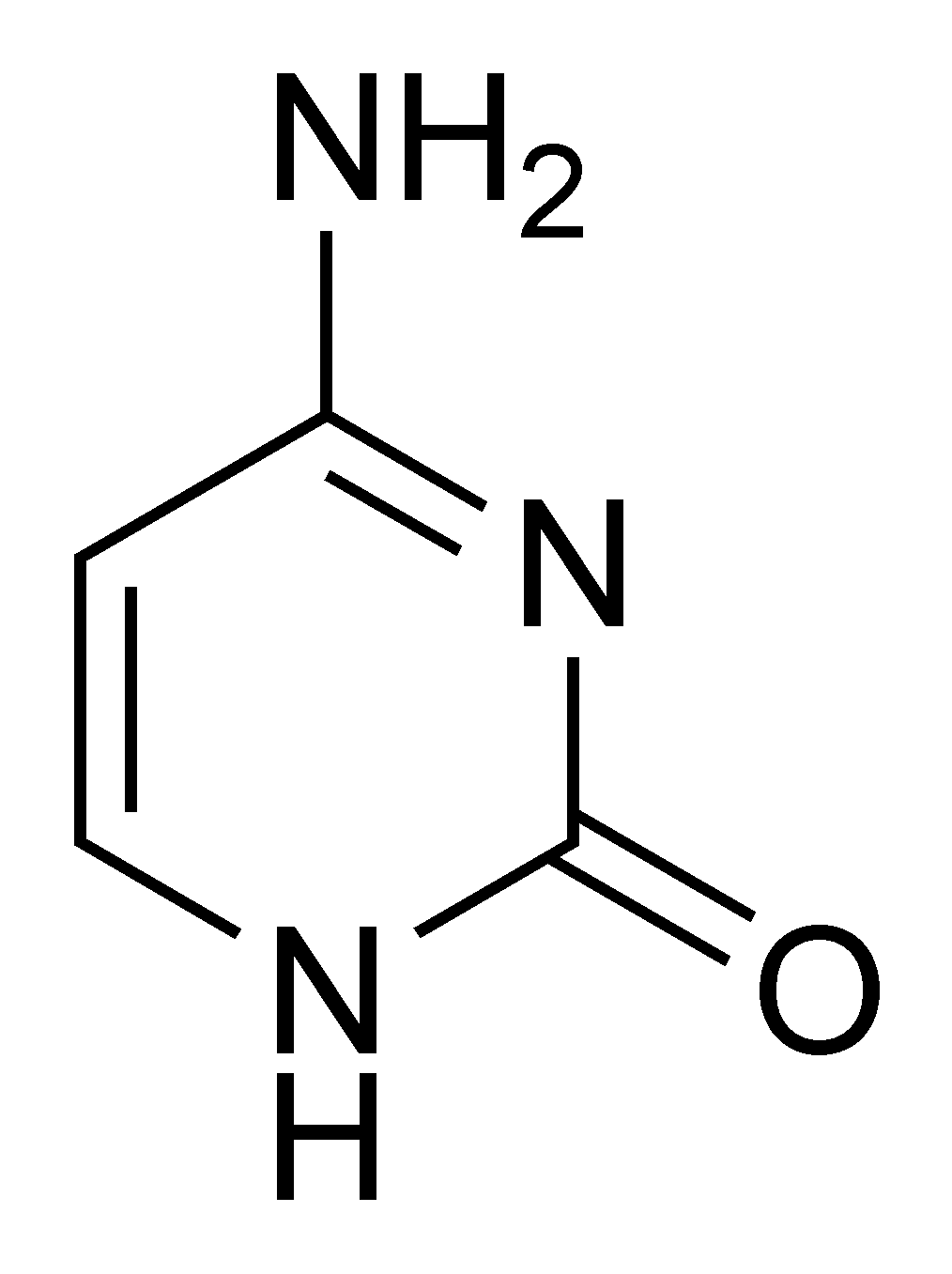
- La thymine[7] (T) et la cytosine[8] (C) sont de la famille des pyrimidiques.
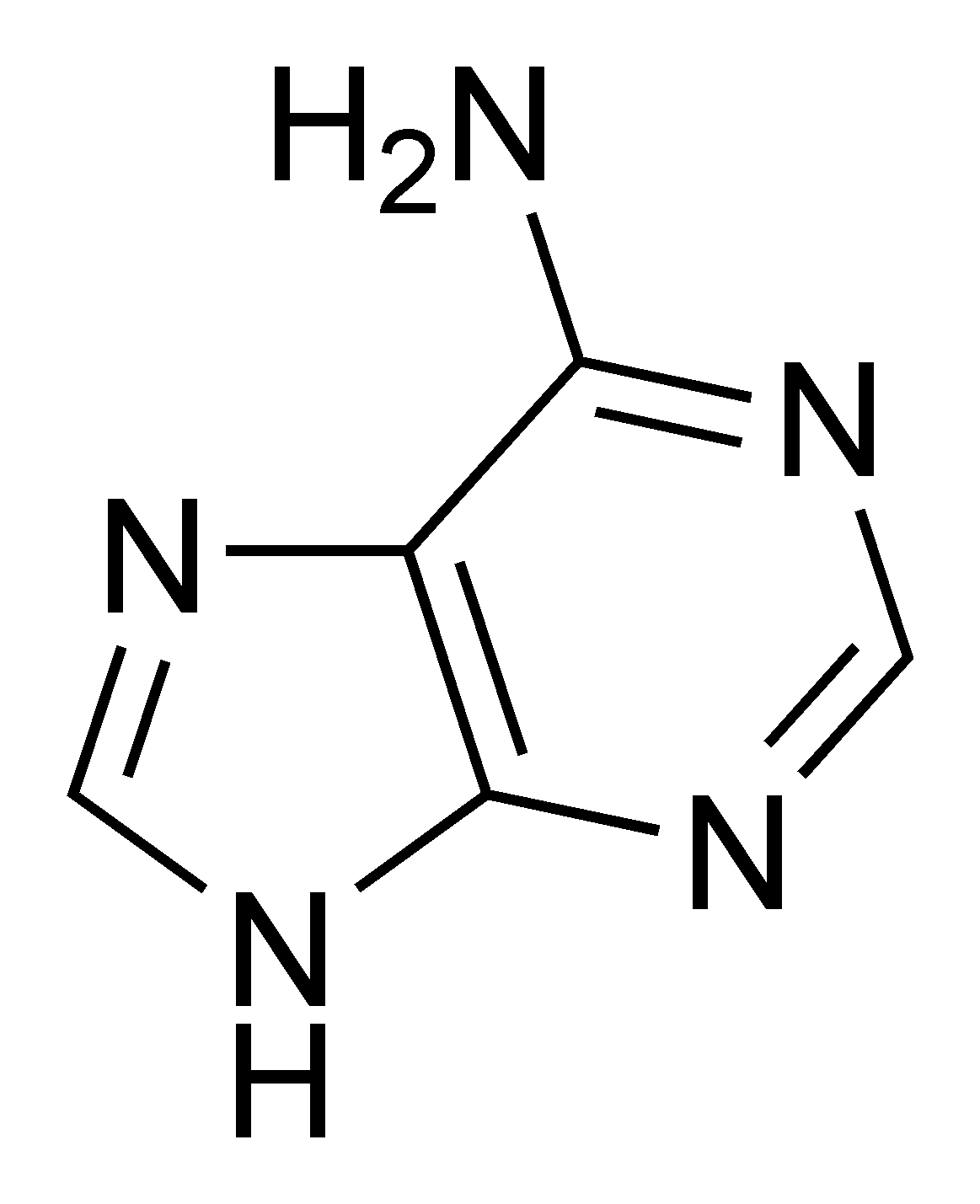
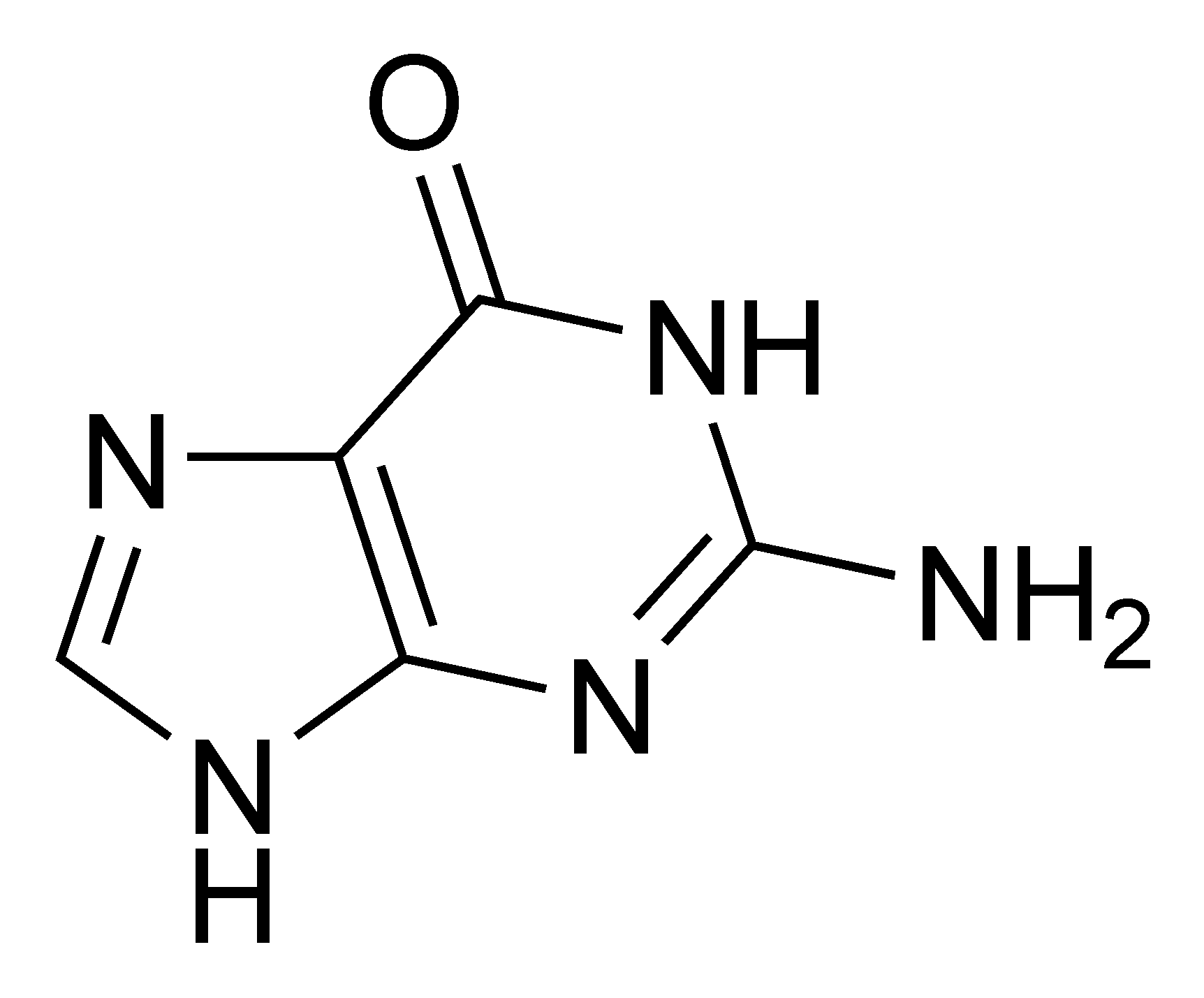
- L'adénine[9] (A) et la guanine[10] (G) sont de la famille des puriques.
Un nucléotide est formé par un groupe de phosphate, du désoxyribose et une base azotée. Par conséquent, il existe quatre nucléotides différents. Un « brin » d'ADN est formé par la répétition ordonnée de ces nucléotides. Les bases azotées sont complémentaires deux à deux, une purique s'associant toujours à une pyrimidique: l'adénine s'associant avec la thymine et la guanine avec la cytosine. Les bases azotées complémentaires sont reliées entre-elles par des liaisons hydrogène.
Complémentarité des deux brins d'ADN
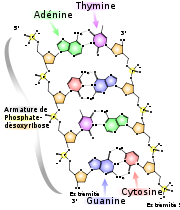 Structure chimique de l'ADN, les liaisons hydrogène sont représentées en lignes pointillées. En haut, la paire AT avec deux liaisons hydrogène, en bas, la paire GC avec trois liaisons hydrogène.
Structure chimique de l'ADN, les liaisons hydrogène sont représentées en lignes pointillées. En haut, la paire AT avec deux liaisons hydrogène, en bas, la paire GC avec trois liaisons hydrogène.L'ADN est composé de deux brins se faisant face, et formant une double hélice. Ceci est possible car les nucléotides trouvés dans un brin possèdent des nucléotides complémentaires avec lesquels ils peuvent interagir par des liaisons hydrogènes (liaisons faibles). Il y a deux liaisons hydrogènes entre A et T et trois entre C et G. En face d'une adénine, il y a toujours une thymine; en face d'une cytosine, il y a toujours une guanine. On a donc les interactions possibles suivantes :
A-T et T-A
G-C et C-G
Pour un brin d'ADN possédant vingt nucléotides comme dans l'exemple suivant, on peut retrouver la séquence du brin complémentaire et reconstituer la double séquence de la double hélice.
5'-ATTGCCGTATGTATTGCGCT-3' 3'-TAACGGCATACATAACGCGA-5'Les deux brins antiparallèles d'ADN sont toujours étroitement reliés entre-eux par des liaisons hydrogène (également appelées « ponts hydrogène » ou encore simplement « liaisons H » ou « ponts H ») formées entre les bases complémentaires A-T et G-C. Ces deux brins d'ADN sont dits complémentaires car les purines (adénine et guanine) d'un brin font toujours face à des pyrimidines de l'autre brin (thymine et cytosine). Les nucléotides sont complémentaires entre-eux. Ainsi, l'adénine est complémentaire à la thymine et la guanine est complémentaire à la cytosine. Deux liaisons hydrogènes retiennent ensemble la paire A-T et trois retiennent la paire G-C.
 Structure 3D de la molécule d'ADN.
Structure 3D de la molécule d'ADN.Les brins d'ADN sont orientés dans le sens 5' vers 3' (et ceci en raison de notations liées à la géométrie du désoxyribose). Deux brins d'une double hélice sont complémentaires et antiparallèles, c'est-à-dire assemblés tête bêche (l'extrémité 5' de l'un est en contact avec l'extrémité 3' de l'autre et inversement). Ce caractère antiparallèle des brins explique l'existence de deux sillons (l'un grand, l'autre petit) autorisant l'accès à la séquence des nucléotides sans avoir à "ouvrir" la molécule en séparant les brins entre eux. Ainsi, une représentation structurale comme celle en haut de page s'avère t-elle inexacte, les deux sillons n'étant pas distincts (deux sillons de même largeur). La représentation animée est donc plus réaliste (deux sillons de largeurs différentes), tout comme celle de la division de l'ADN. Comme une molécule d'ADN est double-brin, on dit qu'elle est bicaténaire.
Grâce à l'alternance des 4 bases azotées A, C, T, G, toutes ces séquences constituent un message codé, portant les informations génétiques. En effet, l'ordre, la nature, et le nombre de nucléotides déterminent l'information génétique. Le lien entre l'information génétique, et les caractères de l'organisme (le phénotype), est gouverné par le code génétique.
La structure de l'ADN permet l'hérédité
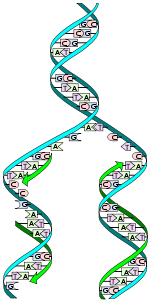 Réplication de l'ADN semi-conservative.
Réplication de l'ADN semi-conservative.Avant chaque division cellulaire, la molécule d'ADN double-brin doit être dupliquée en deux molécules d'ADN filles identiques. Cela assure la transmission de l'information génétique lors de la reproduction, c'est l'hérédité.
Chacune de ces nouvelles molécules hérite d'un brin de la molécule d'ADN initiale ou « mère »; l'autre brin est synthétisé à partir de nucléotides libres. Les nouveaux nucléotides se placent par complémentarité A-T et C-G, de manière à reconstituer à l'identique le brin manquant.
On dit que c'est une réplication semi-conservative. L'hypothèse d'un tel modèle de réplication fut émis par les découvreurs de la structure de l'ADN dès 1953. Quelques années plus tard, les expériences de Meselson et Stahl validèrent ce modèle.
Lors de la réplication, les paires de bases sont tout d'abord désappariées par la rupture des liaisons hydrogènes de l'ADN par une enzyme appelée ADN hélicase. Une fourche de réplication va alors se former donnant 2 brins d'ADN simple-brin distincts. Chacun de ces brins va être copié par l'action des ADN polymérases, pour former 2 nouvelles molécules d'ADN double brins identiques à la molécule initiale.Ce mécanisme de réplication nécessite donc deux brins aux séquences complémentaires, tout deux reliés par des liaisons faibles, pour que la séparation (ou dénaturation) et le réassemblage des brins se fassent facilement.
Article détaillé : Réplication de l'ADN.L'ADN peut subir des modifications
Malgré les liaisons fortes et la complémentarité des bases azotées qui assurent la stabilité de l'information génétique au cours des réplications, la séquence d'un ADN peut se modifier.
- Si la modification se fait sur un ou quelques nucléotides on parle de mutation. Celles-ci sont spontanées, sûrement dues à des erreurs d'appariement au cours de la réplication. Les mutations peuvent être aussi favorisées ou induites par certains agents de l'environnement, appelés facteurs mutagènes (radioactivité, ultra-violet....).
- Les modifications peuvent aussi consister en un échange de portion dans la séquence de nucléotides avec un autre ADN. On parle de recombinaison génétique. Ces recombinaisons génétiques peuvent se faire naturellement (transformation génétique des bactéries, reproduction sexuée), mais aussi artificiellement, par les techniques du génie génétique (cela aboutit alors à des OGM).
Ces processus sont à l'origine des différentes variations des ADN dans le monde vivant. C'est ce qui est à l'origine de la diversité actuelle des êtres vivants c'est-à-dire la biodiversité.
Propriétés physico-chimiques
Fusion ou dénaturation
La température de fusion (ou dénaturation) Tm (melting temperature) des acides nucléiques comme l'ADN est la température pour laquelle 50 % des molécules d'ADN sont désappariées ou dénaturées (i.e. sous forme simple brin). Cette propriété est visible par lecture de l'absorption optique de la solution contenant l'ADN à 260 nm : la densité optique augmente au cours du désappariement (phénomène d'hyperchromicité). L'énergie thermique apportée devient alors suffisante pour rompre les liaisons H interbrins. Cette température dépend donc de la quantité de liaisons hydrogènes présentes. Ce sont d'abord les appariements A-T qui se séparent les premiers au cours de la montée de la température car ils ne possèdent que deux liaisons hydrogènes contrairement aux appariements G-C qui en possèdent trois. Ainsi, lors d'une élévation progressive de la température, il se forme des yeux d'ouvertures dans l'ADN. Plusieurs formules empiriques permettent de calculer la valeur de la température de fusion. Elles tiennent compte du pourcentage de base (G+C), de la salinité du milieu ainsi que de divers facteurs correctifs, tels que la présence de structures secondaires intra ou extra moléculaires (repliement de l'ADN sur lui-même, formation d'appariements entre deux brins). La connaissance de la température de fusion est un élément important au laboratoire lorsqu'il s'agit de faire de la PCR (Réaction en chaîne par polymérase), par exemple.
Un lien hydrogène est une mise en commun d'un proton entre un accepteur et un donneur. Plus il y a de liaisons hydrogènes dans une molécule d'ADN, plus l'énergie de liaison est élevée et plus sa température de fusion sera élevée.
Ainsi une molécule d'ADN double brin composée uniquement d'appariements de C (de G) avec des G (des C) (3 liens H) nécessitera plus d'énergie pour être dénaturée sous la forme de molécules simple-brins, qu'un ADN de même taille composé d'appariements de A (de T) avec des T (des A) (2 liens H). Ceci explique pourquoi la température de fusion de l'ADN varie en fonction de deux facteurs principaux :
- sa taille (exprimée en nombre de bases, généralement en kilobase kb ou mégabase Mb …),
- son rapport (A+T)/(C+G), appelé relation de Chargaff, donnant un indice des proportions de paires A-T versus C-G.
Stabilité
Les enzymes qui hydrolysent les acides nucléiques sont les nucléases. La quasi totalité des cellules possèdent différents types de nucléase dont le but est de faire le ménage pendant le métabolisme des acides nucléiques. Les nucléases sont des phosphodiestérase et elles catalyse l'hydrolyse des liaisons phosphodiester entre les nucléotides. Certaines enzymes clivent spécifiquement l'ADN (des ADNases) ou de l'ARN (ARNases) mais d'autre ne sont pas spécifique et sont appelés tout simplement nucléases.
L'ADN traitée par une solution d'acide chlorhydrique 1M subit une hydrolyse des liaisons glycosidiques au niveau des bases pyrimidiques spécifiquement. L'ADN résiste à l'hydrolyse alcaline. [réf. nécessaire]
Expression de l'information portée par l'ADN
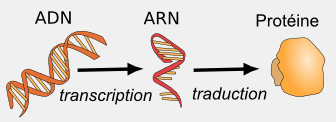
L'information génétique qui constitue le génotype d'un organisme s'exprime pour donner naissance à un phénotype, c'est-à-dire l'ensemble des caractères de cet organisme. Cette expression du génome se fait en interaction avec divers facteurs de l'environnement (nutriments, lumière...). Elle se fait en plusieurs étapes:
- La transcription, qui est le transfert de l'information génétique de l'ADN vers une autre molécule, l'ARN.
- La traduction, qui est un transfert d'information depuis l'ARN vers les protéines.
- L'activité des protéines.
L'activité des protéines détermine l'activité des cellules , qui vont ensuite déterminer le fonctionnement des organes et de l'organisme.
La transcription
Article détaillé : Transcription (biologie).Même, si pour les procaryotes et les eucaryotes, l'ADN ne se trouve pas sous la même forme, il renferme dans les 2 cas l'information génétique, c'est-à-dire que des zones de l'ADN appelé “gènes” codent les protéines. Mais, comment une séquence d'acides nucléiques peut-elle coder une séquence d'acides aminés ? En fait, lorsque la cellule aura besoin de protéines (par exemple, des protéines de structure lors de sa division, ou des enzymes pour fabriquer les molécules dont elle a besoin pour fonctionner), elle va transcrire, c'est-à-dire recopier une partie de ses gènes (c'est-à-dire les gènes codant les protéines d'intérêt) sous forme d'ARN grâce à une enzyme nommée “ARN polymérase ADN dépendante de type II”. Cette enzyme va produire un ARN messager (ARNm) identique à la séquence d'ADN (par exemple : AUGUCUUUAUGU…UAG) du gène. L'existence de l'ARNm a été démontrée par Jacques Monod et ses collaborateurs, ce qui lui valut le prix Nobel de Médecine en 1965. À l'inverse de l'ADN, l'ARNm n'est pas sous forme de double hélice et il adopte des structures secondaires complexes. Il est moins stable que l'ADN, c'est-à-dire qu'il est dégradé plus facilement, de par la présence d'un ribose à la place d'un désoxyribose. Le ribose est très sensible à l'hydrolyse alcaline tandis que le désoxyribose y est totalement insensible.
La transcription est un processus complexe et l'élucidation de ses mécanismes fut l'une des grandes avancées de la biologie de la seconde moitié du XXe siècle. C'est un processus hautement régulé, notamment grâce à des protéines appelées facteurs de transcription qui, en réponse à des hormones par exemple, vont permettre la transcription de gènes cibles (par exemple les gènes exprimés quand la cellule reçoit des œstrogènes, ou de la progestérone, des hormones dites sexuelles). Une dérégulation des mécanismes de contrôle et la machinerie s'emballe, les ARN sont transcrits de manière désordonnée, les protéines sont présentes en excès, entraînant un fonctionnement aberrant des cellules, un fonctionnement cancéreux. En effet, dans un grand nombre de cancers, la transcription de certains gènes est altérée, ce qui entraîne un dérèglement total de la cellule qui se divise activement et de façon désordonnée.
La traduction
Cet ARNm sera traduit en protéine par des ribosomes. Ces ribosomes vont décoder l'ARNm, c'est-à-dire le code AUG UCU CUU … pour assembler les acides aminés correspondants et faire une protéine. Le ribosome est un complexe comprenant des ARN ribosomiaux (ARNr) et des protéines. Chez les eucaryotes, les ARNm sont d'abord maturés avant d'être traduits, grâce à des ARNsn (snRNA en anglais, petits ARN nucléaires)…
Le code génétique
Article détaillé : Code génétique.Le code génétique est le système de correspondance entre les séquences de nucléotides de l'ADN et les séquences en acides aminés des protéines.
L'enchaînement des quatre nucléotides A, C, T, G, dans une séquence génique doit coder l'enchaînement des 20 acides aminés au niveau de la protéine. Si une base codait un seul acide aminé, seuls 4 acides aminés pourraient être codés de façon non ambiguë. Le codage d'un acide aminé nécessite donc au minimum une suite de 3 bases (64 possibilité d'arrangement, ou codons). Il est possible donc de coder 61 acides aminés différents et 3 codons d'arrêt de la traduction: UAA UAG ET UGA. Le code est dit dégénéré (on parle de redondance du code génétique), un acide aminé peut être codé par plusieurs codons pour cette raison.
Variations possibles de la structure spatiale de l'ADN
- ADN bombé:l'ADN qui bouge a des structures dynamiques et fait des mouvements.
les structures d'ADN vus in vivo par ailleurs possèdent un role fonctionnel: la recombinaison génétique et la mutation.
- ADN Z: une double hélice lévogyre dont le squelette présente une conformation de structure zigzag mai plus lisse que l'ADN B.
il y a un seul sillon qui ressemble au grand sillon de l'ADN B. les paires de bases qui forment dans l'ADN B le grand sillon proche de l'axe sont rejetés à l'exterieur au niveau de l'ADN Z. les phosphores sont plus proches les uns des autres. l'ADN Z ne peut pas former le nucléosome.une proportion est formée de bases G-C favorise la conformation Z et la méthylation de la cytosine.
- ADN fusiforme et ADN à épingle à cheveux:
les jonctions de Holiday formées lors de la recombinaison sont des structures cruciformes de répétitions inversées en miroir de segment ADN polypurines, polypyrimidiques est également produit des structures cruciformes ou épingle à cheveux par appariment intrabrin.
- ADN H ou ADN triplex: des répétitions inversées (polychrome) des segments d'ADN polyurine, polypyrimidine peuvent former des structures triplex. on obtient alors ADN triple brin + un simple brin.
l'ADN H pourrait avoir un role dans la réguation fonctionnelle de l'expression des gènes, ainsi que sur les ARN. par exemple: repression de la transcription.
- ADN G: ADN quadruplex, repliment des séquences double brin riches en G et C sur elle-même formant des appariments de bases de type Hoogsteen entre 4 guanines et la structure particulièrement stable et souvent près des promoteurs des gènes et au niveau des télomères.
Différentes formes de l'ADN
Comme expliqué précédemment, deux molécules d'ADN sont appariées via les liaisons hydrogènes entre leurs bases azotées pour former la double-hélice d'ADN (ADN sous forme double-brin). C'est sous cette forme stable que l'ADN est présent dans les organismes vivants. Pourtant cette double-hélice peut être ouverte afin de permettre l'exécution de processus biologiques fondamentaux (tels la réplication ou la transcription) générant ainsi de l'ADN sous forme simple brin. Suivant les conditions du milieu, ces deux formes d'ADN (simple et double brin) peuvent voir leur structure varier. Ces structures sont dans l'ensemble rares, et leur fonctions biologiques (si elles en ont) mal connues.
Plusieurs types d'ADN double-brin
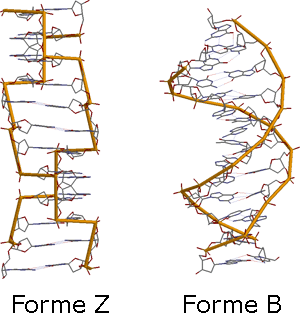 Formes ou types Z et B
Formes ou types Z et BSelon la composition du milieu extérieur, en particulier le pourcentage d'eau lié aux phosphates hydrophiles, la double-hélice d'ADN peut adopter trois structures:
- 95 % d'eau : type B
- 70 % d'eau : type A
- 50 % d'eau : type Z
Ces structures existent aussi in-vivo :
- ADN-B : forme d'ADN la plus commune. Elle a une hélice à pas dextrogyre, des plateaux de base perpendiculaires à l'axe de l'hélice passant au centre de l'appariement de ces dernières. Elle possède 10,5 paires de bases par tour (soit 21 nucléotides) soit une rotation de 36° entre chaque sucre (ou 34A). Les sucres sont en position anti (noyau des bases à l'extérieur des sucres), endo et radiale par rapport aux bases. L'espace vertical entre chaque paire de base est de 0,34 nm .
- ADN-A : forme d'ADN spécifique à la transcription. En effet l'ARN étant de type A, lors de la transcription, l'ARN stimule un transfert de l'ADN du type B vers A. À la fin de la transcription, lorsque l'ARN s'est détaché, l'ADN reprend sa conformation B.
Le type A est caractérisé par des plateaux de base très incliné, une position tangentielle des sucres (ainsi que anti et endo), un axe passant dans le grand sillon et non plus par le milieu d'appariement des bases, et 11 paires de bases par tour soit 32,7° entre chaque sucre.
- ADN-Z : son rôle est de favoriser l'interaction des bases avec les protéines régulatrices. C'est une hélice à pas lévogyre. Il est caractérisé par des plateaux peu inclinés (9° à peu près), et une position alternative des sucres en radiale et tangentielle (ainsi qu'une alternance 3'endo syn/5'endo anti). L'axe passe par le petit sillon et présente 12 paires de bases par tour soit 30° entre chaque sucre. Le passage de l'ADN-B en ADN-Z est favorisée par la présence de multiples cytosines au sein des promoteurs.
Autres structures de l'ADN
- Les triplex: structure formée lorsque une molécule d'ADN simple brin vient s'apparier dans le grand sillon d'une double hélice d'ADN
- Les G-quadruplexes: Structure secondaire formée par de l'ADN simple brin lorsqu'il est riche en guanine, formant un empilement de plateaux ("quartets") constitués chacun de 4 guanines.
- Les hairpines
Propriétés mécaniques
Les propriétés mécaniques de l'ADN peuvent être étudiées par des simulations numériques de dynamique moléculaire ainsi que par des expériences de manipulation de molécules uniques (par exemple, à l'aide de pincettes optiques ou magnétiques). Comme tous les polymères, l'ADN est une molécule élastique. Sous des contraintes faibles, un double brin peut être décrit par des modèles standards de la physique des polymères (modèle du ver, etc.). Cependant, en appliquant une force de 65pN aux extrémités d'un double brin, l'on fait transiter celui-ci vers une nouvelle forme, environ 1.7 fois plus longue, dite ADN-S (stretched). Ceci peut s'interpréter par une rotation des paires de base : la double hélice se transforme en "échelle", ou en "fibre". Il semblerait que cette transition joue un rôle dans certains processus biologiques, telles que la réparation de l'ADN par certaines protéines
Différents types d'enzymes liées à l'ADN
- ADN hélicase : enzyme qui catalyse le déroulement des brins complémentaires d’une double hélice d’ADN.
- ADN ligase : enzyme catalysant la liaison entre deux molécules séparées d’ADN, formant des liaisons phosphodiesters entre l’extrémité 3'-hydroxyl de l’une et l’extrémité 5'-phosphate de l’autre. Son rôle naturel réside dans la réparation et la réplication de l’ADN. C'est un outil essentiel dans la technologie de l’ADN recombinant puisqu'elle permet l’incorporation d’ADN étranger dans les vecteurs.
- ADN polymérase : enzyme catalysant la polymérisation (5' vers 3') des monodésoxynucléotides triphosphates qui constituent l'ADN. En absence de monodésoxynucléotides triphosphates, elle a un rôle d' exonucléase, supprimant les nucléotides non-appareillées des brins "sticky" ( sens 3' vers 5')
- ADN primase : enzyme qui catalyse la synthèse de courtes amorces d’ARN à partir desquelles débute la synthèse des brins d’ADN.
- ADN topo-isomérase ou topoïsomérase (ex. ADN gyrase) : enzyme qui catalyse l’introduction ou l’enlèvement des surenroulements dans l’ADN.
ADN et art
Article détaillé : Influence de l'acide désoxyribonucléique dans la culture.La structure hélicoïdale a inspiré un certain nombre d'artistes. Le plus célèbre reste le peintre surréaliste Salvador Dali qui s'en inspire dans neuf tableaux entre 1956 et 1976 dont Le Grand masturbateur dans un paysage surréaliste avec ADN et Galacidalacidesoxyribonucleicacid [11]
Notes et références
- ↑ ou, surtout dans les plus vieux ouvrages, DNA de l'anglais deoxyribonucleic acid
- ↑ Le premier âge de l'ADN, éditions Vuibert
- ↑ (en) O.T. Avery, C.M. McLeod et M. McCarthy, « Induction of transformation by a desoxyribonucleic acid fraction isolated from Pneumococcus Type III », dans J. Exp. Med., vol. 79, 1944, p. 137-158
- ↑ publication originale (en)[pdf]
- ↑ inventée par William Lawrence Bragg 40 ans plus tôt
- ↑ Par exemple le chromosome 1 humain mesure (245 millions)X(3.4.10-10) mètre = 0.083 mètre, soit plus de 8 cm.
- ↑ 5-méthyluracile (ajout d'un méthyle sur un uracile) — 2,4 dihydroxy-5méthylpyrimidine
- ↑ ou 2 hydroxy, 4-aminopyrimidine (fonction amine en position 4).
- ↑ ou 6-aminopurine
- ↑ ou 2-amino, 6-hydroxypurine
- ↑ Acide Dalioxyribonucléique, M Morange, Pour la science, sept 2006, p 96-97
- Arrêté de terminologie du 14 septembre 1990.
Voir aussi
Liens internes
- Une liste des différents types d'ADN (ADN-T, ADN chloroplastique, ADNc, etc.)
- Une liste des différents types d'ARN
- ARN
- ADN mitochondrial
- Analyse de l'ADN
- Empreinte génétique (« test ADN »)
- Extraction d'ADN
- Génome
- Ordinateur à ADN
- Paire de bases
- Réparation de l'ADN
- Liste de sigles de biologie moléculaire
Liens externes
- Les empreintes génétiques et l'identification judiciaire
- (en) Le journal Nature fête les 50 ans de l'ADN
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire Portail de la biochimie
Portail de la biochimie
Catégorie : ADN








Wikimedia Foundation. 2010.