- Tyrannosaurus
-
 Pour les articles homonymes, voir Tyrannosaurus rex (homonymie).
Pour les articles homonymes, voir Tyrannosaurus rex (homonymie). Tyrannosaurus
Tyrannosaurus
Le Tyrannosaurus rex "Sue" Classification Règne Animalia Classe Sauropsida Ordre Saurischia Sous-ordre Theropoda Super-famille Coelurosauria Famille Tyrannosauridae Genre Tyrannosaurus
Osborn, 1905Espèces de rang inférieur Synonymes - Manospondylus gigas Cope, 1892
- Dynamosaurus imperiosus Osborn, 1905
- Nanotyrannus lancensis? Gilmore, 1946
- Aublysodon molnari Paul, 1988
- Dinotyrannus megagracilis Olshevsky, 1995
- Stygivenator molnari (Paul, 1988) Olshevsky, 1995
Tyrannosaurus, ou tyrannosaure, est un genre de dinosaure théropode appartenant au clade des Tyrannosauridae et ayant vécu à l'extrême fin du Crétacé (Campanien et Maastrichtien, il y a environ 70 à 65 millions d'années) dans ce qui est actuellement l'Amérique du Nord. Tyrannosaurus rex, souvent abrégé en « T. rex » et dont l'étymologie du nom signifie « roi des lézards tyrans », est l'une des plus célèbres espèces de dinosaure et l'unique espèce de Tyrannosaurus si le taxon Tarbosaurus bataar n'est pas considéré comme une espèce de Tyrannosaurus. Ce dernier fut l'un des derniers dinosaures non-aviens à avoir vécu jusqu'à l'extinction survenue à la limite Crétacé-Paléocène il y a 65 millions d'années.
Tout comme les autres membres du clade des Tyrannosauridae, Tyrannosaurus fut un carnassier bipède doté d'un crâne massif équilibré par une longue queue puissante. Comparés à ses larges membres postérieurs, les bras du Tyrannosaurus étaient petits et atrophiés et ne portaient que deux doigts griffus. Bien que d'autres théropodes rivalisaient voire dépassaient Tyrannosaurus en taille, il fut le plus grand Tyrannosauridae connu et l'un des plus grands carnivores terrestres ayant existé sur la planète, mesurant jusqu'à près de 12 mètres de longueur[1], 4 mètres à hauteur de hanches[2] et pesant jusqu'à 6,7 tonnes[3]. De loin le plus grand des carnivores de son temps, le T. rex a pu être un superprédateur au sommet de la chaîne alimentaire, chassant notamment des herbivores de grande taille tels que les Hadrosauridae et les Ceratopsidae, même si certains experts suggèrent qu'il était avant tout charognard.
Plus de 30 spécimens fossiles de Tyrannosaurus rex ont été recensés, certains étant presque complets. Des restes de tissus mous et de protéines fossilisés ont été retrouvés sur au moins l'un de ces spécimens[4]. Cette abondance de matériaux a permis de nombreuses avancées dans bien des aspects de l'histoire et de la biologie de cette espèce. Si certains points sont consensuels, d'autres restent controversés, tels que ses habitudes alimentaires, sa physiologie ou sa vitesse de pointe. Même sa place taxinomique est sujette à débat, certains considérant T. rex comme la seule espèce de la famille des Tyrannosauridae alors que d'autres considèrent le Tarbosaurus bataar d'Asie, sensiblement identique au niveau anatomique, comme la seconde espèce de cette famille. Plusieurs autres genres de la famille des tyrannosauridés ont également été synonymisés avec Tyrannosaurus.
Sommaire
Étymologie
Tyrannosaurus signifie « lézard tyran », nom proposé en 1905 par Henry Fairfield Osborn, créateur du département de paléontologie des vertébrés à l’American Museum of Natural History de New York. Dérivé des mots grecs τύραννος (Tyrannos, « tyran ») et σαῦρος (Sauros, « lézard ») ainsi que du terme latin rex qui signifie roi. Osborn lui attribua cette appellation car ce fut un prédateur impressionnant, avec des griffes et des dents gigantesques. Le nom binominal complet, Tyrannosaurus rex, peut être traduit par « roi des lézards tyrans », soulignant la dominance imaginée de l'animal sur les autres espèces de son temps[5].
On l'appelle souvent « T. rex ». Cependant, on trouve souvent le diminutif abusif « T-Rex ». Il est faux car il n'y a pas de trait d'union dans « Tyrannosaurus rex ». Le point dit que le « T » est l'abréviation de « Tyrannosaurus » alors que le trait d'union n'a jamais ce rôle.
Son ancien nom proposé par Barnum Brown lors de sa découverte, Dynamosaurus imperiosus (Saurien dynamique impérial), sera vite oublié[6].
Description
 Comparaison de taille entre l'un des plus grands spécimens de Tyrannosaurus rex retrouvés et un être humain.
Comparaison de taille entre l'un des plus grands spécimens de Tyrannosaurus rex retrouvés et un être humain.
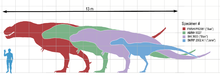 Comparaison de taille entre différents spécimens de T. rex et un humain.
Comparaison de taille entre différents spécimens de T. rex et un humain. Comparaison de taille entre différents dinosaures théropodes géants, le Tyrannosaurus rex est en violet.
Comparaison de taille entre différents dinosaures théropodes géants, le Tyrannosaurus rex est en violet.Le Tyrannosaurus rex est l'un des plus grands carnivores terrestres de tous les temps. Le plus grand spécimen découvert à ce jour, répertorié sous le code FMNH PR2081 et surnommé « Sue », du nom de la paléontologue Sue Hendrickson, mesure 11,2 mètres de long et 4 mètres de haut, à hauteur de hanche[2]. Les différentes estimations de la masse du Tyrannosaurus rex ont grandement varié au cours des années, allant selon les auteurs de plus de 7,2 tonnes[7] à moins de 4,5 tonnes[8],[9] avec d'après les estimations les plus récentes une fourchette allant de 5,4 à 6,7 tonnes[3],[10],[11],[12].
Si le Tyrannosaurus rex était plus grand que l'Allosaurus, un autre théropode bien connu du Jurassique, il était légèrement moins imposant que le Spinosaurus et le Giganotosaurus, deux carnivores du Crétacé[13],[14].
Comme chez les autres théropodes, le cou du T. rex forme une courbe en forme de « S » afin de maintenir la tête au-dessus du corps, mais il est particulièrement court et musculeux afin de supporter la tête massive. Les bras sont courts et se terminent par deux doigts. En 2007, un spécimen possédant trois doigts à chaque main a été découvert dans la formation de Hell Creek dans le Montana, suggérant la possible présence d'un troisième doigt vestigial chez Tyrannosaurus[15], hypothèse restant à confirmer[16]. Proportionnellement à la taille du corps, les jambes du T. rex sont parmi les plus longues de tous les théropodes. La queue est longue et massive, constituée parfois de plus de quarante vertèbres, agissant comme un balancier permettant d'équilibrer la lourde tête et le torse. Afin d'alléger l'animal et de lui permettre de se mouvoir suffisamment rapidement, de nombreux os sont creux, réduisant la masse sans perte significative de solidité[1].
Le plus grand crâne de T. rex mesure 1,535 mètre (5 pieds) de longueur[17]. De larges cavités aériennes permettaient de réduire la masse du crâne, et laissaient la place aux attaches des muscles de la mâchoire, comme chez tous les théropodes carnivores[18]. Mais, sur d'autres aspects, le crâne de Tyrannosaurus est significativement différent de celui des autres grands théropodes. Extrêmement large à l'arrière et muni d'un museau étroit, il permet un très bonne vision stéréoscopique[19],[20].
Les os du crâne sont massifs et certains os de la face dont l'os nasal sont fusionnés, empêchant tout mouvement. Beaucoup sont « pneumatisés » (constitués d'une structure en nid d'abeilles de petites poches d'air) ce qui aurait pour conséquence de les rendre plus souples et plus légers. Ces caractéristiques du crâne des tyrannosauridés leur aurait donné une morsure très puissante dépassant largement celle de tous les non-tyrannosauridés[21],[22],[23].
L'extrémité de la mâchoire supérieure est en forme de « U » (alors qu'elle est en forme de « V » chez la plupart des carnivores non-tyrannosauroidés), ce qui augmente la quantité de chair et d'os pouvant être arrachée à chaque bouchée, tout en augmentant l'effort exercé sur les dents frontales[24],[25]. L'étude des dents de Tyrannosaurus rex montrent une importante hétérodontie, c'est-à-dire la présence de dents de morphologie différente[1],[26].
Les prémaxillaires à l'avant de la mâchoire supérieure sont resserrés, avec des crêtes de renforcement sur la surface arrière, en forme d'incisive recourbée vers l'arrière, de façon à réduire le risque que les dents ne s'arrachent quand le Tyrannosaurus mordait et tirait. Les autres dents sont robustes, plus largement espacées et également renforcées par des crêtes[27]. Les dents de la mâchoire supérieure sont plus grandes que toutes les autres. La plus grande trouvée à ce jour est estimée à 30 cm de long, racine comprise, ce qui en fait la plus grande dent de dinosaure carnivore[2].
Dans les premiers temps, les paléontologues pensaient qu'il se tenait presque verticalement à cause de sa bipédie. Mais suite à la découverte de nouveaux squelettes et à des études biomécaniques, il s'avère qu'il se serait tenu à l'horizontale car c'est la seule manière pour que ses vertèbres supportent son poids. Le tyrannosaure ne devait donc pas dépasser 6 mètres.
Il se tenait sur ses deux pattes arrières. Ses membres postérieurs, terminés par un pied à trois orteils griffus, étaient particulièrement puissants. Sa vision frontale lui permettait d'évaluer efficacement les distances. Afin de pouvoir soutenir son immense tête, ses membres antérieurs étaient atrophiés (« miniaturisés »). Ses bras avaient néanmoins des muscles développés et ils disposaient de deux doigts avec des griffes acérées. Ils servaient sans doute à maintenir la nourriture, mais étaient trop courts (comparables à ceux d'un homme) pour pouvoir la ramasser au sol. Le tyrannosaure était donc obligé de se pencher pour ronger les carcasses de ses proies. Certaines de ses dents, particulièrement impressionnantes (atteignant 18 cm de long), étaient crénelées comme des couteaux à viande. On suppose qu'il pouvait déplacer l'un de ses maxillaires vers l'arrière. D'autre part, l'usure des dents fossilisées indique qu'il mâchait des aliments relativement durs. La mâchoire du tyrannosaure était d'une puissance phénoménale. Elle est considérée comme la plus puissante de tout le règne animal vivant ou éteint : il était ainsi capable d'arracher des kilos de chair d'un seul coup.
Certains scientifiques pensent que sa morsure était empoisonnée en raison des bouts de chair devant rester coincés entre les dents et se décomposant là.
Il n'est pas exclu que le tyrannosaure, comme d'autres reptiles de cette époque, ait été pourvu de plumes. Une équipe de chercheurs a d'ailleurs récemment découvert dans un fémur brisé des tissus mous » (phénomène extrêmement rare). « Les vaisseaux (sanguins) et leur contenu sont similaires à ceux observés dans les os d'autruche », dit la paléontologue Mary Schweitzer. Un élément qui vient à nouveau appuyer l'hypothèse selon laquelle les oiseaux seraient cousins, sinon les descendants des dinosaures. Cette découverte de « tissus mous » offre de nombreux espoirs pour de prochaines études.
Systématique
Tyrannosaurus appartient à la superfamille des Tyrannosauroidea, à la famille des Tyrannosauridae, au sous-ordre des Theropoda et à l'ordre des saurischiens. La famille des tyrannosauridés inclut également le Daspletosaurus, dinosaure de l'Amérique du Nord et l'asiatique Tarbosaurus[28],[29], tout deux ayant parfois été considérés comme synonymes de Tyrannosaurus[25]. On a considéré autrefois les tyrannosauridés comme étant des descendants des premiers grands théropodes comme les mégalosaures et les carnosaures, mais ils ont été reclassés plus récemment parmi les cœlurosauriens[24].
En 1955, le paléontologue soviétique Evgeny Maleev découvre en Mongolie une nouvelle espèce qu'il va nommer Tyrannosaurus bataar[30]. Mais en 1965, les spécialistes découvrent que cette espèce se distingue du Tyrannosaurus nord-américain, et elle est alors renommée Tarbosaurus bataar[31]. De nombreuses analyses phylogénétiques ont permis de découvrir que le Tarbosaurus bataar était un taxon frère du Tyrannosaurus rex[29] et il a été souvent considéré comme étant une espèce asiatique du Tyrannosaurus[24],[32],[33]. Une nouvelle description du crâne du Tarbosaurus bataar a permis de démontrer qu'il est plus étroit que celui du Tyrannosaurus rex et que la répartition de la pression lors d'une morsure devait être bien différente, plus proche de celle de l'Alioramus, un autre tyrannosaure asiatique[34]. Une analyse cladistique a révélé que c'était Alioramus, et non pas Tyrannosaurus, qui était le taxon frère de Tarbosaurus, ce qui suggérerait que Tarbosaurus et Tyrannosaurus devraient bien rester distincts[28].
D'autres fossiles de tyrannosauridés trouvés dans les mêmes formations que Tyrannosaurus rex ont été initialement classés comme des taxons distincts, y compris Aublysodon et Albertosaurus megagracilis[25], ce dernier étant renommé Megagracilis dinotyrannus en 1995[35]. Cependant, ces fossiles sont désormais universellement considérés comme étant des formes juvéniles de Tyrannosaurus rex[36].
Un petit crâne presque complet trouvé dans le Montana, mesurant 60 cm de long, pourrait être une exception. Ce crâne a été classé initialement comme une espèce de Gorgosaurus (G. lancensis) par Charles Whitney Gilmore en 1946[37] mais il a été ensuite rattaché à un nouveau genre, Nanotyrannus[38]. Les avis restent partagés sur la validité de N. Lancensis. De nombreux paléontologues considèrent qu'il appartient à un juvénile de Tyrannosaurus rex[39]. Il existe des différences mineures entre les deux espèces, notamment un plus grand nombre de dents chez N. lancensis, ce qui conduit certains à recommander de conserver les deux genres distincts jusqu'à ce que de nouvelles découvertes permettent de clarifier la situation[29],[40]. Depuis, un crane avec une taille et un nombre de dents intermédiaires corrobore la thèse d'une seule espèce[41],[42].
Manospondylus
 Crâne du spécimen type au Carnegie Museum of Natural History. Restauré de façon erronée, avec pour modèle un Allosaurus[réf. nécessaire], il a depuis été démonté[réf. nécessaire].
Crâne du spécimen type au Carnegie Museum of Natural History. Restauré de façon erronée, avec pour modèle un Allosaurus[réf. nécessaire], il a depuis été démonté[réf. nécessaire].Le premier fossile à avoir pu être attribué à Tyrannosaurus rex consiste en deux morceaux de vertèbres (dont l'un a été perdu) découvert par Edward Drinker Cope en 1892 et décrit comme Manospondylus gigas. Osborn reconnait la similitude entre M. gigas et Tyrannosaurus rex dès 1917 mais, en raison de la nature fragmentaire des vertèbres, il ne peut pas conclure qu'il s'agit de synonymes[43].
En juin 2000, le Black Hills Institute localise M. gigas dans le Dakota du Sud où il déterre des os appartenant au même individu, identiques à ceux de Tyrannosaurus rex. Selon les règles du Code international de nomenclature zoologique (CINZ), l'appellation originale, Manospondylus gigas , devrait avoir la priorité sur Tyrannosaurus rex. Toutefois, la quatrième édition du CINZ, datant du 1er janvier 2000, stipule que « l'usage en vigueur doit être maintenu » quand « le synonyme ou l'homonyme le plus ancien n'a pas été utilisé après 1899 » et « le synonyme ou l'homonyme le plus récent a été utilisé pour un taxon particulier dans au moins 25 ouvrages, publiés par au moins 10 auteurs dans les 50 dernières années »[44]. Tyrannosaurus rex peut être considéré comme étant le nom valide en vertu de ces conditions et serait probablement considéré comme nomen protectum (« dénomination protégée ») alors que Manospondylus gigas serait considérée nomen oblitum (« dénomination oubliée »)[45].
Découverte
 Modèle de la première reconstitution d'un Tyrannosaurus rex réalisée par H.F. Osborn pour une exposition de l'American Museum of Natural History.
Modèle de la première reconstitution d'un Tyrannosaurus rex réalisée par H.F. Osborn pour une exposition de l'American Museum of Natural History.En 1874, A. Lakes découvre près de Golden, dans le Colorado des dents ayant appartenu au Tyrannosaurus. Dans les années 1890, J. B. Hatcher rassemble des éléments post-crâniens à l'est du Wyoming. À l'époque, les paléontologues pensaient avoir trouvé des fossiles d'une espèce de grand Ornithomimus (O. grandis), mais ils appartenaient en réalité au Tyrannosaurus rex. Les fragments de vertèbres découverts dans le Dakota du Sud par E. D. Cope en 1892 et nommés Manospondylus gigas ont également été reclassés Tyrannosaurus rex[15].
Les premiers restes significatifs furent découverts en 1902 et l'animal fut décrit et baptisé par Henry Fairfield Osborn en 1905. Des découvertes de squelettes entiers, en 1988 (au Montana) et 1990 (Dakota du Sud), ont fait considérablement évoluer notre connaissance du tyrannosaure. En 2006, on a découvert, dans un fémur exhumé dans le Montana, des tissus mous appelés « os médullaire » qui n'existent aujourd'hui que chez les oiseaux femelles. Après déminéralisation, les paléontologues y ont retrouvé des vaisseaux sanguins ayant conservé leur élasticité.
Barnum Brown, conservateur adjoint de l'American Museum of Natural History, trouva le premier squelette partiel de Tyrannosaurus rex dans l'Est du Wyoming en 1900. H.F. Osborn appela ce squelette Dynamosaurus imperiosus dans un article en 1905. Brown trouva un autre squelette partiel dans la formation de Hell Creek au Montana en 1902. Osborn utilisa cet holotype pour décrire Tyrannosaurus rex et D. imperiosus dans le même article[5]. En 1906, Osborn reconnait les deux comme étant des synonymes, et choisi Tyrannosaurus comme étant le nom valide[46]. Les éléments d'origine de Dynamosaurus se trouvent dans les collections du Natural History Museum de Londres[47].
Au total, Brown trouva cinq squelettes partiels de Tyrannosaurus. Le spécimen de 1902 fut vendu au Musée d'histoire naturelle Carnegie de Pittsburgh en Pennsylvanie en 1941. La quatrième trouvaille de Brown, la plus complète, fut également retrouvée à Hell Creek et se trouve au Musée américain d'histoire naturelle à New York[48].
Bien qu'il existe de nombreux squelettes de Tyrannosaurus de par le monde, une seule trace de pas a été documentée - celle du Philmont Scout Ranch, dans le Nord du Nouveau-Mexique. Elle a été découverte en 1983 et identifiée et documentée en 1994[49].
Spécimens notables
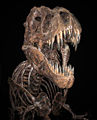
Le 12 août 1990 Sue Hendrickson, paléontologue amateur américaine, découvre le plus complet (environ 85 %) et, jusqu'en 2001, le plus grand, squelette fossile de Tyrannosaurus connu dans la formation de Hell Creek près de Faith en Dakota du Sud. Ce Tyrannosaurus, surnommé « Sue » en son honneur, a fait l'objet d'une bataille juridique concernant son propriétaire. En 1997, c'est Maurice Williams, le propriétaire du terrain d'origine, qui a été déclaré propriétaire du fossile, acheté par le Field Museum aux enchères pour 7,6 millions de dollars américains, ce qui en fait le squelette de dinosaure le plus cher à ce jour. De 1998 à 1999, plus de 25 000 heures de travail ont été nécessaires pour nettoyer le squelette[50]. Les os ont ensuite été expédiés au New Jersey où le montage a été réalisé. Une fois achevé, le squelette a été démonté, les os numérotés ont été renvoyés sur Chicago pour l'assemblage final. Le squelette a été montré au public le 17 mai 2000 dans la grande salle du musée Field. Une étude des os fossilisés a montré que « Sue » avait atteint sa taille adulte à 19 ans et était morte à 28 ans, ce qui en fait le tyrannosaure le plus âgé jamais découvert[51]. L'hypothèse initiale voulant que Sue soit morte d'une morsure à l'arrière de la tête n'a pas été confirmée. Bien que l'étude du squelette ait retrouvé de nombreuses pathologies osseuses, aucune trace de morsures n'a été retrouvée[52]. Les dégâts de l'arrière de son crâne pourraient avoir été causé par un piétinement post mortem. Des spéculations récentes indiquent que Sue pourrait être morte après avoir contracté une infection parasitaire consécutive à l'ingestion de viande contaminée ; l'infection aurait provoqué une inflammation de la gorge et une impossibilité à s'alimenter, menant finalement Sue à mourir de faim. Cette hypothèse est étayée par la présence de trous dans son crâne, semblables à ceux causés par un parasite similaire, un trichomonas touchant les oiseaux d'aujourd'hui[53],[54],[55].
Un autre Tyrannosaurus, surnommé « Stan », en honneur du paléontologue amateur Stan Sacrison, fut trouvé dans la formation de Hell Creek près de Buffalo en Dakota du Sud, au printemps 1987. Après 30 000 heures de travail de préparation et de fouille, un squelette complet à 65 % fut mis au jour. Stan est actuellement exposé à l'Institut de recherche géologique de Black Hills à Hill City dans le Dakota du Sud, après avoir effectué une tournée mondiale. Les scientifiques, dont Peter Larson qui avait déjà examiné Sue, ont également retrouvé sur ce tyrannosaure de nombreuses lésions osseuses, notamment des fractures consolidées au niveau des côtes et de la nuque ainsi qu'un trou spectaculaire à l'arrière de sa tête, de la taille d'une dent de Tyrannosaurus.
Durant l'été 2000, Jack Horner découvrit cinq squelettes de Tyrannosaurus près de Fort Peck Reservoir dans le Montana. Un des spécimens, surnommé « C. rex » est l'un des plus grands Tyrannosaurus jamais retrouvé[56].
 « Jane », juvénile de Tyrannosaurus, au Cleveland Museum of Natural History, Cleveland, Ohio.
« Jane », juvénile de Tyrannosaurus, au Cleveland Museum of Natural History, Cleveland, Ohio.En 2001, un squelette complet à 50 % d'un juvénile de Tyrannosaurus est découvert dans la formation Hell Creek, par une équipe du Burpee Museum of Natural History de Rockford, Illinois. Surnommé « Jane », le squelette fut initialement considéré comme étant le premier spécimen connu du tyrannosauridé nain Nanotyrannus mais des études ultérieures révélèrent que c'était plus probablement un juvénile de Tyrannosaurus[57]. Il s'agit du juvénile le plus complet et le mieux préservé à ce jour. Jane fut examiné par Jack Horner, Pete Larson, Robert Bakker, Greg Erickson, et plusieurs autres paléontologues de renom. Elle est exposé au Burpee Museum of Natural History de Rockford, Illinois[58],[59].
En avril 2006, la Montana State University révéla qu'elle était en possession du plus grand crâne de Tyrannosaurus jamais découvert. Trouvé dans les année 1960 et seulement récemment remonté, le crâne mesure 150 cm de long, comparé aux 141 cm de Sue, une différence de 6,5 %[60],[61].
Paléobiologie
 Vue d'artiste d'un tyrannosaure, la robe de l'animal demeure inconnue et ne peut qu'être imaginée.
Vue d'artiste d'un tyrannosaure, la robe de l'animal demeure inconnue et ne peut qu'être imaginée.Le paléontologue Jack Horner a découvert cinq tyrannosaures dans un même gisement. Il suppose qu'ils vivaient en groupe. Ces individus n'ont pas pu être attirés par une proie ou une charogne : aucun reste d'herbivore n'a été trouvé à cet endroit.
Jack Horner s'est demandé si le tyrannosaure était un chasseur ou un charognard. En effet il n'est pas un bon coureur. Cependant, il aurait eu un flair très sensible, compensant ainsi sa mauvaise vision : cela lui aurait donc permis de repérer facilement une charogne. Néanmoins, on a découvert une morsure de tyrannosaure sur un edmontosaurus qui avait cicatrisé ; l'animal est décédé postérieurement. De plus le tyrannosaure détenait une mâchoire d'une puissance phénoménale, qu'il devait probablement utiliser pour tuer des animaux vivants, ce qui en ferait un chasseur.
Les scientifiques se demandent donc toujours s'il mangeait de la viande fraîche ou de la charogne. Il n'est d'ailleurs pas exclu qu'il ait pu être à la fois prédateur et charognard, selon les opportunités qui se présentaient à lui. À titre de comparaison, dans les comportements que l'on peut observer de nos jours, il peut arriver que les vautours par exemple, des charognards, chassent lorsque la faim les y pousse. À l'inverse, les lions, découvrant un cadavre encore frais, n'hésitent pas à s'en repaître. Ils s'avèrent par là, opportunistes.
Des scientifiques ont découvert dans les fossiles de T. Rex, des traces de goutte. Ce grand carnivore devait donc souffrir de douleurs aiguës et soudaines dans les articulations, ce qui devait probablement jouer sur son comportement[62].
Durée de vie
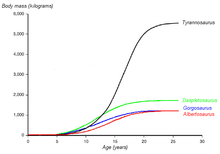 Courbe de croissance de quatre tyrannosauridés (poids et âge). La courbe de Tyrannosaurus rex est en noir. D'après Erickson et al. 2004.
Courbe de croissance de quatre tyrannosauridés (poids et âge). La courbe de Tyrannosaurus rex est en noir. D'après Erickson et al. 2004.L'identification de plusieurs spécimens de jeunes Tyrannosaurus rex a permis aux scientifiques de documenter les changements ontogénétiques, d'évaluer le cours de leur vie, et de déterminer la croissance de ces animaux. Le plus petit individu connu (LACM 28471, le théropode « Jordan ») devait peser seulement 29,9 kg, alors que les plus grands, comme le spécimen FMNH PR2081 (Sue), devaient bien atteindre plus de 5 400 kg (soit presque 6 tonnes). Des analyses histologiques d'os de T. rex ont démontré que le spécimen LACM 28471 était âgé de deux ans seulement, alors que Sue avait 28 ans, probablement l'âge limite de cette espèce[3].
L'histologie a également permis de déterminer l'âge d'autres spécimens. Il est possible de dessiner les courbes de croissance lorsqu'on peut reporter sur un graphique l'âge et le poids de différents spécimens. Celle du T. rex est une courbe en S, les jeunes ne dépassaient pas les 1 800 kg jusqu'à 14 ans environ, puis leur taille augmentait de façon significative. Durant cette phase de croissance rapide, un jeune T. rex pouvait gagner en moyenne 600 kg par an pendant quatre ans. À partir de 18 ans, la courbe se stabilise, ce qui signifie que la croissance de l'animal augmente plus lentement. Par exemple, seulement 600 kg séparent le spécimen Sue âgé de 28 ans du spécimen canadien âgé quant à lui de 22 ans (RTMP 81.12.1)[3]. Une autre étude histologique plus récente menée par différents scientifiques corrobore ces résultats, démontrant que la croissance rapide commence à ralentir à partir de 16 ans[63]. Cette cassure brutale de la vitesse de croissance entre 14 et 18 ans pourrait témoigner de l'existence d'une phase de maturité physique, synonyme de maturité sexuelle. Une hypothèse soutenue par la découverte en 2005 de tissus osseux riches en calcium, connus aussi sous le nom d'os médullaires, dans le fémur d'une T. rex âgée de 18 ans (MOR 1125, également connu sous le nom de « B-rex »)[64]. Dans la nature, ces tissus osseux ne sont retrouvés que chez les oiseaux femelles matures juste avant la ponte; ils permettent de renforcer la coquille des œufs. Les dinosaures étant également ovipares, le même phénomène intervenait peut être à l'époque[65],[66] et cette découverte pourrait donc indiquer que la jeune « B-rex » était sexuellement mature[67]. La même découverte a été réalisée chez une femelle Allosaurus âgée de 10 ans et chez une femelle Tenontosaurus âgée de 8 ans, laissant penser que la maturité sexuelle des dinosaures serait intervenue beaucoup plus tôt que ce qui était jusqu'alors pensé. Des femelles dinosaures pouvaient donc pondre des œufs dès leur pré-adolescence, et de là, devenir mères[66],[68]. D'autres tyrannosauridés ont des courbes de croissances très similaires à celles de T. rex, même si leurs vitesses de croissance plus lentes correspondent à des tailles plus petites à l'âge adulte[69].
Le taux de mortalité augmente à l'approche de la maturité sexuelle, un schéma que l'on retrouve chez les autres tyrannosaures, les grands oiseaux et les mammifères. Ces espèces connaissent après une mortalité infantile élevée, un taux de survivance élevé, qui décline rapidement à la maturité sexuelle. La mortalité augmente à la maturité sexuelle, en partie dû au stress des femelles qui accompagne la ponte. Une étude démontre que le faible nombre d'échantillons de juvéniles est dû en partie à leur taux de mortalité bas, ces animaux ne mouraient pas en grand nombre à ces âges[69].
Dimorphisme sexuel
 Reconstitution d'un couple de Tyrannosaurus en train de s'accoupler (Jurassic Museum of Asturias).
Reconstitution d'un couple de Tyrannosaurus en train de s'accoupler (Jurassic Museum of Asturias).Avec la découverte de spécimens de plus en plus nombreux, les scientifiques ont commencé à analyser les différences entre individus et ils ont identifié chez Tyrannosaurus rex, deux types morphologiques distincts, similaires à d'autres espèces de théropodes. En prenant en compte leur morphologie générale, l'un a été dénommé le morphotype « robuste » alors que l'autre était dénommé « gracile ».
Plusieurs caractéristiques associées à ces deux morphotypes ont permis aux scientifiques d'en déduire qu'il s'agissait possiblement d'un dimorphisme sexuel, et que le type robuste était probablement femelle, alors que le type gracile était probablement mâle. Ainsi le pelvis de plusieurs spécimens robustes est plus large, peut être afin de faciliter le passage des œufs dans le bassin lors de la ponte[70].
De plus chez le type robuste, on retrouve un chevron - os protégeant certains éléments vitaux de la partie ventrale de la queue - de taille réduite au niveau de la première vertébrale caudale, peut être également afin de faciliter le passage des œufs dans le conduit génital[71]. Cette hypothèse avait été déjà proposée par Romer en 1956 pour les crocodiliens[72].Au début des années 2000, l'existence d'un dimorphisme sexuel chez T. rex a été remise en question. Ainsi en 2005, une étude concluait que la présence de chevrons ne permettait pas de différencier le genre des crocodiliens, et jetait ainsi le doute sur le bien fondé de ce critère pour différencier le genre de T. rex[73]. De plus les scientifiques ont découvert que le premier chevron de Sue, un spécimen extrêmement robuste, était en fait situé très près de son pelvis, comme chez certains reptiles mâles, démontrant que la position des chevrons variait trop pour être un bon indicateur du sexe, chez les T. rex comme chez les reptiles modernes[74]. L'existence de différences morphologiques entre les différents spécimens retrouvés pourrait ne pas être liée à un dimorphisme sexuel, mais plutôt à des variations géographiques, ou à des variations dues à l'âge, les individus robustes étant les plus âgés[1].

En 2009, nous ne connaissons avec certitude le sexe que d'un seul et unique spécimen de T. rex. L'examen de « B.rex » (MOR 1125) a permis de retrouver des tissus mous préservés provenant d'os médullaire, un tissu spécialisé retrouvé chez les oiseaux femelles modernes, source de calcium permettant la production de la coquille des œufs lors de l'ovulation. Le tissu osseux médullaire n'est retrouvé naturellement que chez les femelles oiseaux, suggérant fortement que « B.rex » était une femelle, et qu'elle est morte en période ovulatoire[67]. Des études récentes ont montré que le tissu médullaire n'était jamais retrouvé chez les crocodiles, qui sont les animaux vivants les plus proches des dinosaures, avec les oiseaux. La présence partagée de tissu médullaire chez les oiseaux et les dinosaures théropodes montre les liens évolutionnaires étroits entre les deux[75].
Posture
 Ancienne représentation de 1919 par l'artiste Charles R. Knight, suggérant trois points d'appui.
Ancienne représentation de 1919 par l'artiste Charles R. Knight, suggérant trois points d'appui.Comme beaucoup de dinosaures bipèdes, Tyrannosaurus rex a été, de façon erronée, historiquement décrit comme un tripode marchant sur ses deux pattes postérieures avec sa queue servant d'appui au sol : avec le corps presque à la verticale, un peu à la façon d'un kangourou. Cette conception date de la reconstitution en 1865 d'un Hadrosaurus par le paléontologue américain Joseph Leidy, qui fut le premier à décrire un dinosaure en bipédie[76].
 Une réplique de T. rex au Senckenberg Museum, montrant une conception moderne de la posture de T. rex, sur ses deux pattes arrière.
Une réplique de T. rex au Senckenberg Museum, montrant une conception moderne de la posture de T. rex, sur ses deux pattes arrière.Henry Fairfield Osborn, ancien président de l'American Museum of Natural History (AMNH) de New York, pensant que l'animal se tenait à la verticale, inaugura la première reconstitution d'un squelette complet de T. rex en 1915. Celui-ci resta en position debout pendant près d'un siècle, jusqu'à son démantèlement en 1992[77].
Vers 1970, les scientifiques réalisent que cette posture en position verticale avec trois points d'appui n'est pas anatomiquement possible ; elle aurait entraîné la luxation ou la détérioration de nombreuses articulations telles que celles des hanches ou celle comprise entre le crâne et les premières vertèbres cervicales[78].
La reconstitution fausse à l'AMNH a inspiré de nombreux films et peintures jusque dans les années 1990, lorsque les films comme Jurassic Park ont représenté le tyrannosaure dans une posture plus réaliste. Les représentations modernes dans les musées, l'art et les films montre le Tyrannosaurus rex avec le corps presque parallèle au sol et la queue tendue à l'arrière pour équilibrer la tête.
Bras
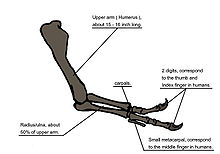 Diagramme illustrant l'anatomie osseuse des bras du Tyrannosaurus rex.
Diagramme illustrant l'anatomie osseuse des bras du Tyrannosaurus rex.Lorsqu'on a découvert le Tyrannosaurus rex, l'humérus était la seule partie connue des pattes antérieures[5]. Le premier squelette montré au public en 1915 et monté par Osborn montraient le Tyrannosaurus rex avec une main à trois doigts, comme pour l'Allosaurus[43]. Pourtant un an plus tôt, Lawrence Lambe avait décrit les membres antérieurs de Gorgosaurus, une espèce proche et apparentée au T. rex, comme étant courts et munis de deux doigts seulement[79]. Ce qui pouvait suggérer que Tyrannosaurus rex avait des bras similaires, hypothèse qui ne fut confirmée que bien plus tard, lorsque les premiers restes de bras entiers furent identifiés en 1989, appartenant à MOR 555, surnommé le "Wankel rex", découvert à Hell Creek[48]. Les restes de "Sue" comprennent également des bras complets[1].
 Détails des bras d'un squelette reconstitué.
Détails des bras d'un squelette reconstitué.Les bras du Tyrannosaurus rex sont très petits par rapport au reste du corps, mesurant seulement 1 mètre de long. Cependant ces bras ne sont pas des structures vestigiales et montrent de larges zones d'attachement musculaires, indiquant une force considérable. Dès 1906, Osborn avait imaginé que les bras pouvaient être utilisés pour tenir fortement le partenaire lors de l'accouplement[46]. Il a été également suggéré que les bras soient utilisés pour aider l'animal à se redresser à partir d'une position couchée[78]. Ou pour tenir sa proie alors que les mâchoires font le reste, hypothèse qui serait soutenue par des analyses biomécaniques. Ainsi les os des bras possèdent une corticale particulièrement résistante. Le muscle biceps brachial d'un adulte était capable de soulever à lui seul une charge de 199 kg, chiffre augmentant avec l'action combinée des autres muscles du bras comme le brachial. Les mouvements des bras étaient limités, avec des amplitudes articulaires de respectivement 40 et 45 ° aux épaules et aux coudes - en comparaison, chez Deinonychus, les amplitudes de ces deux mêmes articulations sont 88 et 130 °, et chez l'homme 360 et 165 °. Ainsi, les bras du Tyrannosaurus rex, avec leur structure osseuse particulièrement solide, la force considérable développée par leurs muscles et leur amplitude de mouvement limitée, pourrait indiquer un système anatomique conçu pour agripper vite et fort une proie se débattant[80].
Peau et plumes
 Restauration d'un jeune Tyrannosaurus, représenté avec des plumes.
Restauration d'un jeune Tyrannosaurus, représenté avec des plumes.En 2004, la revue scientifique Nature a publié un rapport décrivant un tyrannosauridé, le Dilong paradoxus, un ancêtre du T. Rex retrouvé sur le site géologique de la formation Jehol en Chine. Comme de nombreux théropodes découverts dans cette formation, le squelette fossilisé a été préservé dans un manteau de structures filamenteuses reconnues comme étant des structures précurseurs des plumes. C'est ainsi qu'on a également supposé que le Tyrannosaurus et d'autres tyrannosauridés proches possédaient des protoplumes. Cependant, des empreintes de peau retrouvées sur de grands tyrannosauridés montrent des écailles en mosaïques[81]. Il est possible que ces protoplumes aient existé sur des parties du corps qui n'ont pas été préservé. Mais les gros animaux ont, proportionnellement à leur volume, une surface moins importante que les plus petits. Donc plus un animal est gros, moins il souffre des déperditions de chaleur. Les protoplumes deviendraient alors inutiles et pourraient avoir été secondairement perdues lors de l'évolution des grands tyrannosauridés comme Tyrannosaurus, particulièrement sous le chaud climat du crétacé[82].
Thermorégulation
On a longtemps pensé que le Tyrannosaurus, comme la plupart des dinosaures était poïkilotherme, c'est-à-dire qu'il ne contrôlait pas sa température corporelle et avait le « sang froid » , tout comme les reptiles. C'est dans les années 1960 que des scientifiques comme Robert T. Bakker et John Ostrom ont émis l'idée que le métabolisme des dinosaures ressemblait davantage à celui des mammifères et des oiseaux qu'à celui des animaux à sang froid[83],[84]. Suite à l'analyse d'un squelette, des scientifiques ont déclaré que le Tyrannosaurus rex était homéotherme (à sang chaud), impliquant ainsi une vie très active[9]. Depuis, plusieurs paléontologues ont cherché à déterminer la capacité du Tyrannosaurus à réguler la température de son corps. Des preuves histologiques de taux de croissance rapides chez le jeune T. rex, comparables à ceux des mammifères et des oiseaux, pourrait supporter l'hypothèse d'un métabolisme élevé. Les courbes de croissance indiquent que, tout comme chez les mammifères et les oiseaux, la croissance du Tyrannosaurus rex est limitée à l'âge adulte, contrairement à la croissance indéterminée retrouvée chez de nombreux autres vertébrés[63].
Les ratios d'isotopes d'oxygène présents dans les os fossilisés sont utilisés pour déterminer la température à laquelle l'os a été formé, car ces ratios sont corrélés avec la température. Chez un spécimen de T. rex on a retrouvé des ratios indiquant que la différence de température entre une vertèbre du thorax et un tibia n'était que de 4 à 5 °C. Selon le paléontologue Reese Barrick et le géochimiste William Showers cette différence minime entre le corps de l'animal et ses extrémités indique que Tyrannosaurus rex maintenait sa température interne corporelle constante, définition de l'homéothermie, et qu'il possédait un métabolisme situé entre celui des reptiles ectothermes et des mammifères endothermes[85]. D'autres scientifiques ont fait remarquer que le ratio des isotopes d'oxygène présent dans les fossiles actuels était probablement différent du ratio originel au temps où Tyrannosaurus vivait, et qu'il avait dû être modifié durant ou après le processus de fossilisation appelé la diagenèse[86]. Barrick et Showers se sont défendus en retrouvant des résultats similaires chez un autre dinosaure théropode, le Giganotosaurus, vivant sur un autre continent et dix millions d'années plus tôt[87].
Des ornithischiens montrent également des preuves d'homéothermie, alors que ce n'est pas le cas pour des varans retrouvés dans la même formation[88]. Même si le Tyrannosaurus rex montre des preuves d'homéothermie, cela ne signifie pas obligatoirement qu'il était endotherme. Une telle thermorégulation peut s'expliquer par la gigantothermie, comme chez certaines tortues de mer[89],[90]. Ainsi certains grands poïkilothermes, grâce à un rapport volume / surface de peau favorable, sont capables de maintenir une température de corps et un métabolisme relativement élevés.
Locomotion
 Réplique d'une séquence de traces de pas d'un théropode attribuée à Megalosaurus à l'OUMNH. Aucune séquence de ce type n'a été retrouvée pour les tyrannosaures, rendant difficile l'estimation de leur allure et de leur vitesse.
Réplique d'une séquence de traces de pas d'un théropode attribuée à Megalosaurus à l'OUMNH. Aucune séquence de ce type n'a été retrouvée pour les tyrannosaures, rendant difficile l'estimation de leur allure et de leur vitesse.Il y a deux questions principales qui font débat concernant les capacités locomotrices du Tyrannosaurus : comment pouvait-il tourner et quelle était sa vitesse de pointe. Ces deux questions sont liées au débat concernant le fait de savoir s'il était chasseur ou charognard.
Le Tyrannosaurus devait être lent à se retourner, prenant probablement une à deux secondes pour se tourner de 45° — à titre de comparaison l'Homme, qui se tient debout et n'a pas de queue, peut se retourner en une fraction de seconde[91]. Cette lenteur est due au moment d'inertie, qui quantifie la résistance d'un corps soumis à une mise en rotation et qui est particulièrement important chez le Tyrannosaurus, dont une grande partie de la masse est située à distance de son centre de gravité. Il pouvait diminuer cette résistance en arquant son dos et sa queue et en rapprochant sa tête et ses bras de son corps, à la façon d'un patineur qui se regroupe pour tourner sur lui-même plus vite[92].
Les scientifiques ont avancé de nombreux chiffres concernant la vitesse maximale de course de Tyrannosaurus rex, la plupart autour de 11 m/s soit 40 km/h, avec des minima de 5–11 m/s (18–40 km/h) et des maxima autour de 20 m/s (72 km/h). Diverses techniques d'estimation ont été utilisées pour aboutir à ces chiffres car, s'il existe de nombreuses traces de pas de grands théropodes en train de marcher, aucune trace de théropodes en train de courir n'a été encore retrouvé, ce qui pourrait indiquer qu'ils ne couraient simplement pas[93]. Les scientifiques qui pensent que Tyrannosaurus était capable de courir soulignent que certaines caractéristiques anatomiques permettent à un adulte de ne peser que 4,5 tonnes et que certains animaux tels que les autruches ou les chevaux, possédant de longues jambes flexibles, sont capables d'atteindre de grandes vitesses grâce à des foulées plus lentes mais plus longues. De plus, certains avancent que Tyrannosaurus avait des muscles aux membres inférieurs plus larges que chez n'importe quel animal vivant actuel, ce qui lui aurait permis de courir jusqu'à 40 à 70 kilomètres par heure[94].
Jack Horner et Don Lessem avancèrent en 1993 que le Tyrannosaurus était lent et ne pouvait probablement pas courir (présence d'une phase de suspension en l'air) car le ratio de la longueur de son fémur sur celle de son tibia était supérieur à 1, comme chez la plupart des grands théropodes et comme les éléphants modernes[48]. Cependant, Holtz écrit en 1998 que les tyrannosauridés et des groupes proches avaient les éléments distaux des membres postérieures (cheville, métatarse et orteils) significativement plus longs que la longueur du fémur de la plupart des autres théropodes, et que les tyrannosauridés et des groupes proches avait un métatarse plus efficace pour transmettre les forces de locomotion du pied à la jambe que chez les premiers théropodes. Il conclut que les tyrannosauridés et des groupes proches étaient les plus rapides des larges théropodes[95].
Christiansen écrit en 1998 que les os de la jambe de Tyrannosaurus n'étaient pas significativement plus solides que ceux des éléphants, qui sont relativement limités concernant leur vitesse de pointe et qui n'ont jamais pu courir (pas de phase de suspension en l'air), et il proposa que la vitesse maximale de Tyrannosaurus devait être d'environ 11 mètres par seconde (40 km/h), ce qui est comparable à la vitesse d'un sprinter humain. Il fit remarquer que cette estimation dépendait de nombreuses hypothèses douteuses[96].
Farlow et ses collègues avancent en 1995 qu'un Tyrannosaurus pesant 5,4 à 7,3 tonnes serait gravement voire fatalement blessé s'il tombait alors qu'il était en train de se mouvoir rapidement, car son thorax frapperait le sol avec une décélération de 6 g sans que ses petits bras puissent réduire l'impact[10]. Cependant, des animaux comme les girafes peuvent galoper à 50 km/h malgré le risque que cela représente[97],[98] et il est donc possible que Tyrannosaurus ait également pu se déplacer rapidement en cas de nécessité et malgré les risques que cela pouvait représenter[99],[100].
Des études récentes sur la vitesse de déplacement de Tyrannosaurus avançaient une allure modérée ne dépassant pas 40 km/h. Par exemple, en 2002 dans la revue Nature, un modèle mathématique est présenté[101] dont le but est de permettre d'estimer la masse musculaire nécessaire au niveau des jambes pour courir rapidement, à plus de 40 km/h[94]. L'article conclut que des vitesses supérieures à 40 km/h sont impossibles à atteindre car elles nécessiteraient de très gros muscles représentant plus de 40 à 86 % de la masse corporelle totale. Si ces muscles avaient été moins massifs, seule une vitesse d'environ 18 km/h aurait pu être atteinte. Faire des conclusions sur cette modélisation est difficile car on ne sait pas quel était le volume des muscles de Tyrannosaurus[94].
Une étude de 2007 utilisant des modèles informatiques estimait la vitesse maximale de course de T. rex à 8 mètres par seconde, soit 29 km/h[102], soit légèrement plus vite qu'un footballeur professionnel et moins vite qu'un sprinter qui peut atteindre 12 mètres par seconde soit 43 km/h[103].
Ceux qui estiment que Tyrannosaurus était incapable de courir estiment sa vitesse de pointe à 17 km/h, ce qui est toujours plus rapide que les hadrosauridés et les cératopsiens qui devaient être ses principales proies[94]. De plus, certains de ceux qui défendent le fait que Tyrannosaurus était un prédateur avancent que sa vitesse de course n'était pas si cruciale, car même s'il était lent, il était toujours plus rapide que ses proies[104]. Cependant, Paul et Christiansen (2000) écrivent que les derniers Ceratopsia avaient des pattes arrières verticales et que les gros spécimens devaient pouvoir courir aussi vite que les rhinocéros[105]. On a retrouvé des fossiles de Ceratopsia présentant des cicatrices de morsures de Tyrannosaurus. Or, si les Ceratopsia ayant vécu en même temps que les T. rex étaient rapides, cela jette le doute sur l'argument que Tyrannosaurus n'avait pas à être rapide pour attraper ses proies[100].
Empreintes de pas
 Probable empreinte de pas d'un Tyrannosaurus retrouvée au Nouveau-Mexique.
Probable empreinte de pas d'un Tyrannosaurus retrouvée au Nouveau-Mexique.Deux différentes empreintes de pas fossilisées ont été proposées comme appartenant à Tyrannosaurus rex. La première a été découverte au Philmont Scout Ranch dans l'État du Nouveau-Mexique, en 1983, par le géologue américain Charles Pillmore. On pensait initialement qu'elle appartenait à un hadrosauridé ou « dinosaure à bec de canard » mais l'examen attentif de l'empreinte de pas révéla d'une part un large talon inconnu chez les ornithopodes et d'autre part les restes de ce qui aurait pu être un hallux, quatrième orteil en forme d'ergot. Cette empreinte fut décrite comme appartenant à l'ichnotaxon Tyrannosauripus pillmorei en 1994 par Martin Lockley et Adrian Hunt, qui suggéraient qu'elle pouvait avoir été faite par un Tyrannosaurus rex, ce qui en ferait la première empreinte de pas connue de l'espèce. La trace est imprimée dans ce qui fut autrefois de la boue provenant d'une terre humide végétale. Elle mesure 83 cm de longueur sur 71 cm de largeur[106].
Une seconde empreinte de pas qui pourrait avoir été faite par un Tyrannosaurus fut découverte dans la formation de Hell Creek dans le Montana en 2007 par le paléontologue britannique Phil Manning. Cette seconde trace de 76 cm de long est plus petite que celle décrite par Lockley et Hunt. Qu'elle appartienne à un Tyrannosaurus n'est pas certain, bien que Tyrannosaurus et Nanotyrannus sont les deux seuls grands théropodes qui ont été retrouvés à Hell Creek. Des études plus approfondies, comparant notamment cette trace à celle du Nouveau-Mexique, sont prévues[107].
Alimentation
Le débat concernant le comportement charognard ou prédateur du Tyrannosaurus est aussi vieux que celui sur sa locomotion.
En 1917, Lambe étudie un squelette de Gorgosaurus, une espèce proche de Tyrannosaurus, et conclut que le Tyrannosaurus était un pur charognard car les dents du Gorgosaurus ne montraient aucune trace d'usure[108]. Cet argument n'est plus considéré comme valide depuis que l'on sait que les théropodes remplacent leurs dents rapidement.
Depuis la première découverte de Tyrannosaurus, la plupart des scientifiques ont la certitude qu'il est un prédateur, bien que tout comme de nombreux prédateurs modernes, il aurait pu aussi voler la prise d'un autre prédateur ou être parfois charognard si l'occasion se présentait[109].
Jack Horner est le principal défenseur de la théorie du tyrannosaure charognard[48],[110],[111]. Il a présenté plusieurs arguments défendant cette hypothèse :
- Les bras du tyrannosaure sont courts comparés à ceux des prédateurs connus. Selon Horner, les bras sont trop courts pour avoir la force nécessaire pour maintenir une proie[112].
- Les tyrannosaures avaient de grands bulbes et nerfs olfactifs par rapport à la taille de leur cerveau. Cela suppose qu'ils possédaient un odorat bien développé qui aurait pu leur permettre de repérer des carcasses à de grandes distances comme les vautours modernes. Des recherches faites sur les bulbes olfactifs de 21 dinosaures démontrent que c'est le tyrannosaure qui avait l'odorat le plus développé[113].
- Les dents du tyrannosaure pouvaient broyer les os, et donc extraire des carcasses, généralement les parties les moins nutritives, autant de nourriture (la moelle osseuse) que possible. Karen Chin et son équipe ont trouvé des fragments d'os dans des coprolithes (excréments fossilisés) attribués à des tyrannosaures, mais ils indiquent que les dents du tyrannosaure n'étaient pas adaptées pour broyer systématiquement des os comme le font les hyènes pour extraire la moelle[114].
- Comme les proies potentielles du Tyrannosaurus pouvaient se déplacer rapidement, les preuves qu'il ne pouvait pas courir vite démontrent qu'il était un charognard[110],[115]. D'un autre côté, des analyses récentes démontrent que le Tyrannosaurus bien que plus lent que certains prédateurs terrestres modernes pouvait être assez rapide pour chasser de grands hadrosaures et des cératopsiens[94],[104].
 Les orbites du tyrannosaure regardent vers l'avant, permettant une vision binoculaire stéréoscopique avantageant les prédateurs.
Les orbites du tyrannosaure regardent vers l'avant, permettant une vision binoculaire stéréoscopique avantageant les prédateurs.D'autres preuves suggèrent un comportement de prédateur. Les orbites du tyrannosaure étaient placées vers l'avant, lui donnant ainsi une vision binoculaire ce qui lui permettait d'évaluer les distances, bien mieux que les faucons modernes. Horner note aussi une amélioration importante de la vision binoculaire tout au long de l'évolution des tyrannosauridés, avançant qu'il n'était pas évident que la sélection naturelle favorise cette caractéristique si les tyrannosaures avaient été de purs charognards, pour qui une vision stéréoscopique n'aurait pas été un atout[19],[20]. Chez les animaux modernes, la vision binoculaire est principalement retrouvée chez les prédateurs[116].
Selon certains scientifiques, si le Tyrannosaurus était un pur charognard, un autre dinosaure devait être le superprédateur du Crétacé supérieur amérasien. Les autres tyrannosauridés partageant les caractéristiques de Tyrannosaurus, seuls les petits dromæosauridés auraient le potentiel pour être ce superprédateur. Les proies en haut de la chaîne alimentaire étaient alors les marginocéphales et les ornithopodes. Des supporters de l'hypothèse du charognard suggèrent que la taille et la puissance de Tyrannosaurus lui aurait alors permis de voler les proies tuées par de plus petits prédateurs[115]. La plupart des paléontologues acceptent l'hypothèse que le Tyrannosaurus était à la fois prédateur et charognard, comme beaucoup de grands carnivores.
Salive infectée
William Abler a proposé que Tyrannosaurus avait peut-être une salive infectieuse, qu'il utilisait pour tuer sa proie[117]. En examinant les dents de tyrannosaures, il s'est aperçu qu'il existait des dentelures pouvant, comme chez le Dragon de Komodo, retenir des morceaux de carcasse permettant la prolifération de bactéries, rendant leur morsure infectante. Horner note cependant que les dentelures de Tyrannosaurus ressemblent plus à des cubes alors que celles des Komodo sont arrondies[118].
Le tyrannosaure dans la culture populaire
Depuis sa découverte en 1905, le tyrannosaure est devenu le dinosaure le plus célèbre dans la culture populaire. C'est le seul dinosaure dont le nom scientifique Tyrannosaurus rex est connu du grand public, de même son abréviation T. rex est aussi répandue. Les expositions du tyrannosaure sont très populaires ; on estime à 10 000 le nombre de visiteurs venus au Field Museum de Chicago en 2003 pour voir Sue, le fossile le plus complet exposé.
Le tyrannosaure est apparu plusieurs fois à la télévision et au cinéma, notamment dans le Monde perdu, King Kong, Fantasia, Un million d'années avant J.C., plusieurs versions filmiques et de dessins animés avec Godzilla (monstre des croyances d'après-guerre, suite aux chocs des bombes atomiques de Hiroshima et Nagasaki) comme héros, et Jurassic Park. De nombreux livres et bandes dessinées, dont Calvin et Hobbes, et le manga Gon de Masashi Tanaka, ont aussi représenté le tyrannosaure, qui est souvent montré comme le plus grand et le plus terrifiant carnivore de tous les temps. Le groupe de rock T-Rex a pris ce nom d'après l'espèce, le troisième album du groupe The Hives est intitulé Tyrannosaurus Hives.
Cinéma
- Dans King Kong, réalisé en 1933 par Merian C. Cooper et Ernest B. Schoedsack, le gorille géant affronte un Tyrannosaurus pour protéger la jeune femme Ann. Jack. Dans le remake de 2005 de Peter Jackson, le tyrannosaure est remplacé par un « Vastatosaurus rex », une évolution imaginaire du T. rex.
- Dans le Sacre du printemps, un passage du film Fantasia, produit par Walt Disney Pictures, un Tyrannosaurus terrorise d'autres dinosaures et engage un combat avec un Stegosaurus, ce qui est un anachronisme, les deux animaux ayant vécu à des périodes différentes.
- Pour créer le célèbre monstre japonais Godzilla, les designers de Toho se sont inspirés du stégosaure pour les plaques dorsales, du tyrannosaure pour la tête et de l'Allosaurus pour le corps.
- Une des apparitions les plus célèbres du Tyrannosaurus se trouve dans Jurassic Park, réalisé en 1993 par Steven Spielberg. On peut voir le dinosaure s'évader de son enclos et parcourir le parc pour chasser les visiteurs et les employés.
- Dans le Monde Perdu, un Tyrannosaurus est attrapé et emmené à San Diego.
- Dans La Nuit au musée, un squelette de Tyrannosaurus prend vie une fois que la nuit est tombée et se comporte comme un chien en rapportant un os que le gardien lui lance.
Dessins animés
- Dans Toy Story, Toy Story 2 ou Toy Story 3, le personnage Rex est un Tyrannosaurus en plastique.
- Dans le Petit Dinosaure, Petit-Pied et ses amis fuient un Tyrannosaurus qu'ils appellent « Dents tranchantes ». Par la suite, ils adopteront Gobeur, un bébé T. Rex.
- Dans Dinosaur King, le Gang Alpha possède un Tyrannosaurus nommé Terry.
Littérature
- Dans le manga de Masashi Tanaka, Gon est un bébé tyrannosaure à la force redoutable.
- Dans la bande dessinée Calvin et Hobbes de Bill Watterson, Calvin, un enfant de six ans, aime se transformer en dinosaure, et en général en Tyrannosaurus.
Jeux
Très présent dans les jeux vidéo, il constitue souvent un ennemi redoutable comme dans Dino Crisis, Tomb Raider, Turok,Paraworld ou encore King Kong. Il est aussi présent dans le jeu vidéo Jurassic Park Opération Genesis. Il est aussi décliné en jouets et maquettes. Son image est assez prisée dans la publicité.
Notons aussi l'unique apparition de Gon (du manga éponyme) en tant que personnage jouable dans la version Playstation de Tekken 3.
Valeur marchande
Le 4 octobre 1997, chez Sotheby's à New York un squelette de Tyrannosaurus rex, renommé Sue d'après le nom de sa découvreuse, fut adjugé à 8 362 500 dollars, la somme la plus importante jamais payée pour un fossile[119],[120]. Parmi neuf enchérisseurs, ce fut le Field Museum de Chicago, qui remporta les enchères notamment grâce à des partenaires industriels.
En mars 2007, un crâne de Tyrannosaurus rex fut acheté 276 000 dollars par un collectionneur de Californie, la troisième plus importante somme jamais payée pour un spécimen préhistorique[121].
Divers
Le tyrannosaure n'est pas le plus grand des dinosaures carnivores. Voici quelques-uns des plus grands :
- Giganotosaure de 14 mètres (Amérique du Sud)
- Carcharodontosaure dont la taille varie de 8 à 12 mètres (Afrique)
- Spinosaure de 12 mètres et peut-être 17 (Afrique)
- Mapusaure de 13 mètres (Amérique du Sud)
Le tyrannosaure ne les a cependant jamais croisés, tous ayant vécu dans des régions et/ou des périodes différentes.
-

Squelette de tyrannosaure
-
Squelette de tyrannosaure
-

Détail membre antérieur
-

Détail des mâchoires
Annexes
Articles connexes
- Voir Liste de dinosaures
- Theropoda | Tyrannosauridae
- Nanotyrannus : peut-être la forme juvénile de Tyrannosaurus rex
- Tarbosaurus : peut-être le représentant asiatique de Tyrannosaurus rex
Liens externes
- (fr) Description du dinosaure Tyrannosaurus sur PaleoWiki
- (fr) Tyrannosaurus sur Gigadino
- (fr) Le tyrannosaure sur Terra Nova
Références taxinomiques
- Référence The Paleobiology database : Tyrannosaurus Osborn 1905 (en)
Bibliographie
- Tyrannosaurus rex, the tyrant king. Par Pedro Larson, Peter L. Larson, Kenneth Carpenter. Édition: illustrated. Publié par Indiana University Press, 2008. (ISBN 0-253-35087-5), 9780253350879. 435 pages. Aperçu en ligne
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Tyrannosaurus » (voir la liste des auteurs)
- (en) Brochu Christopher A., Osteology of Tyrannosaurus Rex: Insights from a Nearly Complete Skeleton and High-resolution Computed Tomographic Analysis of the Skull, Northbrook, Illinois, 2003 (réimpr. Society of Vertebrate Paleontology) (OCLC 51651461)
- (en)Sue's vital statistics, Sue at the Field Museum, Field Museum of Natural History. Consulté le 2007-09-15
- (en) Gregory M. Erickson, Peter J. Makovicky, Philip J. Currie, Norrel, Mark A., Yerby, Scott A. et Christopher A., « Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs », dans Nature, vol. 430, no 7001, 204, p. 772–775 [lien DOI]
- (en)Brian Switek, « Tyrannosaurus yeck?: Another look at preserved proteins » sur http://scienceblogs.com, 30. Consulté le 12 février 2009
- (en) H. F. Osborn, « Tyrannosaurus and other Cretaceous carnivorous dinosaurs », dans Bulletin of the AMNH, New York, American Museum of Natural History, vol. 21, no 14, 1905, p. 259–265 [texte intégral]
- (en)Dynamosaurus imperiosus sur DinoData
- (en) DM Henderson, « Estimating the masses and centers of mass of extinct animals by 3-D mathematical slicing », dans Paleobiology, vol. 25, no 1, 1999, p. 88–106 [texte intégral]
- (en) JF Anderson, AJ Hall-Martin et Dale Russell, « Long bone circumference and weight in mammals, birds and dinosaurs », dans Journal of Zoology, vol. 207, no 1, 1985, p. 53–61
- Robert T. Bakker, The Dinosaur Heresies, New York, Kensington Publishing, 1986 (ISBN 0-688-04287-2) (OCLC 13699558)
- (en) JO Farlow, MB Smith et JM Robinson, « Body mass, bone "strength indicator", and cursorial potential of Tyrannosaurus rex », dans Journal of Vertebrate Paleontology, vol. 15, no 4, 1995, p. 713–725 [texte intégral]
- (en) Frank. Seebacher, « A new method to calculate allometric length-mass relationships of dinosaurs », dans Journal of Vertebrate Paleontology, vol. 21, no 1, 2001, p. 51–60 [lien DOI]
- (en) Per Christiansen, Fariña et Richard, « Mass prediction in theropod dinosaurs », dans Historical Biology, vol. 16, no 2-4, 2004, p. 85–92 [lien DOI]
- (en) Cristiano dal Sasso, « New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities », dans Journal of Vertebrate Paleontology, vol. 25, no 4, 2005, p. 888–896 [texte intégral, lien DOI]
- (en) Jorge O. Calvo et Rodolfo Coria, « New specimen of Giganotosaurus carolinii (Coria & Salgado, 1995), supports it as the as the largest theropod ever found », dans Gaia Revista de Geociências, vol. 15, décembre 1998, p. 117–122 [texte intégral [PDF]]
- (en) Elizibeth D. Quinlan, « Anatomy and function of digit III of the Tyrannosaurus rex manus », dans Geological Society of America Annual Meeting - Abstracts with Programs, 2007, p. 77 [texte intégral] [abstract only]
- (en)Brian Switek, « A Tyrannosaurus with THREE fingers? » sur [http://www.scienceblogs.com Scienblogs, 11. Consulté le 12 février 2009
- (en) Museum unveils world's largest T-rex skull sur [tp://www.montana.edu Montana State University], Montana State University, 2006-04-07. Consulté le 13 septembre 2008
- (en) LM Witmer et RC Ridgely, « The paranasal air sinuses of predatory and armored dinosaurs (archosauria: theropoda and ankylosauria) and their contribution to cephalic structure. », dans Anat. Rec., vol. 291, no 11, novembre 2008, p. 1362-88 [résumé, texte intégral (pages consultées le 12 février 2009)]
- Kent A. Stevens, « Binocular vision in theropod dinosaurs », dans Journal of Vertebrate Paleontology, vol. 26, no 2, juin 2006, p. 321–330 [texte intégral [PDF], lien DOI]
- Eric Jaffe, « Sight for 'Saur Eyes: T. rex vision was among nature's best », dans Science News, vol. 170, no 1, 2006-07-01, p. 3 [texte intégral, lien DOI (pages consultées le 2008-10-06)]
- Eric Snively, « Fused and vaulted nasals of tyrannosaurid dinosaurs: Implications for cranial strength and feeding mechanics », dans Acta Palaeontologica Polonica, vol. 51, no 3, 2006, p. 435–454 [texte intégral [PDF] (page consultée le 2008-10-08)]
- G.M. Erickson, « Bite-force estimation for Tyrannosaurus rex from tooth-marked bones », dans Nature, vol. 382, 1996, p. 706–708 [lien DOI]
- M.B. Meers, « Maximum bite force and prey size of Tyrannosaurus rex and their relationships to the inference of feeding behavior », dans Historical Biology: A Journal of Paleobiology, vol. 16, no 1, août 2003, p. 1–12 [texte intégral, lien DOI]
- Thomas R. Holtz, « The Phylogenetic Position of the Tyrannosauridae: Implications for Theropod Systematics », dans Journal of Palaeontology, vol. 68, no 5, 1994, p. 1100–1117 [texte intégral (page consultée le 2008-10-08)]
- (en) Paul, Gregory S., Predatory dinosaurs of the world: a complete illustrated guide, New York, Simon and Schuster, 1988 (ISBN 978-0-671-61946-6) (OCLC 18350868) (LCCN 88023052)
- Smith, J.B., « Heterodonty in Tyrannosaurus rex: implications for the taxonomic and systematic utility of theropod dentitions », dans Journal of Vertebrate Paleontology, vol. 25, no 4, décembre 2005, p. 865–887 [texte intégral [PDF], lien DOI]
- Douglas K, Young S, « The dinosaur detectives », dans New Scientist, 1998 [texte intégral (page consultée le 2008-10-16)] :
« One palaeontologist memorably described the huge, curved teeth of T. rex as 'lethal bananas' »
- (en) Philip J. Currie, Jørn H. Hurum et Karol Sabath, « Skull structure and evolution in tyrannosaurid dinosaurs », dans Acta Palaeontologica Polonica, vol. 48, no 2, 2003, p. 227–234 [texte intégral [PDF]]
- Thomas R., Jr. Holtz, The dinosauria, Berkeley, David B. Weishampel, Peter Dodson et Halszka Osmólska, 2004, 111–136 p. (ISBN 0-520-24209-2)
- (ru) E. A. Maleev, « Dinosaures carnivores géants de Mongolie », dans Doklady Akademii Nauk, vol. 104, no 4, 1955, p. 634–637
- AK Rozhdestvensky, « Growth changes in Asian dinosaurs and some problems of their taxonomy », dans Paleontological Journal, vol. 3, 1965, p. 95–109
- (en) Kenneth Carpenter, Aspects of nonmarine Cretaceous geology, Beijing, China Ocean Press, 1992 (ISBN 9787502714635) (OCLC 28260578), « Tyrannosaurids (Dinosauria) of Asia and North America »
- Thomas D. Carr, « A New Genus and Species of Tyrannosauroid from the Late Cretaceous (Middle Campanian) Demopolis Formation of Alabama », dans Journal of Vertebrate Paleontology, vol. 25, no 1, mars 2005, p. 119–143 [lien DOI]
- Jørn H. Hurum et Karol Sabath, « Giant theropod dinosaurs from Asia and North America: Skulls of Tarbosaurus bataar and Tyrannosaurus rex compared », dans Acta Palaeontologica Polonica, vol. 48, no 2, 2003, p. 161–190 [texte intégral [PDF]]
- George Olshevsky, « The origin and evolution of the tyrannosaurids », dans Kyoryugaku Saizensen [Dino Frontline], vol. 9–10, 1995, p. 92–119
- T.D. Carr, « Diversity of late Maastrichtian Tyrannosauridae (Dinosauria: Theropoda) from western North America », dans Zoological Journal of the Linnean Society, vol. 142, 2004, p. 479–523 [lien DOI]
- C.W. Gilmore, « A new carnivorous dinosaur from the Lance Formation of Montana », dans Smithsonian Miscellaneous Collections, vol. 106, 1946, p. 1–19
- R.T. Bakker, « Nanotyrannus, a new genus of pygmy tyrannosaur, from the latest Cretaceous of Montana », dans Hunteria, vol. 1, no 5, 1988, p. 1–30
- TD Carr, « Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Theropoda) », dans Journal of Vertebrate Paleontology, vol. 19, 1999, p. 497–520
- Philip J. Currie, « Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous of Alberta, Canada », dans Acta Palaeontologica Polonica, vol. 42, no 2, 2003, p. 191–226 [texte intégral [PDF] (page consultée le 2008-10-09)]
- (en) T. Rex More Hyena Than Lion sur usnews.com le 24 février 2011
- (fr) Documentaire Coup de jeune sur les dinos produit par National Geographic Channel
- H. F. Osborn, « Skeletal adaptations of Ornitholestes, Struthiomimus, Tyrannosaurus », dans Bulletin of the American Museum of Natural History, New York, American Museum of Natural History, vol. 35, no 43, 1917, p. 733–771 [texte intégral (page consultée le 2008-10-08)]
- (en) , International code of zoological nomenclature, Londres, International Commission on Zoological Nomenclature, 1999, 4e éd. (ISBN 978-0-85301-006-7) (OCLC 183090345) (LCCN 2001347021) [lire en ligne (page consultée le 8 octobre 2008)]
- So why hasn't Tyrannosaurus been renamed Manospondylus?, The Dinosaur FAQ, 2002-08-27. Consulté le 2008-10-08
- Henry Fairfield Osborn, « Tyrannosaurus, Upper Cretaceous carnivorous dinosaur », dans Bulletin of the AMNH, New York, American Museum of Natural History, vol. 22, no 16, 1906, p. 281–296 [texte intégral (page consultée le 2008-10-06)]
- BH Breithaup, « Dynamosaurus Imperiosus and the earliest discoveries of Tyrannosaurus Rex in Wyoming and the West », dans New Mexico Museum of Natural History and Science Bulletin, vol. 35, 2006, p. 257–258 [texte intégral [PDF] (page consultée le 2008-12-09)]
- (en) , The complete T. rex, New York, Simon & Schuster, 1993 (ISBN 978-0-671-74185-3) (LCCN 93000211)
- Footprint of a Giant, Online guide to the continental Cretaceous-Tertiary boundary in the Raton basin, Colorado and New Mexico, United States Geological Survey. Consulté le 2008-10-09
- Preparation and mounting, Sue at the Field Museum, The Field Museum, 2007. Consulté le 11 février 2010
- Erickson, G., Makovicky, P. J., Currie, P. J., Norell, M., Yerby, S., Brochu, C. A., « Gigantism and life history parameters of tyrannosaurid dinosaurs », dans Nature, vol. 430, no 7001, 26 mai 2004, p. 772–775 [texte intégral, lien PMID, lien DOI]
- Brochui, C.A., « Lessons From A Tyrannosaur: The Ambassadorial Role Of Paleontology », dans PALAIOS, vol. 18, no 6, décembre 2003, p. 475 [texte intégral, lien DOI]
- Common Avian Infection Plagued the Tyrant Dinosaurs. Ewan D S Wolff, Steven W Salisbury, John R Horner, David J Varricchio. Infectious Diseases/Protozoal Infections Common Avian Infection Plagued the Tyrant Dinosaurs Tyrannosaur ... Common Avian Infection Plagued the Tyrant Dinosaurs ... that tyrannosaurids were commonly infected by a Trichomonas gallinae. PLoS ONE: Research Article, published 30 Sep 2009 10.1371/journal.pone.0007288
- University of Wisconsin-Madison (30 septembre 2009). Was Mighty T. Rex 'Sue' Felled By A Lowly Parasite? ScienceDaily. Retrieved August 1, 2010, from http://www.sciencedaily.com /releases/2009/09/090929133117.htm
- La petite bête qui dévore la grande. Par J.I. pour Sciences-et-Avenir.com le 30/09/2009.
- Dig pulls up five T. rex specimens, BBC News, 10 octobre 2000. Consulté le 13 décembre 2008
- PJ Currie, « Skull structure and evolution in tyrannosaurid dinosaurs », dans Acta Palaeontologica Polonica, vol. 48, no 2, 2003, p. 227–234 [texte intégral [PDF] (page consultée le 2008-10-16)]
- (en) M Henderson, The origin, systematics, and paleobiology of Tyrannosauridae, Dekalb, Illinois, Northern Illinois University Press, In press, « Nano No More: The death of the pygmy tyrant »
- Visit Jane the Dinosaur at the Burpee Museum, Rockford, Illinois. Consulté le 2008-10-16
- Museum unveils world's largest T-rex skull.. Consulté le 2006-04-07
- Ryan, M. J, « New Biggest T-rex Skull. ». Consulté le 2006-04-12
- (fr)Le tyrannosaure Rex souffrait-il de goutte? sur European Vegetarian Union, 1997
- (en) JR Horner et K Padian, « Age and growth dynamics of Tyrannosaurus rex », dans Proceedings. Biological sciences / the Royal Society, vol. 271, no 1551, septembre 2004, p. 1875–80 [texte intégral (page consultée le 16 mars 2009)]
- (en) Andrew H. Lee et Sarah Werning, « Sexual maturity in growing dinosaurs does not fit reptilian growth models », dans Proceedings of the National Academy of Sciences, vol. 105, no 2, 2008, p. 582–587 [lien DOI]
- (en) Andrew H. Lee et Sarah Werning, « Sexual maturity in growing dinosaurs does not fit reptilian growth models », dans Proc Natl Acad Sci U S A, vol. 105, no 2, 15 janvier 2008 Jan 15, p. 582-7 [texte intégral (page consultée le 21 mars 2009)]
- Maturité sexuelle précoce chez les dinosaures sur http://www.radio-canada.ca, lundi 14 janvier 2008. Consulté le 17 mars 2009
- (en) MH Schweitzer, JL Wittmeyer et JR Horner, « Gender-specific reproductive tissue in ratites and Tyrannosaurus rex », dans Science (New York, N.Y.), vol. 308, no 5727, juin 2005, p. 1456–60 [texte intégral, lien DOI (pages consultées le 16 mars 2009)]
- Rapid Growth, Early Maturity Meant Teen Pregnancy For Dinosaurs sur http://www.sciencedaily.com, ScienceDaily, 15 janvier 2008. Consulté le 17 mars 2009
- (en) GM Erickson, PJ Currie, BD Inouye et AA Winn, « Tyrannosaur life tables: an example of nonavian dinosaur population biology », dans Science (New York, N.Y.), vol. 313, no 5784, juillet 2006, p. 213–7 [texte intégral, lien DOI (pages consultées le 16 mars 2009)]
- (en) Kenneth Carpenter, Dinosaur Systematics: Approaches and Perspectives, Cambridge, Kenneth Carpenter et Philip John Currie, 1992 (réimpr. Cambridge University Press), 141–145 p. (ISBN 0-521-43810-1), « Variation in Tyrannosaurus rex »
- Larson, P.L. 1994. Tyrannosaurus sex. In: Rosenberg, G.D. & Wolberg, D.L. Dino Fest. The Paleontological Society Special Publications. 7: 139–155.
- Romer, A. S. 1956. Osteology of the Reptiles. University of Chicago, Chicago, Illinois, 800 p.
- (en) « Androgynous rex – The utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs », dans Zoology (Jena), vol. 108, no 4, 2005, p. 277-86 [résumé (page consultée le 21 mars 2009)]
- All about Sue - Important Discoveries sur http://www.aucklandmuseum.com, 2009. Consulté le 13 avril 2009
- « Do egg-laying crocodilian (Alligator mississippiensis) archosaurs form medullary bone? », dans Bone, vol. 40, no 4, avril 2007, p. 1152–8 [texte intégral, lien PMID, lien DOI (pages consultées le 13 avril 2009)]
- (en) Joseph Leidy, « Memoir on the extinct reptiles of the Cretaceous formations of the United States », dans Smithsonian Contributions to Knowledge, vol. 14, 1865, p. 1–135
- Tyrannosaurus, American Museum of Natural History. Consulté le 15 avril 2009
- 1970 Newman, « Stance and gait in the flesh-eating Tyrannosaurus », dans Biological Journal of the Linnean Society, vol. 2, 1970, p. 119–123 [résumé (page consultée le 15 avril 2009)]
- L. M. Lambe, « On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon », dans Ottawa Naturalist, vol. 27, 1914, p. 129–135
- (en) Kenneth Carpenter et Matt Smith, Mesozoic vertebrate life, Bloomington, Indiana University Press, 2001 (ISBN 0-253-33907-3), p. 90–116
- (en) , Tyrannosaurus rex, the Tyrant King (Life of the Past), Bloomington, Indiana University Press, 2008 (ISBN 978-0-253-35087-9), « The extreme lifestyles and habits of the gigantic tyrannosaurid superpredators of the Late Cretaceous of North America and Asia », p. 316
- « Basal tyrannosauroids from China and evidence for protofeathers in tyrannosauroids », dans Nature, vol. 431, no 7009, 2004-10-07, p. 680–684 [lien PMID, lien DOI (pages consultées le 2008-10-07)]
- Robert T. Bakker, « The superiority of dinosaurs », dans Discovery, vol. 3, no 2, 1968, p. 11–12 [texte intégral [PDF] (page consultée le 2008-10-07)]
- Robert T. Bakker, « Anatomical and ecological evidence of endothermy in dinosaurs », dans Nature, vol. 238, 1972, p. 81–85 [texte intégral [PDF], lien DOI (pages consultées le 2008-10-07)]
- Reese E. Barrick, « Thermophysiology of Tyrannosaurus rex: Evidence from Oxygen Isotopes », dans Science, New York, vol. 265, no 5169, juillet 1994, p. 222–224 [texte intégral, lien PMID, lien DOI (pages consultées le 2008-10-07)]
- Clive Trueman, « Diagenetic effects on the oxygen isotope composition of bones of dinosaurs and other vertebrates recovered from terrestrial and marine sediments », dans Journal of the Geological Society, vol. 160, no 6, 2003, p. 895 [lien DOI]
- Reese E. Barrick, « Thermophysiology and biology of Giganotosaurus: comparison with Tyrannosaurus », dans Palaeontologia Electronica, vol. 2, no 2, octobre 1999 [texte intégral (page consultée le 2008-10-07)]
- (en) Reese E. Barrick et Michael K. Stoskopf et William J. Showers, The Complete Dinosaur, Bloomington, Indiana University Press, 1999 (ISBN 0-253-21313-4), « Oxygen isotopes in dinosaur bones », p. 474–490
- (en) Frank V. Paladino et James R. Spotila et Peter Dodson, The Complete Dinosaur, Bloomington, Indiana University Press, 1999 (ISBN 0-253-21313-4), « A blueprint for giants: modeling the physiology of large dinosaurs », p. 491–504
- (en) Anusuya Chinsamy et Willem J. Hillenius, The dinosauria, Berkeley, University of California Press, 2004 (ISBN 0-520-24209-2), « Physiology of nonavian dinosaurs », p. 643–659
- JR Hutchinson JR, V Ng-Thow-Hing V et FC Anderson, « A 3D interactive method for estimating body segmental parameters in animals: application to the turning and running performance of Tyrannosaurus rex », dans Journal of theoretical biology, vol. 246, no 4, juin 2007, p. 660–80 [lien DOI]
- David R. Carrier, « Influence of rotational inertia on turning performance of theropod dinosaurs: clues from humans with increased rotational inertia », dans Journal of Experimental Biology, Company of Biologists, vol. 204, no 22, 2001-11-15, p. 3917–3926 [texte intégral, lien PMID]
- Hutchinson, J.R., « Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa », dans Journal of Morphology, vol. 262, no 1, 2004, p. 441–461 [texte intégral [PDF], lien PMID, lien DOI]
- Hutchinson JR, Garcia M, « Tyrannosaurus was not a fast runner », dans Nature, vol. 415, no 6875, février 2002, p. 1018–21 [lien PMID, lien DOI]
- Holtz, « Phylogenetic taxonomy of the Coelurosauria (Dinosauria; Theropoda) », dans Journal of Paleontology, vol. 70, no 3, 1996-05-01, p. 536–538 [texte intégral (page consultée le 2008-10-03)]
- Christiansen, P., « Strength indicator values of theropod long bones, with comments on limb proportions and cursorial potential », dans Gaia, vol. 15, 1998, p. 241–255 (ISSN 0871-5424) [texte intégral [PDF]]
- Giraffe, WildlifeSafari.info. Consulté le 2006-04-29
- The History of Woodland Park Zoo – Chapter 4. Consulté le 2006-04-29
- Alexander, R.M., « Dinosaur biomechanics », dans Proc Biol Sci., The Royal Society, vol. 273, no 1596, August 7, 2006, p. 1849–1855 [lien PMID, lien DOI]
- Rebecca R. Hanna, « Multiple injury and infection in a sub-adult theropod dinosaur (Allosaurus fragilis) with comparisons to allosaur pathology in the Cleveland-Lloyd dinosaur quarry collection », dans Journal of Vertebrate Paleontology, vol. 22, no 1, 2002, p. 76–90 [lien DOI] catalogs the injuries of the Allosaurus known as "Big Al" – at least one was attributed to a fall.
- Hutchinson, J.R., « Biomechanical Modeling and Sensitivity Analysis of Bipedal Running Ability. II. Extinct Taxa », dans Journal of Morphology, vol. 262, no 1, 2004, p. 441–461 [texte intégral [PDF], lien PMID, lien DOI]
- Sellers, W.I., and Manning, P.L., « Estimating dinosaur maximum running speeds using evolutionary robotics », dans Proc. R. Soc. B, The Royal Society, vol. 274, no 1626, juillet 2007, p. 2711 [texte intégral, lien PMID, lien DOI]
- T. rex 'would outrun footballer', BBCNews, 2007-08-21. Consulté le 2008-10-16
- (en) Manning P, Tyrannosaurus rex, the Tyrant King (Life of the Past), Bloomington, Indiana University Press, 2008 (ISBN 0-253-35087-5), « T. rex speed trap », p. 205–228
- Paul, G.S. et Christiansen, P., « Forelimb posture in neoceratopsian dinosaurs: implications for gait and locomotion », dans Paleobiology, vol. 26, no 3, septembre 2000, p. 450 [texte intégral, lien DOI]
- MG Lockley, « A track of the giant theropod dinosaur Tyrannosaurus from close to the Cretaceous/Tertiary boundary, northern New Mexico », dans Ichnos, vol. 3, 1994, p. 213–218 [lien DOI]
- T.rex footprint discovered, Natural History Museum, 2007. Consulté le 2008-12-09
- Lambe, L. B., « The Cretaceous theropodous dinosaur Gorgosaurus », dans Memoirs of the Geological Survey of Canada, vol. 100, 1917, p. 1–84
- Farlow, J. O. and Holtz, T. R., « The Fossil Record of Predation », 2002
- Horner, J.R., « Steak knives, beady eyes, and tiny little arms (a portrait of Tyrannosaurus as a scavenger) », dans The Paleontological Society Special Publication, vol. 7, 1994, p. 157–164
- Amos, J., « T. rex goes on trial », BBC, 2003-07-31
- The king of killers or fearsome freeloader? It is one of the big questions currently in palaeontology.
- T. Rex brain study reveals a refined "nose", Calgary Herald, 2008-10-28. Consulté le 2008-10-29
- Chin, K., Erickson, G.M. et al., « A king-sized theropod coprolite », dans Nature, vol. 393, 1998-06-18, p. 680 [texte intégral, lien DOI] Summary at Monastersky, R., « Getting the scoop from the poop of T. rex », dans Science News, vol. 153, no 25, 1998-06-20, p. 391 [texte intégral, lien DOI]
- (en) Walters, Martin, Bloomsbury Illustrated Dictionary of Prehistoric Life (Bloomsbury Illustrated Dictionaries), Godfrey Cave Associates Ltd, 1995 (ISBN 978-1-85471-648-4)
- (fr)Comment les dinosaures voyaient-ils le monde ?
- The Teeth of the Tyrannosaurs, Scientific American, September 1999. Consulté le 16 mai 2010
- John R. Horner, The Complete T. Rex: How Stunning New Discoveries Are Changing our Understanding of the World's Most Famous Dinosaur, 1993, pp. 214-215.
- Catherine Vincent, « Pour les paléontologues, la science n'a pas de prix », dans Le Monde, 21 mars 2007 [texte intégral (page consultée le 13 février 2009)] :
« Le 4 octobre 1997, chez Sotheby's, à New York, eut lieu une vente historique : un Tyrannosaurus rex, nommé Sue en l'honneur de sa découvreuse, fut adjugé à 8 362 500 dollars au terme d'une bataille ayant opposé neuf enchérisseurs. »
- (en)Terry Wentz, « Sotheby’s Auction Brings A Record $8.36 Million for Tyrannosaurus rex "Sue" » sur Full Context, Black Hills Institute of Geological Research, 15. Consulté le 13 février 2009
- (en)Sean Alfano, « T-Rex Skull Auctioned For $276,000 » sur CBS NEWS, 26. Consulté le 2007-12-06
 Portail de la paléontologie
Portail de la paléontologie Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant Portail des dinosaures
Portail des dinosaures Portail de la zoologie
Portail de la zoologie
Catégories :- Carnosaure
- Dinosaure d'Amérique du Nord
- Dinosaure du Crétacé









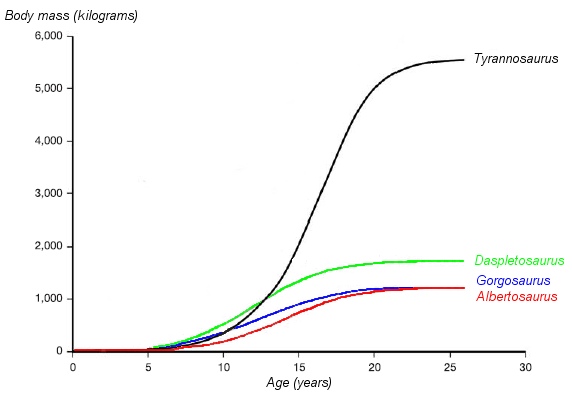



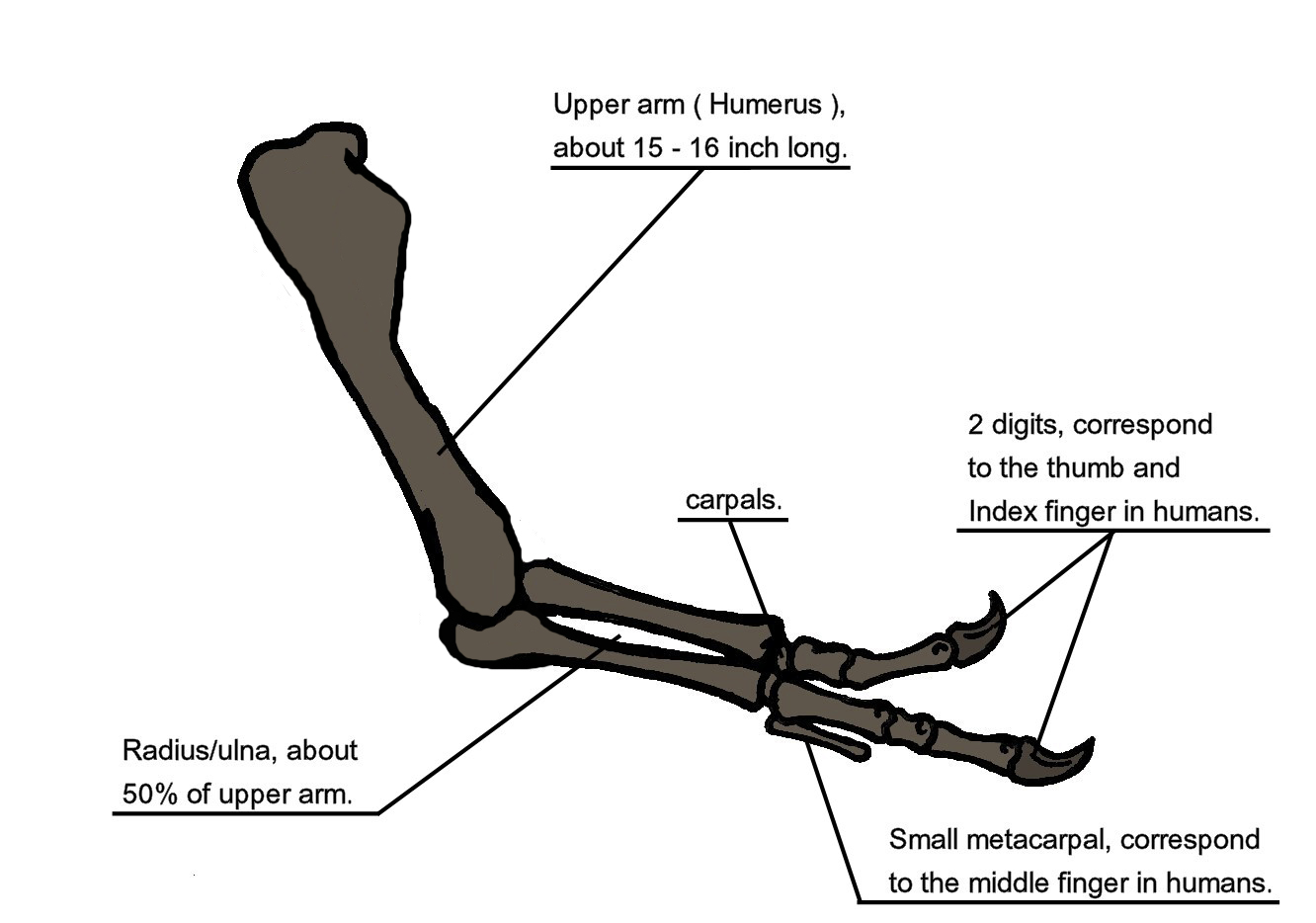

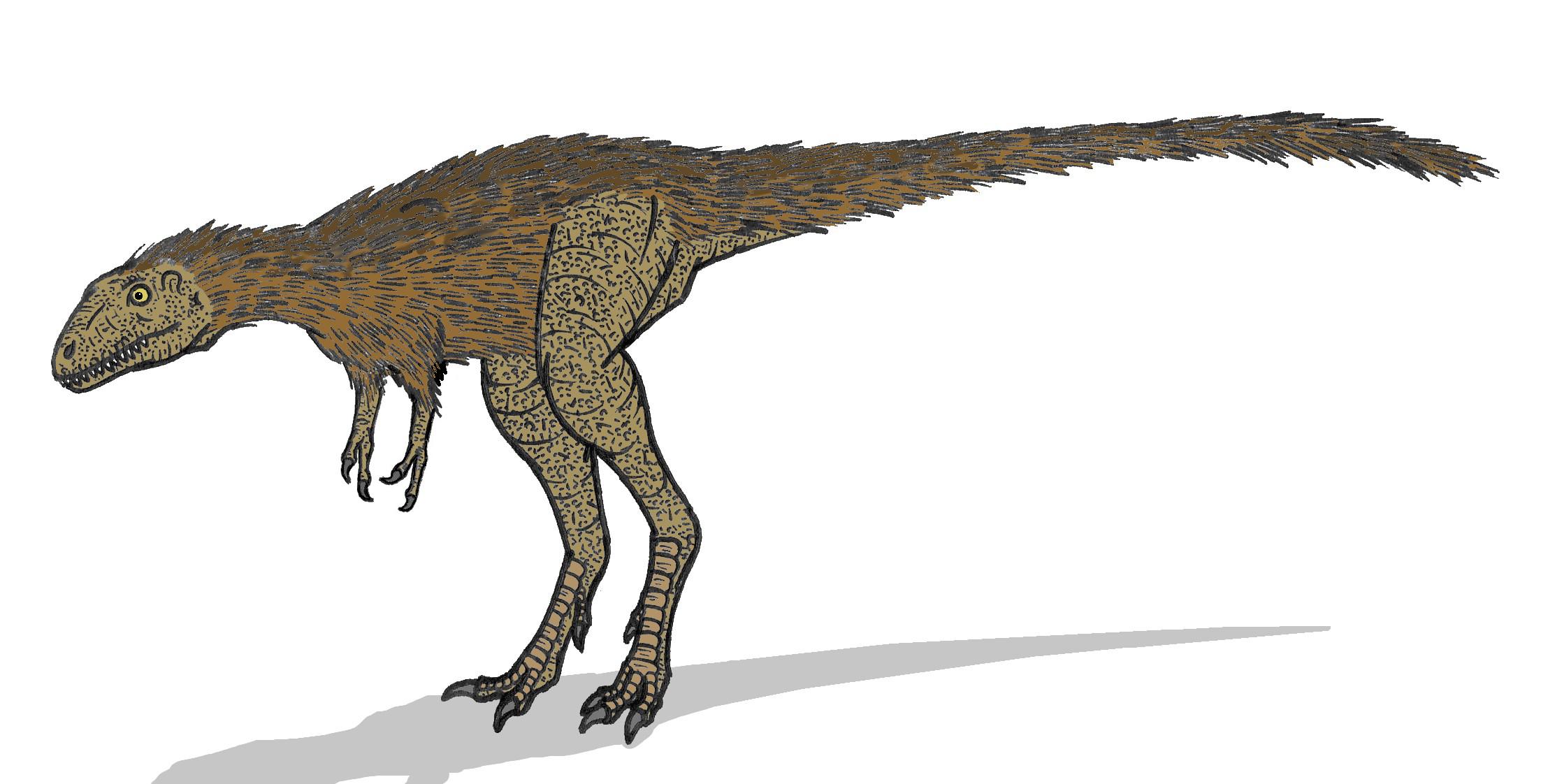







Wikimedia Foundation. 2010.