- Apparition de la vie
-
Origine de la vie
Cet article est consacré aux origines de la vie d'un point de vue scientifique. Les aspects mythiques et religieux sont traités dans l'article Cosmogonie. La précédente théorie scientifique de l'origine de la vie est traitée dans l'article génération spontanée.
Les origines de la vie sur Terre demeurent incertaines. Cependant, de nombreuses théories scientifiques existent pour expliquer l'apparition de la vie, telle que nous la connaissons aujourd'hui, dont on pense qu'elle remonte à environ 3,5 à 3,8 milliards d'années.
Cet article traite des événements antérieurs à l'apparition des trois grandes lignées du vivant.
 Stromatolites du précambrien, dans la formation de Siyeh. En 2002, William Schopf a affirmé que ces formations étaient âgées de 3,5 milliards d'années, elles seraient alors les plus anciennes traces de vie sur Terre.
Stromatolites du précambrien, dans la formation de Siyeh. En 2002, William Schopf a affirmé que ces formations étaient âgées de 3,5 milliards d'années, elles seraient alors les plus anciennes traces de vie sur Terre.

L'apparition de la vie
John Maynard Smith et Eörs Szathmáry définissent huit transitions majeures de l'évolution, dont trois concernent l'apparition de la vie :
- transition depuis des molécules autoréplicantes vers une population de molécules dans un compartiment isolé ;
- passage de réplicateurs indépendants aux chromosomes ;
- transition d'un monde à ARN — où ce dernier joue le rôle de génome et d'enzyme — à un monde à ADN et protéines.
John Desmond Bernal, quant à lui, suggère les trois étapes suivantes :
- apparition de monomères organiques,
- transitions vers des polymères organiques,
- évolution depuis des molécules vers la cellule.
De fait, il n'existe pas de modèle « standard » pour décrire l'origine de la vie. Cependant le modèle le plus couramment accepté est fondé sur l'enchaînement supposé des événements suivants :
- Des conditions prébiotiques plausibles entraînent la création de molécules organiques simples qui sont les briques de base du vivant.
- Des phospholipides forment spontanément des doubles couches qui sont la structure de base des membranes cellulaires.
- Les mécanismes qui produisent aléatoirement des molécules d'ARN (acide ribonucléique), en mesure d'agir comme des ARN-enzymes capables, dans certaines conditions très particulières, de se dupliquer. C'est une première forme de génome, et nous sommes alors en présence de protocellules.
- Les ARN-enzymes sont progressivement remplacées par des protéines-enzymes, grâce à l'apparition des ribozymes, ceux-ci étant capables de réaliser la synthèse des protéines.
- L'ADN apparaît et remplace l'ARN dans le rôle de support du génome, dans le même temps les ribozymes sont complétés par des protéines, formant les ribosomes. C'est l'apparition de l'organisation actuelle des organismes vivants.
Les étapes 2 et 3 sont parfois inversées, l'isolement en compartiment étant alors présenté après l'apparition des ARN autoréplicants.
L'origine des molécules organiques
Les plus anciennes traces de molécules organiques ont été retrouvées en 2006, dans des fossiles de crinoïdes. Âgés de 350 millions d'années, il s'agit de composés s'apparentant à des pigments, découverts par Christina O'Malley et ses collègues de l'Université d'Ohio[1].
Une explication : la condensation sur surfaces minérales
 Cristaux de pyrite
Cristaux de pyriteL'assemblage de petites molécules (comme les acides aminés) en macromolécules (comme les protéines) nécessite l'élimination de molécules d'eau. Or, la thermodynamique indique qu'il est défavorable de réaliser une telle condensation dans l'eau elle-même. Il est possible pour résoudre cette contradiction de faire appel à des surfaces minérales, comme les argiles ou les pyrites. L'adsorption des petites molécules sur ces surfaces les concentre et les modifie chimiquement, ce qui peut rendre la formation de macromolécules plus favorable.
L'argile, par exemple, se trouve très abondamment sur Terre et est constituée d'un empilement de couches fines. Entre les différentes couches de l'argile peuvent se glisser certaines petites molécules organiques, ce qui permet une adsorption importante. L'argile est aussi un catalyseur très efficace pour de nombreuses réactions organiques, et aurait donc pu permettre la polymérisation des acides aminés et/ou des acides nucléiques. Le chimiste anglais Cairns-Smith a développé cette hypothèse dans Seven clues to the origin of life en 1985 (traduction française : L'énigme de la vie, 1990).
L'expérience Urey-Miller et l'origine des molécules organiques
En 1953, Stanley Miller, accompagné de Harold Urey, a voulu reproduire les conditions de la Terre primitive. Ils ont enfermé dans un ballon des gaz (méthane CH4, ammoniac NH3, hydrogène H2 et eau H2O) et soumis le mélange à des décharges électriques pendant sept jours.
Ils ont obtenu des molécules organiques, les briques du vivant, et notamment de l'urée (CON 2H 4), du formaldéhyde (H 2CO), de l'acide cyanhydrique (HCN), des bases et des acides aminés (AA). Certains composés étant présents à plus de 2%.
Miller et Urey ont utilisé une atmosphère réductrice, méthane CH4, NH3, H2, H2O et non pas une atmosphère oxydante. Depuis l'expérience a été refaite plusieurs fois, en variant la composition de l'atmosphère et la source d'énergie (utilisation du rayonnement ultraviolet notamment). Cependant, une atmosphère moins réductrice (dioxyde de carbone CO2, azote N2, eau H2O) qui provient du volcanisme donne de très mauvais rendements.
L'exploitation de l'idée de Miller
Suite aux expériences de Miller, il a fallu déterminer les réactions chimiques qui se sont produites dans l'enceinte (le ballon dans lequel il avait enfermé les différents gaz). Ainsi est née la chimie organique dans l'eau.
Ces réactions nécessitent de fortes concentrations, des domaines de température et de pH très étroits qui font que ces mécanismes sont très peu probables : une mare en voie d'assèchement pourrait peut-être expliquer les fortes concentrations.
L'expérience fut à l'époque très critiquée à cause de cela. De plus, sa fiabilité a été remise en cause car les molécules organiques obtenues pourraient a priori provenir d'une contamination extérieure. Une contamination extérieure est toutefois improbable pour les raisons évoquées ci-après.
L'expérience de Powner-Sutherland et l'origine des nucléotides à base pyrimidique
En 2009, Sutherland et son équipe[2] sont parvenu à établir une voie de synthèse des nucléotides à base pyrimidique, l'acide uridylique et l'acide cytidylique. Les chercheurs butaient sur cette voie de synthèse pré-biotique depuis 40 ans en raison de la difficulté de trouver la bonne façon de lier le ribose à la base azotée. La clé de cette voie est de passer par un précurseur commun au ribose et la base azotée. Ce précurseur, le 2-aminooxazol, est obtenu à partir de molécules organiques élémentaires : le glycéraldéhyde, le cyanimide, le cyanoacétaldéhyde, le cyanoacétylène et le phosphate inorganique. Le mélange réactionnel alimenté par de l'azote gazeux est soumis à un cycle de chauffage-refroidissement afin de simuler le cycle d'évaporation d'une mare par le rayonnement solaire et l'alimentation par la pluie. Après une semaine le 2-aminooxazol s'accumule dans le réacteur. Le précurseur se transforme ensuite en ribose et cytosine liés ensemble. Le phosphate est ensuite ajouté au milieu réactionnel en présence d'UV durant trois jours (l'absence d'ozone dans les conditions pré-biotiques engendrait un bombardement intense d'UV). L'acide cytidylique se synthétise sous l'effet du rayonnement UV et quelques nucléotides portant une cytosine se transforme en acide uridylique. Pour les deux autres nucléotides, l'équipe de Sutherland travaillent sur un précurseur commun aux acides nucléiques à base purique.
Asymétrie des biomolécules
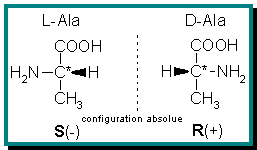
Les molécules « chirales » sont des molécules pouvant exister sous deux formes possibles : lévogyre (gauche) et dextrogyre (droite), de la même façon que la main gauche et la main droite sont l'image symétrique l'une de l'autre. On appelle énantiomères ces différentes formes.
Une analyse poussée des molécules obtenues dans l'expérience de Miller montre que l'on obtient un mélange racémique de molécules (autant de formes droites que de gauches), alors que l'on sait depuis le milieu du XIXe siècle (notamment avec des travaux de Pasteur en 1847) que les acides aminés naturels n'existent pratiquement que sous une de leurs deux formes énantiomères (on parle alors d'homochiralité) : la forme lévogyre.
On a cependant retrouvé des traces d'acides aminés de dextrogyre sous forme libre, dans des peptides ou même des protéines. Ces formes, peu fréquentes, auraient par ailleurs des fonctions physiologiques. Dans le monde vivant, on constate cependant que les sucres présents dans l'ADN sont uniquement de type dextrogyre, ou que les agents de saveur ont un goût différent selon leur forme.
Théories sur l'origine de l'homochiralité
Il existe deux grandes catégories de théories expliquant l'homochiralité : les théories biotiques et les théories abiotiques.
Dans la première, on postule que la vie serait apparue à partir d'un mélange d'énantiomères, et que l'homochiralité ne serait apparue que progressivement au cours de l'évolution. Selon Laurent Nahon, ce type de théorie n'est cependant plus beaucoup soutenu, du fait que l'on a découvert que les protéines ne peuvent se replier correctement si les acides aminés qui les composent ne sont pas chiraux. On estimerait donc que l'homochiralité serait plutôt antérieure à l'apparition de la vie, ce sont les théories abiotiques.
Cristaux et énantiomères
Les propriétés des cristaux permettent d'imaginer un scénario : certains cristaux inorganiques exposent des faces possédant une chiralité intrinsèque, comme la calcite ou le quartz. Ces faces pourraient adsorber préférentiellement une des deux formes énantiomères et donc la concentrer sélectivement aux dépens de l'autre.[3] Cependant, rien n'indique que les cristaux aient pu jouer un rôle dans la formation de molécules organiques.
Asymétricité et thermodynamique
Il a été découvert que la matière est intrinsèquement asymétrique. « Lorsque l'on place des atomes de cobalt dans une géométrie asymétrique, c'est-à-dire dans des champs magnétiques, les électrons produits de la désintégration de neutron se déplacent toujours dans la direction opposée à leur spin (aligné sur le champ). Les électrons sont donc intrinsèquement gauches. » Cette expérience de Tsung Dao Lee et Chen Ning Yang qui reçurent le prix Nobel en 1957 peut être reproduite avec n'importe quel atome. Un gaz de vapeur de césium par exemple dans un champ électromagnétique a un pouvoir rotatoire. C'est ce sur quoi a travaillé Marie-Anne Bouchiat, directrice de recherche CNRS à l'ENS. Ce phénomène est facilement observable sur les atomes lourds car la force mise en jeu est la force d'interaction faible entre le noyau et l'électron.
Ainsi il a été calculé que les acides aminés naturels sont thermodynamiquement plus stables que leur image dans un miroir.
Formation d'énantiomères dans l'espace
 Une météorite.
Une météorite.Dans les années 1970, des acides aminés ont été découverts dans la météorite de Murchison, or, ils étaient présents majoritairement sous leur forme lévogyre. On a ainsi découvert 70 acides aminés différents, dont 3 seulement font partie des 20 acides aminés naturels. Le pourcentage exact de molécules lévogyres est cependant beaucoup discuté, du fait d'éventuelles contaminations, et varie entre 50% et -5% suivant les équipes de recherche.
L'idée que l'homochiralité aurait pour origine des molécules venues de l'espace s'est donc développée.
Les énantiomères absorbent différemment la lumière lorsque celle-ci est polarisée « circulairement » droite ou gauche. Or, ces molécules se dégradent après absorption, conduisant donc à un excès d'une forme énantiomérique.
 La nébuleuse d'Orion.
La nébuleuse d'Orion.On a découvert, en 1997, que la nébuleuse d'Orion produit de la lumière polarisée circulaire à 17% dans l'infrarouge (IR). L'infrarouge n'a pas assez d'énergie pour casser des liaisons covalentes, mais on peut supposer que les ultraviolets (UV) sont également polarisés circulairement.
Une expérience menée en 2005, a montré qu'un mélange racémique d’un acide aminé simple irradié par un rayonnement UV conduit à un mélange homochiral. Dans cette expérience, la leucine est utilisée à l'état solide, reproduisant les conditions spatiales. Après une irradiation par un rayonnement synchrotron polarisé circulairement droit (proche de la longueur d'onde observée dans l'espace, dans l'UV lointain), l'expérience permet d'obtenir un excès de l'énantiomère lévogyre de 2.6%.
Or, il suffit d'un excès de 1% dans des réactions qui s'entretiennent pour conduire à un mélange homochiral de 100%. D'après Laurent Nahon, aucune expérience liée à des théories concurrentes n'est parvenue à un tel excès.
Des molécules organiques aux protocellules
 Schéma d'une cellule animale.
Schéma d'une cellule animale.Aujourd'hui, de nombreux modèles résolvent le problème de l'apparition des molécules organiques. Les scientifiques arrivent à produire de nombreuses petites molécules biologiques (acides aminés, sucres, bases nucléiques) dans des conditions prébiotiques en laboratoires.
Les expériences de Miller et les modèles qui en sont dérivés ne fournissent pas d'explication sur les étapes suivantes qui incluent la transition des monomères aux biopolymères, puis aux protocellules et finalement aux cellules vivantes ayant un métabolisme de base. Aussi les scientifiques ont exploré d'autres voies de recherche.
Compartiments isolés
L'apparition de compartiments isolés par une membrane pose de sérieux problèmes. Les membranes des cellules vivantes sont composées de lipides, or on connaît aujourd'hui des acides gras à longue chaîne qui peuvent spontanément former des petites membranes sphériques. Bien que l'on puisse produire de tels compartiments en laboratoires, ces acides gras restent synthétisés par des enzymes. Le processus permettant de former de tels compartiments en l'absence de ces enzymes demeure inconnu.
Protocellules
Un compartiment isolé par une membrane ne forme cependant pas une protocellule. Selon Maynard Smith, deux conditions sont nécessaires pour former une véritable protocellule :
- Les molécules capables de répliquer la forme de base (les réplicateurs) doivent se lier entre elles en un « chromosome », formant ainsi une unité structurelle, garantissant aux réplicateurs de former un tout cohérent après la réplication ;
- la membrane doit posséder des mécanismes d'échange avec le milieu extérieur, autres que les systèmes à protéines actuels.
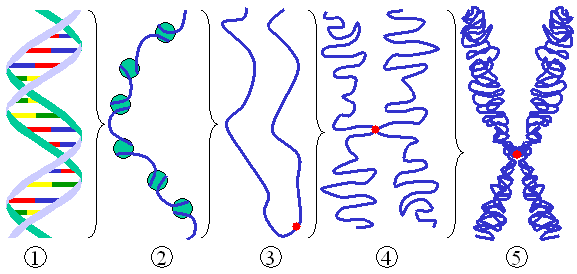
 Schéma général d'un chromosome eucaryote.
Schéma général d'un chromosome eucaryote. Schéma d'une membrane semi-perméable.
Schéma d'une membrane semi-perméable.L'apparition du génome
Les modèles « gènes d'abord »
 Une molécule d'ADN.
Une molécule d'ADN.Dans ce modèle, l'apparition du génome a précédé l'apparition du métabolisme. Des molécules d'ADN ou d'ARN auraient ainsi existé seules, s'autorépliquant à partir des molécules présentes dans leur environnement. Les « individus » sont donc représentés par les molécules d'acides nucléiques elles-mêmes.
Les modèles « métabolisme d'abord »
Plusieurs modèles rejettent l'idée de l'autoréplication d'un gène « nu » et font l'hypothèse de l'apparition d'un métabolisme primitif qui aurait précédé l'émergence de la réplication de l'ARN. Une des premières versions de cette hypothèse fut présentée en 1924 par Alexander Oparin avec son idée de vésicules primitives capables de se répliquer, à une époque où on ne connaissait pas encore la structure de l'ADN.
D'autres variantes sont apparues dans les années 80 et 90 comme la théorie de Günter Wächtershäuser sur un monde sulfuro-ferreux, ou les modèles de Christian de Duve basés sur la chimie des thioesters.
D'autres arguments plus abstraits ont aussi été présentés. On peut citer les modèles mathématiques de Freeman Dyson au début des années 1980 sur la probabilité de l'émergence d'un métabolisme sans présence de gènes, ou encore les travaux de Stuart Kauffman sur les ensembles globalement autocatalytiques (voir génération spontanée pour une présentation des idées de Kauffman sur l'origine de la vie).
Métabolisme et génome : un monde à ARN
L'hypothèse du monde à acide ribonucléique (ARN) est que l'ARN était la principale — et sans doute la seule — forme de vie avant l'émergence de la première cellule à ADN. C'est Walter Gilbert qui a utilisé pour la première fois le terme « monde à ARN » (« RNA world » en anglais) en 1986.
L'hypothèse d'un monde à ARN a aujourd'hui la faveur des scientifiques et est fondée sur plusieurs éléments. Notamment sur le fait que l'ARN est en théorie capable aussi bien d'assurer des tâches métaboliques que d'être le support d'une information génétique.
Stockage et réplication
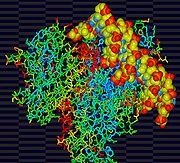 Une protéine liée à un brin d'ADN.
Une protéine liée à un brin d'ADN.L'ARN a la capacité de stocker une information, en utilisant un code génétique similaire à celui de l'ADN. L'ARN peut également se comporter comme un ribozyme (de la contraction de ribose et enzyme) et catalyser certaines réactions, tout comme les protéines. Du point de vue de la reproduction, cette molécule possède donc deux fonctions primordiales : le stockage de l'information et la catalyse nécessaire à l'autoréplication.
L'ADN peut aussi se recopier lui-même, mais seulement avec l'assistance de protéines. Les protéines sont de très bons catalyseurs mais elles sont incapables de stocker l'information requise pour leur propre réplication. L'ARN est lui capable à la fois de catalyse et d'autoréplication. Ainsi, le ribosome est un ribozyme, dans le sens où le responsable de la synthèse des protéines n'est pas une protéine (comme c'est le cas dans la grande majorité des catalyses d'une cellule vivante) mais l'ARN ribosomique lui-même. Ces ribozymes peuvent se replier dans l'espace, faisant apparaître un site actif pour une catalyse, à l'instar des protéines.
L'ADN, formant une double hélice rigide, ne peut se replier pour jouer un rôle de catalyseur.
Efficacité des protéines
 La structure « quaternaire » d'une protéine.
La structure « quaternaire » d'une protéine.Les protéines sont des catalyseurs très efficaces, bien plus que les ribozymes. De même, il existe 20 acides aminés dans le monde vivant, mais seulement quatre nucléotides, les protéines sont donc bien plus diversifiées que les ARN.
D'un point de vue évolutif, il est donc peu probable qu'une protéine-enzyme ait été remplacée par une ARN-enzyme. À l'inverse, si les ARN sont bien apparus avant les protéines, il est plausible qu'ils aient été remplacés par des protéines, plus efficaces.
Cet argument est étayé par le fait que l'ARN joue un rôle dans la synthèse des protéines, via son rôle fondamental dans les ribosomes actuels. L'ARN aurait donc en quelque sorte conduit à l'apparition des protéines.
Les protéines utilisées dans la structure du ribosome, seraient donc venues plus tard : les premières protéines auraient été sélectionnées selon leur capacité d'amélioration du fonctionnement des ribozymes, pour finalement se substituer à eux.
Une grande distribution phylogénétique
Les ARN sont présents dans les trois lignées du monde vivant (archées, procaryotes, eucaryotes). Ils accomplissent dans chacun un grand nombre de tâches différentes, les plus connus sont l'ARN messager (ARNm, véhiculant l'information génétique de l'ADN vers les ribosomes), l'ARN de transfert (ARNt, faisant le lien entre acide nucléique et acide aminé) et l'ARN ribosomique (ARNr, composant structuraux et fonctionnels des ribosomes). À coté de ceux-ci, on peut trouver un grand nombre d'ARN impliqué dans des fonctions tels que des catalyses, des régulations de l'expression de gènes, des contrôles, des défenses antivirales, des extinctions de gènes, des inhibitions de synthèses de protéines, des restaurations génomiques, etc. C'est le cas des ARN interférents (ARNs), dont certains chercheurs qualifient le mécanisme d'« universel ». Les ARNtm des procaryotes, ont également plusieurs fonctions : ils jouent à la fois les rôles d'ARN de transfert et d'ARN messager.
Malgré cette grande diversité structurelle et fonctionnelle, la répartition des ARN permet de retrouver le découpage du vivant. Ainsi, les petits ARN nucléolaires ne sont partagés que par les archées et les eucaryotes, l'ARN de la télomérase n'est lui présent que chez les eucaryotes alors que les procaryotes sont les seules à posséder des ARNtm. De la même façon, les trois grands types d'ARN (ARNt, ARNm et ARNr) sont présents dans les trois lignées.
Le cas de l'ARN de transfert
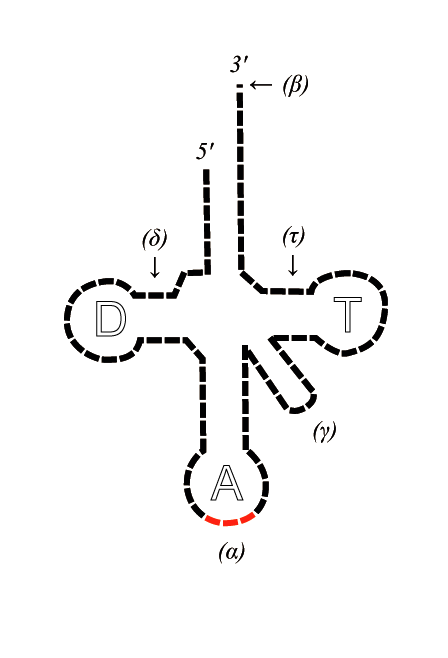
 La structure de l'ARN de transfert.
La structure de l'ARN de transfert.Le rôle de l'ARNt est de transporter un acide aminé vers le ribosome, où s'effectuera la liaison avec un autre acide aminé, pour former un polypeptide (donnant ainsi une protéine). Il existe plusieurs ARNt, possédant chacun trois nucléotides : l'anticodon. L'anticodon correspond à un codon, porté par l'ARNm qui définit l'ordre d'assemblage des acides aminés par le ribosome.
La particularité de l'ARNt est qu'il est, malgré sa petite taille, en partie constitué de nombreux nucléotides que l'on ne rencontre pas ailleurs. Ces nucléotides « exotiques » auraient ainsi une origine prébiotique, vestiges d'un monde à ARN. On retrouve ainsi ces composants dans l'ensemble des trois domaines du vivant.
Virus et ARNt
Il est relativement fréquent d'observer des virus à ARN ou des viroïdes portant des motifs similaires à de l'ARNt. Ainsi, dans le viroïde PSTV (Potato Spindle Tuber Viroïd) et l'ARNt de la tyrosine, on trouve les mêmes motifs structuraux en « feuille de trèfle ». Pour Marie-Christine Maurel, « ces derniers jouent un rôle fondamental dans le vivant et leur ancienneté ne fait pas de doute ».
Autre structure surprenante : chez le virus TYMV (Turnip Yellow Mosaic Virus), l'amorce de la traduction du génome du virus en protéine se fait par le biais d'une structure de type ARNt qui amorce sa propre traduction et qui fixe un acide aminé.
 Structure du virus PSTV.
Structure du virus PSTV.Évolution dirigée d'ARN
En 1990, Larry Gold et Jack Szostak ont mis au point une méthode visant à diriger l'évolution d'ARN, afin de sélectionner ceux montrant une activité catalytique. Ils ont depuis réussi à obtenir des ribozymes capables de lier des nucléotides entre eux, de lier des acides aminés à des ARNs, d'effectuer des réactions d'oxydo-réductions, de se lier à des composants de la membrane, etc.
Il est donc en théorie possible, sur ce modèle, que l'ARN seul suffise à établir un métabolisme primitif. Toutefois, il reste encore à découvrir un ARN capable de se répliquer lui-même.
ARN et hérédité
L'ARN joue un rôle dans la transmission de l'activité des gènes : un tel mécanisme (qualifié d'épigénétique) n'est pas lié à l'ADN, et serait une preuve de la capacité de l'ARN à participer à l'hérédité.
À l'origine de l'ADN dans la cellule
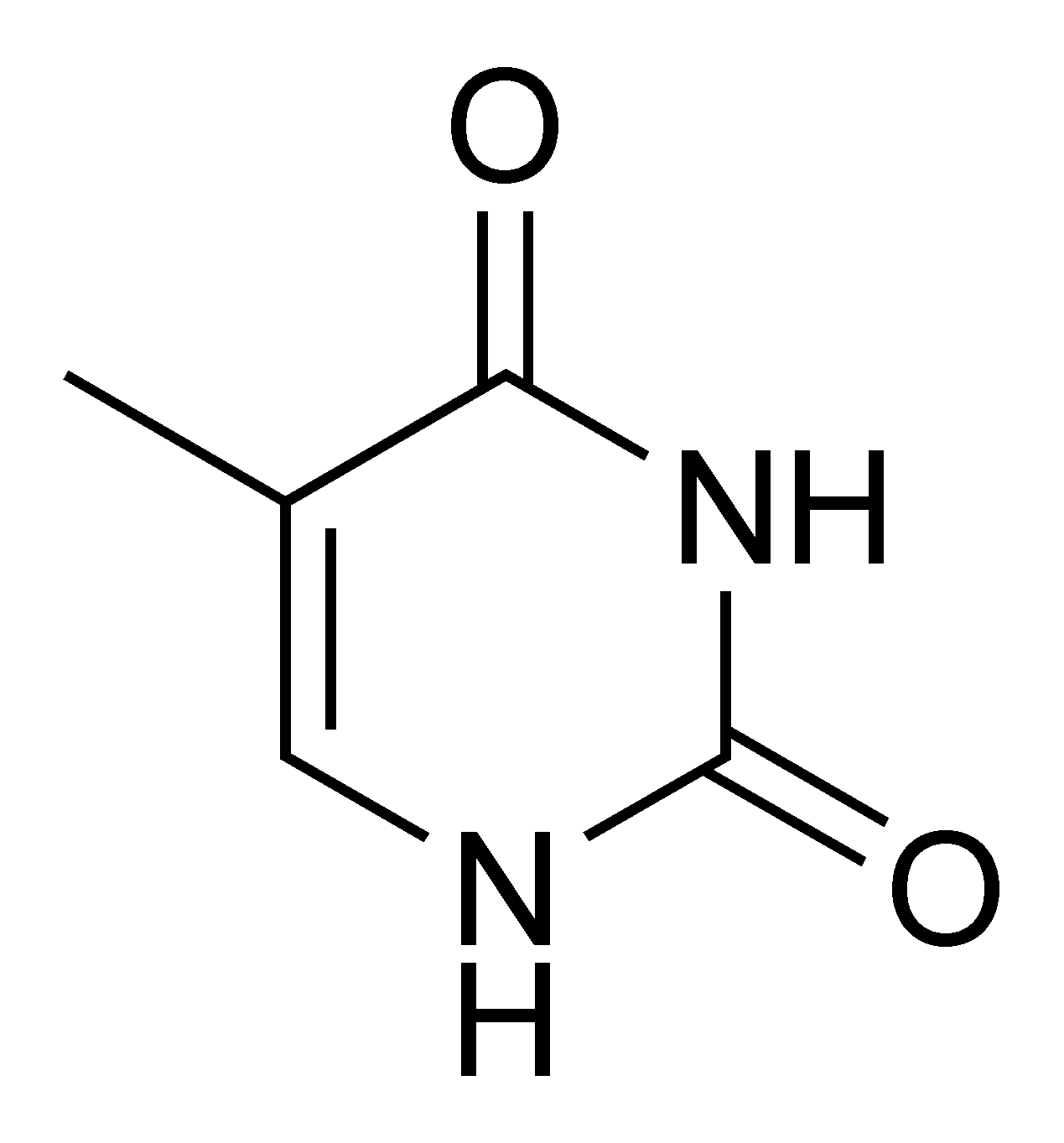
Du point de vue de la biologie cellulaire, l'ADN est produit par modification d'un ARN : les désoxyribonucléotides (précurseurs de l'ADN) sont en effet produits à partir des ribonucléotides (précurseurs des ARN). De plus, le groupement thymine (identifié comme T dans le code génétique), est construit à partir d'un groupement uracile (U). Or, bien que l'uracile soit spécifique à l'ARN et la thymine à l'ADN, le groupement U est, au moment de la synthèse, déjà fixé sur un désoxyribonucléotide.
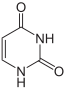 Uracile.
Uracile. Thymine.
Thymine.De l'ARN à l'ADN
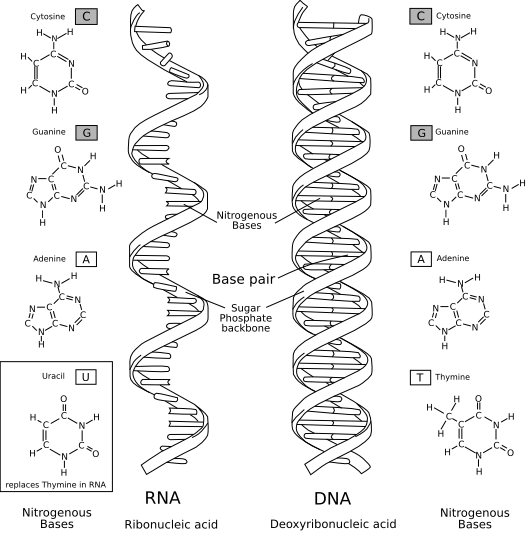
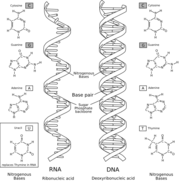 Comparaison entre une molécule d'ARN (à gauche) et une molécule d'ADN (à droite).
Comparaison entre une molécule d'ARN (à gauche) et une molécule d'ADN (à droite).Dans l'hypothèse du monde à ARN, apparaissent tout d'abord des viroïdes ressemblant à des ARN auto-catalytiques, présents dans des compartiments isolés (qu'ils soient membranaires ou cristallins). Puis des protocellules, capables de métabolisme archaïque, sont soumises à une évolution darwinienne, évoluant ainsi vers des cellules à ARN, capables de présenter une activité variée et complexe.
Ribozymes ou protéines ?
Bien que l'ARN soit donc à l'origine de l'ADN dans le métabolisme cellulaire, cette réaction est très difficile à réaliser. De fait, dans les trois lignées, elle est catalysée par des protéines spécialisées : les ribonucléotides-réductases. De plus, cette réaction est très coûteuse en énergie, du fait de la réduction du ribose, et elle produit des radicaux libres, très réactifs, sur la protéine. L'ARN étant une molécule fragile, il paraît improbable qu'elle puisse supporter des radicaux libres sans l'intervention de protéines.
Ainsi, l'origine de l'ADN trouve vraisemblablement sa source après l'apparition des protéines, indispensables à chaque étape de sa synthèse à partir de précurseurs de type ARN, au sein de la cellule.
Intérêt de l'ADN
 Structure chimique de l'ADN.
Structure chimique de l'ADN.L'ADN présente un certain nombre d'avantages sur l'ARN, en termes de conservation de l'intégrité de l'information génétique.
Tout d'abord il se casse moins facilement, car le désoxyribose de l'ADN contient un atome d'oxygène de moins que le ribose de l'ARN. Or l'oxygène peut facilement interagir sur les liaisons entre nucléotides, posant alors un problème de stabilité.
Ensuite, l'ADN permet la réparation d'un problème récurrent : la transformation du groupement thymine (T) en uracile (U). En effet, l'uracile, uniquement présent dans l'ARN, est une anomalie dans l'ADN.
Avantage sélectif de l'ADN : l'hypothèse du virus
 Structure de base d'un virus.
Structure de base d'un virus.Les avantages en termes de stabilité de l'ADN pourraient ne pas suffire à expliquer son adoption. Ainsi, Patrick Forterre avance l'hypothèse qu'un avantage sélectif supplémentaire peut être dû aux conflits entre virus et cellules vivantes.
Dans ce modèle, le premier organisme à ADN serait un virus. L'ADN conférerait au virus le pouvoir de résister à des enzymes dégradant les génomes à ARN, arme de défense probable des cellules. On retrouve le même principe chez des virus actuels, qui altèrent leur ADN pour résister à des enzymes produites par des bactéries infectées.
Actuellement, on peut observer que les enzymes nécessaires à la traduction de l'ARN vers l'ADN sont très présentes chez les rétrovirus, dont le génome est porté par de l'ARN. De la même façon, de nombreux virus codent leurs propres enzymes de synthèse de l'ADN.
Cette hypothèse est également corroborée par la découverte de virus à ADN, dont celui-ci contient, non pas des groupements thymines, mais des groupements uraciles. Du point de vue évolutif, il y aurait donc eu d'abord apparition des désoxyribonucléotides, puis de l'ADN à uracile (ADN-U), puis d'ADN à thymine (ADN-T), qui se serait progressivement imposé. D'après Patrick Forterre, il est même probable que l'ADN-T ait été « inventé deux fois », chez des virus différents.
Les virus à ARN seraient ici des reliques du monde à ARN, les virus à ADN-U seraient alors des reliques du monde ayant précédé celui à ADN-T.
Les virus, premiers organismes à ADN
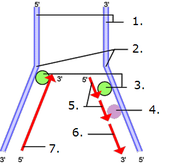 Schéma de la réplication de l'ADN chez les eucaryotes.
Schéma de la réplication de l'ADN chez les eucaryotes.Les virus à ADN pourraient être plus anciens que la première cellule à ADN : la première cellule à ADN l'aurait donc emprunté à un ou plusieurs virus, sous la pression d'une course aux armements (théorie de la reine rouge).
Didier Raoult et Jean-Michel Claverie ont ainsi découvert le mimivirus : un virus géant à ADN (son génome étant deux fois plus long que le plus petit génome bactérien connu). La particularité de ce virus est qu'il peut produire des protéines impliquées dans la traduction de l'ARN en protéines (comme des enzymes chargeant des acides aminés sur des ARNt), il pourrait donc avoir pour ancêtres des virus plus anciens que la première cellule à ADN.
Eugène Koonin et ses collègues ont mis en avant, en comparant des génomes séquencés, que la plupart des enzymes impliqués dans la réplication de l'ADN sont différents entre les eubactéries et les eucaryotes (accompagnés des archées). Ils en concluent que l'ADN aurait été inventé indépendamment dans la lignée des eubactéries et celle conduisant aux eucaryotes et aux archées.[réf. nécessaire]
De même, les enzymes de réplications des virus à ADN sont très différentes d'un virus à l'autre, ainsi que par rapport aux enzymes cellulaires jouant le même rôle.
Ces indices laissent penser que les enzymes liées à l'ADN sont apparues au cours d'un « premier âge » du monde à ADN, où existaient cellules à ARN et virus à ARN et à ADN.
Passage de l'ADN dans les cellules
La nature du génome du plus ancien ancêtre commun à tous les êtres vivants (que les scientifiques prénomment LUCA) reste inconnue : faisait-il encore partie du monde à ARN, ou avait-il déjà un génome à ADN ? Quoi qu'il en soit, LUCA est le fruit d'une longue évolution. Le génome des premières cellules, qui ont précédé LUCA, était sans doute constitué par des molécules d'ARN et non pas d'ADN. Contrairement à l'ADN, l'ARN peut en effet jouer à la fois le rôle d'enzyme et de matériel génétique. Stanley Miller et Christian de Duve pensent que l'apparition de l'ARN a été elle-même un évènement tardif, en effet, cette molécule ne semble pas pouvoir être synthétisée par les méthodes simples de la chimie prébiotique. L'ARN aurait donc été précédé par des molécules dont nous ne connaîtrons sans doute jamais la nature exacte.
Ce qui amène les scientifiques à penser que l'ARN a précédé l'ADN résulte de ce constat : les trois grandes lignées du vivant ne partagent que le système de synthèse des protéines, alors qu'elles diffèrent sur le système de réplication de l'ADN.
Il existe alors plusieurs hypothèses pour le passage de l'ADN (d'origine viral) dans les cellules vivantes : soit ce passage s'est produit une seule fois, soit il a pu avoir lieu plusieurs fois, indépendamment dans les différentes lignées. Dans le premier cas, les premiers gènes des enzymes de réplications auraient donc été remplacés par la suite par ceux d'un autre virus, formant ainsi les trois lignées.
L'hypothèse de Patrick Forterre est que les trois lignées du vivant trouvent leurs sources dans le remplacement du génome à ARN par le génome à ADN de trois virus différents. On retrouve notamment ici l'hypothèse d'une origine virale du noyau des eucaryotes.
Les travaux de Carl Woese semblent appuyer cette hypothèse, en démontrant que la vitesse d'évolution des protéines semble avoir chuté au moment de l'apparition des trois lignées. Cette diminution serait due au passage de l'ARN à l'ADN, les génomes à ADN étant plus stables, et donc moins sensibles aux mutations.
Dans ce scénario, l'apparition de trois lignées uniquement s'explique par le fait que les cellules à ADN ont peu à peu supplanté les cellules à ARN, empêchant ainsi l'apparition de nouvelles lignées par passage ARN→ADN.
Autres modèles
Sources hydrothermales : le monde du soufre
Les sources hydrothermales ont été découvertes en 1977 à 2600 mètres de profondeur, là où deux plaques tectoniques se séparent.
Les monts hydrothermaux sont situés sur la couche sédimentaire. Leur diamètre à la base varie de 25 à 100 mètres et leur hauteur varie de 70 à 100 mètres. Les cheminées de ces fumeurs sont parfois recouvertes d'une croûte d'oxyde de manganèse. Les fumeurs situés sur ces monts sont composés d'un solide friable dont la couleur varie du gris noir à l'ocre, ce sont des sulfures de fer, de cuivre et de zinc.
Ces sources sont particulièrement intéressantes car on y a trouvé la vie où on la croyait impossible : milieu privé d'oxygène, à haute température, chargé de métaux et de soufre, dans l'obscurité la plus totale. Cependant les gradients de température importants autour de ces zones et le fait que les ultraviolets destructeurs ne parviennent pas si profondément (alors qu'ils détruisent toute molécule formée à la surface) sont de bonnes conditions pour l'apparition de la vie.
Ces organismes ont les mêmes formes que ceux que l'on connaît plus près de la surface (ADN, protéines, sucres...) mais puisent leur énergie de l'oxydation de H2S pour transformer le carbone minéral en matière organique.
D'autre part, des expériences ont été menées, au laboratoire de géophysique de Washington, et ont montré que dans les conditions qui existent autour des évents, il y a formation de NH3, forme réduite de l'azote qui est tant nécessaire à la formation des molécules organiques de la première partie et qui n'existait pas dans l'atmosphère oxydante. Les sources hydrothermales sont donc de bonnes sources de NH3.
Une origine extraterrestre primitive (exogenèse)
Une hypothèse alternative est que la vie se soit d'abord formée hors de la Terre. Les composés organiques sont relativement fréquents dans l'espace, notamment dans les zones lointaines du système solaire où l'évaporation des composés volatils est très réduite. Certaines comètes sont enrobées dans des couches de matière sombre, qu'on pense être une sorte de bitume formé par une combinaison de composés carbonés simples et de rayons ultraviolets. La pluie de matériaux cométaires sur la Terre primitive pourrait avoir apporté des quantités de molécules organiques complexes, ce qui aurait causé l'apparition de la vie sur Terre.
Une hypothèse plus large est la panspermie : la vie même serait apparue dans l'espace puis disséminée sur Terre. Selon une variante, la vie serait apparue sur Mars d'abord et des impacts de comètes et d'astéroïdes sur Mars auraient projeté du matériel de la surface martienne sur Terre. Il est encore plus difficile de trouver des indices pour justifier ces hypothèses que les théories plus classiques.
Ces théories d'une origine extraterrestre n'expliquent pas directement comment la vie est apparue, car elles ne font a priori que reporter le problème. Cependant, elles élargissent les conditions dans lesquelles la vie a pu apparaître dans l'univers. Les futurs échantillons de sols ramenés de Mars et de comètes permettront peut-être d'obtenir de nouveaux éléments de réponse.
Voir aussi
- Biologie, Biologie cellulaire, vie
- Biochimies hypothétiques
- Exobiologie
- Habitabilité d'une planète
- Génération spontanée
- Protéinoïde
Sources
- (en) John Desmond Bernal, The origin of life, éditions Weidenfeld & Nicolson, 1967.
- (fr) Guillemette Lauters, Olivier Lenaerts, Daniel Rousselet et Eric Depiereux, Bioscope : L’origine de la vie.
- (fr) John Maynard Smith, Eörs Szathmáry, Les Origines de la vie, éditions Dunod, 2000. ISBN 2100048600
- (en) J. William Schopf, Anatoliy B. Kudryavtsev, David G. Agresti, Thomas J. Wdowiak, Andrew D. Czaja, « Laser−Raman imagery of Earth's earliest fossils », dans Nature, volume 416, pages 73–76, mars 2002 [(en) lire en ligne].
- (fr) Marie-Christine Maurel, La Naissance de la vie : de l'évolution prébiotique à l'évolution biologique, éditions Dunod, 2003 (3e édition). ISBN 2100068822
- (en) N. Fujii, T. Saito, « Homochirality and life », dans The Chemical Records, volume 4, no 5, pages 267–78, 2004 [(en) lire en ligne]
- (fr) Uwe Meierhenrich, Un pas vers l’origine de l’homochiralité…, novembre 2005.
- (fr) Patrick Forterre, « L'origine du génome », dans Les dossiers de La Recherche : l'histoire de la vie, les grandes étapes de l'évolution, no 19, pages 34–40, mai 2005.
- (fr) Marc-André Selosse, « Quelle parenté entre les trois grandes lignées du vivant ? », dans Les dossiers de La Recherche : l'histoire de la vie, les grandes étapes de l'évolution, no 19, pages 42–45, mai 2005.
- (fr) « Laurent Nahon : « L'asymétrie des biomolécules vient de l'espace » », propos recueillis par Franck Daninos, dans La Recherche, no 390, octobre 2005.
- (en) U. Meierhenrich, L. Nahon, C. Alcaraz, J. Bredehöft, S. Hoffmann, B. Barbier, A. Brack, « Asymmetric VUV photodecomposition of the amino acid D,L-Leucine in the solid state », dans Angewandte Chemie International Edition, volume 44, pages 5630–5634, juillet 2005 [(en) lire en ligne].
Bibliographie
- (fr) Le site de la Société Française d'Exobiologie (SFE).
- (fr) Conférence de Patrick Forterre : "Regards croisés sur les origines de la vie" (2009) à l'ENS Cachan.
- (en) Uwe Meierhenrich: Amino acids and the asymmetry of life, Springer-Verlag, 2008. ISBN 978-3-540-76885-2
- (en) Site dédié au mimivirus.
- (en) Anthony M. Poole, What is the Last Universal Common Ancestor ?, septembre 2002.
- (en) Antonio Lazcano The Origins of Life. Have too many cooks spoiled the prebiotic soup?, Natural History magazine, février 2006.
- (en) Erwin Schrödinger, What is Life?, 1944. (fr) Traduction française : Qu'est-ce que la vie?, ed. Seuil.
- (fr) Joël de Rosnay, Les Origines de la vie, éditions du Seuil, 1966. ISBN 2020002167
- (fr) André Brack, Bénédicte Leclercq, La vie est-elle universelle ? – Des premiers êtres vivants à l'exploration spatiale, éditions EDP Sciences, 2003. ISBN 2-86883-674-7.
- (fr) Sous la direction de Muriel Gargaud, Didier Despois, Jean-Paul Parisot, L'Environnement de la Terre Primitive, Collection : L'origine de la Vie sur Terre et dans l'Univers, Presses universitaires de Bordeaux, 2001. ISBN 2-86781-267-4.
Liens externes
- (fr) Comprendre facilement l'origine de la vie sur la Terre
- (fr) Un site qui explique avec des mots simples les origines de la vie
- (fr) Les trois grandes théories de l'apparition de la vie sur Terre
Domaines généraux de la biologie Anatomie | Biochimie | Bio-informatique | Biologie cellulaire | Biologie de l'évolution | Biologie humaine | Biologie marine | Biologie moléculaire | Botanique | Écologie | Exobiologie | Génétique | Géobiologie | Microbiologie | Origines de la vie | Paléontologie | Parasitologie | Physiologie | Taxinomie | Zoologie Notes et références
- ↑ O'Malley, Christina E., Ausich, William I., and Chin, Yu Ping, Biomarkers in colored fossil crinoids: implications for phylogeny, Geological Society of America Annual Meeting, Geological Society of America Abstracts with Programs, Vol. 38, No. 7, p. 482, octobre 2006. résumé de la présentation, article de de vulgarisation
- ↑ Matthew W. Powner, Béatrice Gerland and John D. Sutherland, Synthesis of activated pyrimidine ribonucleotides in prebiotically plausible conditions, Nature, Vol 459, 14 May 2009, pp. 239-242
- ↑ Robert M. Hazen, Timothy R. Filley, and Glenn A. Goodfriend, PNAS May 8, 2001 vol. 98 no. 10 5487–5490Y
 Portail de la chimie
Portail de la chimie Portail de la biochimie
Portail de la biochimie Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant
Catégories : Origine de la vie | Histoire évolutive | Métabolisme








Wikimedia Foundation. 2010.