- Photosynthèse
-
 La feuille est l’organe spécialisé dans la photosynthèse chez les plantes vertes.
La feuille est l’organe spécialisé dans la photosynthèse chez les plantes vertes.
La photosynthèse (grec φῶς phōs, lumière et σύνθεσις sýnthesis, composition) est le processus bioénergétique qui permet aux plantes et à certaines bactéries de synthétiser de la matière organique en exploitant la lumière du soleil. Les besoins nutritifs de ces organismes sont du dioxyde de carbone, de l’eau et des sels minéraux. La photosynthèse est à la base de l'autotrophie de ces organismes. La photosynthèse est la principale voie de transformation du carbone minéral en carbone organique. En tout, les organismes photosynthétiques assimilent environ 100 milliards de tonnes de carbone en biomasse, chaque année[1].
La photosynthèse se déroule dans les membranes des thylakoïdes, chez les plantes, les algues et les cyanobactéries, ou dans la membrane plasmique chez les bactéries photosynthétiques. Une conséquence importante est la libération de molécules de dioxygène.
Sommaire
La découverte du mécanisme
- Dans l'Antiquité, Aristote pensait que le sol fournissait aux plantes les éléments dont elles ont besoin.
- Au XVIIe siècle, Jan Baptist van Helmont démontre qu'un saule planté en bac a pris 77 kg en 5 ans alors que le sol contenu dans le bac ne diminue que de 57 g ; il attribue la différence à l'action de l'eau.
- Au XVIIIe siècle, plusieurs scientifiques mettent en évidence les notions de respiration et de production d’oxygène par les plantes et l'importance de la lumière dans ce dernier phénomène. Ce sont d’abord deux chimistes anglais : Stephen Hales en 1727, qui pense que l'air et la lumière contribuent à la croissance des plantes, et Joseph Priestley entre 1771 et 1777 qui met en évidence le rejet d’oxygène. À leur suite, Jan Ingen-Housz, médecin et botaniste hollandais, établit en 1779 le rôle de la lumière dans la production d’oxygène par les plantes. Puis Jean Senebier, un pasteur suisse, à partir des travaux d’Antoine Lavoisier sur la composition de l'air, comprend que les plantes consomment du dioxyde de carbone et rejettent du dioxygène lors de cette phase.
- Au début du XIXe siècle, Nicolas Théodore de Saussure démontre la consommation d’eau lors de la photosynthèse. La chlorophylle est isolée par des chimistes français en 1817, Pierre Joseph Pelletier et Joseph Bienaimé Caventou.
- Au milieu du XIXe siècle les grandes lignes du mécanisme sont comprises, transformation de l'énergie lumineuse, consommation d’eau et de dioxyde de carbone, production d’amidon et rejet de dioxygène. L'expérience cruciale d’Engelmann (1882), où des bactéries servent d’indicateur de la production d’oxygène, montre clairement le rôle de la couleur de la lumière.
- C'est au cours du XXe siècle que l'explication plus détaillée du processus s’établit. Le début du siècle voit la description de la structure chimique de la chlorophylle puis la découverte de l'existence des types a et b. Robert Emerson établit en 1932[2] que 2500 molécules de chlorophylle sont nécessaires pour émettre 1 molécule d'O2. Dans les années 1930, les travaux de Robert Hill permettent d'y voir plus clair. À l'issue de ses expériences, la photosynthèse se présente comme une réaction d'oxydo-réduction au cours de laquelle le carbone passe d'une forme oxydée à une forme réduite : CO2 → HCHO ; et l'oxygène d'une forme réduite à une forme oxydée : H2O → O2
- On a récemment découvert du nanoplancton qui vit à des profondeurs où il n'y a pratiquement plus de lumière. Certaines algues marines (cryptophytes, dont rhodomonas sp. et chroomonas sp.), à température ambiante et en condition d'illumination défavorable, savent, grâce à des « antennes » protéiques, et semble-t-il grâce à des protéines (bilines) utilisées en plus de la chlorophylle, mieux capter la lumière et utiliser la cohérence quantique pour optimiser leur utilisation du rayonnement solaire incident. Cela leur permet de conduire plus de 95 % des photons jusqu'à leur « but »[3]. Ce phénomène pourrait exister chez d'autres végétaux, voire être courant.
Vue générale
La photosynthèse est l’un des plus anciens processus biogéochimiques de la terre. On a retrouvé par exemple des microfossiles d’organismes similaires à des bactéries, vieux de 3,8 milliards d’années, probablement capables de réaliser la photosynthèse.
C'est aussi le plus important. Elle fournit la quasi totalité de la matière organique et de l’énergie nécessaires à l’existence des écosystèmes de la planète, les autres mécanismes de chimiosynthèse, comme la chimiolithotrophie, (et les organismes qui en dépendent), étant marginaux.
Toutes les plantes terrestres (exceptées certaines plantes parasites comme les Orobanches) et toutes les algues ainsi que certaines bactéries se servent de la photosynthèse.
La photosynthèse comprend trois phases :
- De l’énergie électromagnétique, sous forme de lumière, est absorbée grâce à l’action de pigments dont le plus connu est la chlorophylle.
- Cette énergie est transformée en énergie chimique, sous forme de potentiel d'oxydo-réduction.
- L’énergie chimique est utilisée pour réduire le CO2 et incorporer le carbone dans des composés organiques riches en énergie qui permettent grâce à l’anabolisme (synthèse organique) la croissance des êtres vivants et grâce au métabolisme (transformation) énergétique un apport en énergie[Note 1].
Les premiers éléments ainsi fabriqués, via le cycle de Calvin, sont des sucres (glucose). Ce processus est représenté par l'équation suivante :
6CO2 + 12H2O + lumière → C6H12O6 + 6O2 + 6H2O. On rencontre également souvent, cette équation sous la forme d'un simple bilan de matière, ce qui masque le fait que les atomes du dioxygène produit ne proviennent que de l’eau :
6CO2 + 6H2O + lumière → C6H12O6 + 6O2 Remarque : certains organismes photosynthétiques (des bactéries) ne produisent pas d'O2 et le CO2 n’est pas l'unique source de carbone. Ces molécules d'O2 et de CO2 ne sont donc pas les dénominateurs communs de la photosynthèse. Il serait alors plus rigoureux de définir la photosynthèse comme étant « une série de processus dans lesquels l’énergie électromagnétique est convertie en énergie chimique utilisée pour la biosynthèse du matériel cellulaire » comme le propose Gest (2002)[4].
La photosynthèse peut se faire dans les bactéries (chez les cyanobactéries), certains protistes, et dans les chloroplastes des plantes et des algues. Un seul animal est connu pour être le siège d'un processus de photosynthèse, cependant il ne produit pas lui-même les chloroplastes où elle se déroule, il les trouve dans les algues dont il se nourrit.
Le support de la photosynthèse
Articles détaillés : Chloroplaste et Photosystème.La photosynthèse se déroule dans les membranes des thylakoïdes, très riches en protéines et pigments ; les plus connus étant les chlorophylles ; ces pigments (par exemple : chlorophylle a,b et caroténoïdes) présentent différents spectres d'absorption.
Chez les eucaryotes photosynthètiques, la photosynthèse se déroule dans un organite spécifique : le chloroplaste issu de l'endosymbiose d'une cyanobactérie il y a environ un milliard et demi d'années.
Dans la cellule, au sein des membranes photosynthétiques, des « antennes collectrices de lumière » (décrite au début des années 2000, à une résolution quasi-atomique grâce à la cristallographie-rayons X) augmentent et régulent la section efficace de capture de l'énergie lumineuse. Elles permettent d'orienter l'énergie lumineuse vers d'autres protéines membranaires : les centres réactionnels, qui transformeront cette énergie lumineuse en énergie chimique.L'ensemble des antennes collectrices et des centres réactionnels est appelé « photosystème ». Les photosystèmes interviennent dans la première phase de la photosynthèse en capturant les premiers électrons initiant la réaction photochimique.
Quand la lumière est « trop » intense, un mécanisme de protection (non photochimique) se met en place, évacuant l’excès d’énergie lumineuse sous forme de chaleur ou de fluorescence (quenching non photochimique) pour limiter le stress oxydatif destructeur que causerait sans cela la formation d'espèces réactives de l'oxygène (ROS). D'autres mécanismes permettent également d'éviter la formation de ces ROS en cas de luminosité trop importante : le cycle des xanthophylles, le cycle eau-eau, la chlororespiration et possiblement la photorespiration dont on ignore le but véritable. On a d'abord cru que l’antenne collectrice de lumière LHCII changeait de conformation, mais des travaux récents (spectroscopie) laissent penser qu'elle est rigide, et orientent vers un nouveau mécanisme (peut-être une inactivation par interaction avec une autre protéine) [Note 2],[5].
Les deux phases de la photosynthèse
Si la photosynthèse peut s’étudier de manière globale avec :
6CO2 + 12H2O + lumière → C6H12O6 + 6O2 + 6H2O. Ce processus se déroule en réalité en deux phases bien distinctes :
1. Les réactions photochimiques, appelées communément « phase claire », qui peuvent se résumer ainsi :
12H2O + lumière → 6O2 + énergie chimique (24 Hydrogènes) . 2. Le cycle de Calvin, appelé aussi phase de fixation du carbone ou phase non-photochimique, ou encore improprement appelé « phase sombre » :
6CO2 + énergie chimique (24 Hydrogènes) → C6H12O6 + 6H2O Ce qui est noté « énergie chimique » correspond à 12 molécules de NADPH+H+ et de l'ATP. On aura remarqué que la 2de phase utilise l'énergie chimique fournie par la 1re phase photochimique. La 2de phase dépend aussi de la lumière, mais indirectement. C'est pourquoi l'expression « phase sombre » souvent utilisée dans le passé, est en fait inappropriée.
Les réactions photochimiques ou phase claire
La lumière nous parvient sous forme de photons. Ces photons possèdent un potentiel énergétique différent selon leur longueur d'onde. L’énergie transportée par un photon est inversement proportionnelle à la longueur d’onde. Un photon de lumière rouge possède moins d’énergie qu’un photon de lumière bleue.
Les pigments absorbent mieux certaines longueurs d’onde. Par exemple, la chlorophylle absorbe bien la lumière rouge et la lumière bleue, mais elle n’absorbe pas bien la lumière verte ce qui leur donne cette couleur. Les caroténoïdes quant à eux absorbent mieux la lumière verte mais pas bien la lumière jaune ou la lumière orange ce qui leur donne cette couleur.
Lorsqu’un pigment capte un photon d'énergie correspondant à sa capacité d’absorption un de ses électrons passe à l’état excité. Cette énergie peut se transmettre de 3 façons : soit en la réémettant sous forme de photon lumineux, soit sous forme de chaleur ; ces deux voies ne sont pas utiles au processus photosynthétique. La troisième consiste à transmettre l’énergie par résonance et il n’y a presque aucune perte d’énergie.
Les antennes collectrices ( light harvesting complex, ou LHC) sont des ensembles comprenant des pigments (chlorophylles a et b, caroténoïdes et phycoérythrobiline) et des protéines. Elles collectent l’énergie lumineuse et la distribuent aux centres réactionnels des photosystèmes.
Les centres réactionnels sont le lieu où toute l’énergie converge. Ils sont composés de molécules de chlorophylle a (P680 ou P700) liée à un accepteur primaire d’électrons.
Les photosystèmes sont composés des antennes collectrices qui entourent un centre réactionnel et de plusieurs molécules (phéopytines, plastoquinone, ferrédoxine)servant à transporter des électrons et des protons. À l’exception de quelques transporteurs d’électrons toutes les molécules qui composent les photosystèmes sont raccordées les unes aux autres.
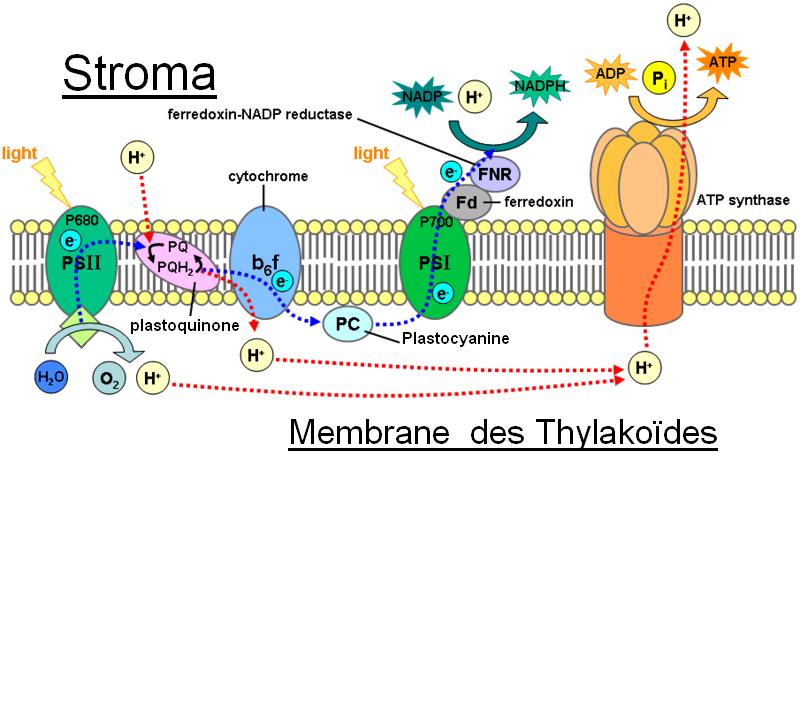
Le photosystème II
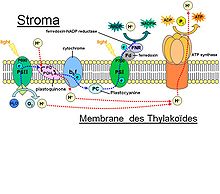 réactions de photosynthèse au niveau de la membrane thylakoide.
réactions de photosynthèse au niveau de la membrane thylakoide.Le photosystème II (appelé ainsi parce qu’il a été découvert en second) et les complexes des cytochromes sont responsables de la libération d’oxygène dans l’atmosphère et il produit de l’ATP à partir d’ADP et d’un phosphate.
- Un pigment des LHC capte un photon qui correspond à une longueur d’onde qu’il peut absorber. Un électron de ce pigment passe à l’état excité. L’énergie est transmise par résonance à un autre pigment.
- L’énergie se transmet ainsi jusqu’au centre réactionnel du PS II.
- Le P680 est une molécule de chlorophylle a située au centre du CR du PS II. Elle est reliée à une phéophytine (phéo). Le P680 capte très bien les photons d’une longueur d’onde aux alentours de 680 nm. Lorsque cette molécule reçoit l’énergie provenant des LHC ou qu’elle capte elle-même un photon, un de ses électrons passe de l’état fondamental à l’état excité. Cet électron n’a pas le temps de retourner à l’état fondamental, car il est capté par la Phéo.
- La Phéo est une molécule de chlorophylle sans atome central de magnésium. Cet atome est remplacé par 2 atomes d’hydrogène. Cette molécule capte l’électron excité du P680.
- Revenons un peu en arrière : Le P680 vient de perdre un électron, il doit en trouver un pour redevenir stable. La tyrosine Z (tyr Z) est le donneur primaire d’électron du PS II. Cette molécule va donner un électron au P680. Cette molécule possède un groupement hydroxyde. Pour demeurer stable elle perdra l’hydrogène du radical OH, cet hydrogène deviendra un proton, car son électron a été cédé au P680.
- La tyr z doit aussi redevenir stable pour que le processus se poursuive. Un agrénat de 4 manganèses est l’enzyme qui sépare l’eau (OEC)[6]. Cette molécule est située du côté sur la paroi intérieure du thylakoïde. Deux molécules d’eau y sont accrochées. La tyrosine qui a perdu son électron va s’emparer d’un atome d’hydrogène d'une molécule d'eau et elle sera de nouveau stable. Cette étape se produit 4 fois. Les deux O vont se libérer de l’agrénat et former du O2 (L’oxygène qu’on respire).
- Revenons à la Phéo qui vient de gagner un électron. Cet électron sera repris par une autre molécule qui le donnera à la plastoquinone (PQ). Elle prendra un proton provenant du stroma pour demeurer stable. Un autre électron arrivera et la PQ prendra un autre proton.
- La molécule va se diriger vers les complexes du cytochromes b6/f. Elle libérera ses protons dans le lumen et donnera ses électrons aux complexes des cytochromes.
- Conséquemment il y a beaucoup plus de protons dans le lumen que dans le stroma. La membrane est très peu perméable aux protons alors ceux-ci doivent traverser par le canal de l’ATP synthase. Le fait qu’un proton passe dans ce canal produit l’énergie nécessaire à la production d’ATP par cette enzyme.
Le photosystème I
Le photosystème I (PS I) est responsable de la libération de NADPH dans le Stroma.
- la plastocyanine (PC) prend un électron des complexes des cytochromes b6/f et l’apporte au P700.
- Le P700 est une molécule de chlorophylle a qui absorbe bien les photons d’une longueur d’onde se situant aux alentours de 700 nm. Le fonctionnement du PS I est semblable au fonctionnement du PS II : les LHC dirigent leur énergie vers le P 700 qui perd un électron au détriment d’une autre molécule. Cet électron est remplacé par un électron de la PC.
- L’électron éjecté du P700 est capté par une molécule de chlorophylle a qui le cède à une autre molécule qui le cède à une autre molécule, qui le cède à une autre molécule (le passage de l’électron d’une molécule à l’autre lui fait perdre de l’énergie), qui le cède à une autre molécule. Cette molécule cède l’électron à la ferrédoxine.
- La ferrédoxine est une molécule composée de 2 atomes de fer et de 2 atomes de soufre. Elle est située près du stroma entre les complexes des cytochromes et le PS I. Elle peut fournir des électrons à plusieurs autres métabolismes tel celui de l’azote. Dans le cas de la photosynthèse elle donne son électron à une molécule appelée ferrédoxine NADP réductase. Cette molécule unira deux protons provenant du stroma à une molécule de NADP à l’aide de l’électron qu’elle vient de recevoir. Elle se situe dans le stroma.
La photophosphorylation non cyclique est le processus expliqué ci-haut car les électrons ne reviennent jamais à la même molécule.
La photophosphorylation cyclique
La photophosphorylation cyclique intervient lorsque le taux de NADPH devient trop élevé, car il faut plus d’ATP que de NADPH.
- Le P700 du PS I devient excité, un électron est éjecté, il suit la chaîne d’électrons jusqu’à la ferrédoxine. La ferrédoxine se déplace jusqu’à la plastoquinone et un électron passe de la ferrédoxine à la plastoquinone.
- la PQ prend un proton du stroma et la dernière étape se reproduit. La PQ ayant 2 protons se dirige vers les complexes des cytochromes b6/f.
- Les électrons retournent vers le P700 par la plastocyanine.
- Les protons induits dans le lumen par PQ et les complexes des cytochromes servent à produire de l’ATP grâce à l’ATP synthase.
Une caroténoïde se trouve près du P680 et du P700. Lorsque la tyr z ou la PC ne peut pas fournir d’électron au P680 ou au P700 la caroténoïde cède un électron pour éviter que le P… détruise tout le CR en enlevant un électron à une molécule avoisinante. Les caroténoïdes peuvent répandre leur énergie sous forme de chaleur si trop d’énergie se dirige vers les CR.
Le cycle de Calvin ou phase chimique non photo-dépendante (ou « phase sombre », expression désormais désuète)
Article détaillé : Cycle de Calvin.Dans un deuxième temps, l'énergie chimique contenue dans l'ATP et le NADPH+H+ permet de fixer le carbone contenu dans le dioxyde de carbone atmosphérique en le liant aux atomes d'hydrogène des molécules d'eau. C'est le cycle de Calvin ou phase de fixation du carbone. Cette étape porte aussi le nom de phase chimique, et parfois le nom de « phase sombre », bien que pouvant se réaliser à la lumière : cette appellation reflète seulement le fait que la lumière n'est pas directement nécessaire à cette étape, contrairement à la phase photochimique (ou « phase claire »), qui est photo-dépendante.
Le carbone fixé se fait ensuite réduire en glucide par l'ajout d'électrons et de protons H+. Le potentiel réducteur est fourni par le NADPH+H+ qui a acquis des électrons grâce à la phase photochimique. Enfin, le cycle de Calvin a besoin d'énergie sous forme d'ATP pour convertir le carbone en glucide.
Cependant, chez la plupart des végétaux, le cycle de Calvin se déroule de jour car c'est durant le jour que la phase photochimique peut régénérer le NADPH+H+ et l'ATP indispensable à la transformation du carbone en glucide. Car sans la présence de la lumière et les produits qui résultent de la phase photochimique, la phase chimique n'aurait pas lieu. La phase photochimique et la phase chimique (non photo-dépendante) sont complémentaires, l'une ne va pas sans l'autre.
Les différents types de fixation du carbone
Il y a trois mécanismes connus de fixation du dioxyde de carbone au cours de la photosynthèse : C3, C4 et CAM. Ces trois mécanismes diffèrent par l'efficacité de cette étape. Le type de photosynthèse de la plante est déterminé par le nombre d'atomes de carbone de la molécule organique formée en premier lors de la fixation du CO2.
Le mécanisme en C3 correspond au mécanisme « de base », c'est celui de 98%[réf. nécessaire] des plantes vertes [Hopkins].
Les types en C4 et CAM sont plus rares, mais on les trouve dans des espèces connues : le maïs est une plante C4, l'ananas une plante CAM. Ces deux adaptations sont apparues chez de nombreux groupes de plantes, vraisemblablement (et principalement) comme adaptation au stress hydrique ou à une réduction de disponibilité de CO2 pendant la journée ; l'amélioration du rendement photosynthétique apparaît être, dans les deux cas, un sous-produit[7], et non le facteur de pression évolutive.
Le mécanisme des plantes en C3
La première des étapes du cycle de Calvin-Benson consiste en une carboxylation (fixation d'une molécule de CO2) sur le ribulose 1,5 bisphosphate, catalysée par la RubisCO, pour donner deux molécules d'un composé à 3 atomes de carbone (Acide 3-phosphoglycérique, APG). Une grande majorité des plantes, dont tous les arbres, fonctionnent selon ce mécanisme. Le CO2 fixé par la RubisCO provient de la diffusion du CO2 atmosphérique au travers des stomates dans un premier temps puis, sous forme dissoute, au travers des cellules de la feuille jusqu'au stroma des chloroplastes.
La RubisCO est capable de catalyser une réaction en utilisant l'oxygène au lieu du CO2, c'est le phénomène de photorespiration, qui diminue le taux de photosynthèse nette[Note 3] mais présente peut-être une utilité encore mal comprise[Note 4].
Le mécanisme des plantes en C4
 Métabolisme des plantes C4.
Métabolisme des plantes C4.Une des adaptations de la plante qui évite la photorespiration est d'augmenter la pression partielle en CO2 autour de la RubisCO.
Le métabolisme C4 dissocie dans l'espace, les phases photochimique et non photochimique. Les plantes qui l'utilisent captent le CO2 atmosphérique, non directement par RubisCO, mais par l'action de la phospho-énol-pyruvate-carboxylase (PEP-carboxylase) qui produit un composé à quatre atomes de carbone (un acide dicarboxylique : oxaloacétate, puis malate ou aspartate). Ces réactions ont lieu dans le mésophylle (assise cellulaire entre les nervures).
Ce composé à 4 atomes de carbone est ensuite transporté vers les cellules de la gaine périvasculaire où une enzyme se charge de le décomposer, libérant ainsi le CO2 et recyclant le transporteur. Le CO2 est donc concentré dans ces cellules et est fixé par la RubisCO, selon le mécanisme des plantes en C3 mais avec un meilleur rendement.
Ce type de photosynthèse existe notamment chez des graminées d'origine tropicale et aride, comme la canne à sucre ou le sorgho. La fermeture de leurs stomates plus longtemps permet à ces plantes de limiter les pertes d'eau, mais cela présente l'inconvénient d'abaisser rapidement la pression partielle en CO2 dans la feuille, car il est prélevé par la photosynthèse. Le métabolisme en C4 permet de compenser, et au-delà, cette baisse de pression et de maintenir le fonctionnement de la photosynthèse et même de l'améliorer. La photorespiration est nulle ou très faible chez ces plantes, du fait de l'enrichissement en CO2.
De nombreuses espèces dans différentes familles sont en C4. Opuntia ficus-indica est un exemple chez les Cactaceae. Le maïs est une plante C4, qui consomme 400 L d'eau par kg de matière sèche produite contre 1500 L pour le blé[8] ; pour autant sa consommation à l'ha est supérieure à celle du blé à cause d'un rendement plus élevé. C'est aussi une des cultures les plus irriguée dans certaines régions françaises (Landes), car les besoins en eau sont concentrés sur juillet/aout[9]. A contrario, le maïs n'est quasiment pas irrigué en Bretagne-Pays de Loire (25% des surfaces en France)[10]. Ce caractère a été hérité de la plante ancêtre, la téosinte, qui, elle, est adaptée à un milieu chaud et sec, car sa production de biomasse est moindre. Il a été maintenu par la sélection artificielle car il assure de bons rendements.
Le mécanisme des plantes CAM (Crassulacean Acid Metabolism)
Article détaillé : Métabolisme acide crassulacéen.Ces plantes possèdent les deux types d'enzymes carboxylantes comme les plantes de type C4. Elles diffèrent de ces dernières du fait que la fixation du carbone n'est pas séparée dans l'espace (mésophylle/gaine périvasculaire) mais dans le temps (nuit/jour). Durant la nuit, lorsque les stomates sont ouverts, un stock de malate est produit, puis stocké dans la vacuole des cellules photosynthétiques. Au cours de la journée, ces malates sont retransformés en dioxyde de carbone et le cycle de Calvin peut s’effectuer, le CO2 restant disponible pour la photosynthèse malgré la fermeture des stomates. Ainsi les pertes d'eau par transpiration sont limitées. Ce mécanisme est observé notamment chez les Crassulaceae (« plantes grasses », comme le cactus).
Ce mécanisme existe aussi dans des milieux aquatiques, lorsque la disponibilité en CO2 est réduite pendant la journée (par exemple du fait de la consommation par les plantes).
Cas particuliers de photosynthèse
Photosynthèse chez un animal
La limace de mer, Elysia chlorotica, est le premier animal découvert capable d'utiliser la photosynthèse pour produire de l'énergie. Cette photosynthèse est effectuée par les chloroplastes de sa principale source de nourriture, l'algue Vaucheria litorea. Ils sont séquestrés par l'épithélium digestif de la limace pendant plusieurs mois, continuant la photosynthèse. Cependant, les chloroplastes de Vaucheria litorea ont transféré une grande partie de leurs gènes nécessaires à la photosynthèse vers le noyau de cette algue au cours de l'évolution[11]. La photosynthèse est néanmoins maintenue chez la limace de mer grâce au transfert d'un gène nécessaire à la photosynthèse du noyau de l'algue vers celui de la limace[11].
Photosynthèse artificielle
La reproduction artificielle des réactions de photosynthèse fait l'objet de nombreuses recherches, qui se focalisent en général sur l'une ou l'autre des deux phases de la photosynthèse.
Ainsi, une équipe du National Renewable Energy Laboratory rapporte en 1998 avoir créé un dispositif permettant de dissocier l'oxygène et l'hydrogène de l'eau à partir d'énergie solaire[12]. Le dispositif est cependant instable et fabriqué à partir de matériaux rares. Une équipe du MIT indique en 2011 être parvenue à des résultats similaires, cette fois avec un dispositif stable et à partir de matériaux courants et bon marchés[13].
Une équipe de l'université de Kyoto annonce quant à elle en 2007 un procédé capable de capter le CO2 atmosphérique 300 fois plus efficacement que les plantes[14],[15], à partir de dioxyde de manganèse.
En mars 2011, une équipe de l'institut de technologie du Massachusetts dirigé par le Dr Daniel Nocera a annoncé avoir réussi à fabriquer une feuille artificielle, faite de matériaux durables et peu couteux, permettant de décomposer l'eau en oxygène et hydrogène, à l'aide de la lumière du soleil afin de créer de l'électricité propre[16]. Les catalyseurs utilisés par Nocera, permettant d'accélérer le processus d’électrolyse de l'eau, ont été réalisés à partir de cobalt (ou de nickel [5]), et fonctionnent à pH 7, à température ambiante et sous pression atmosphérique[17].
Production d'oxygène & captage d'énergie
À l’échelle planétaire, ce sont les algues et le phytoplancton marin qui produisent le plus d’oxygène, suivi des forêts. On a longtemps cru que les mers froides et tempérées étaient les seules à avoir un bilan positif en termes d’oxygène, mais une étude de 2009 [18]montre que les océans subtropicaux oligotrophes sont également producteurs d’oxygène, bien qu'ayant une production saisonnière irrégulière. Ces océans jouent donc un rôle en termes de puits de carbone. Pour le sud de l'hémisphère nord, la production d’oxygène est basse début d’hiver, augmente jusqu’en août pour redescendre à l'automne[réf. à confirmer] [19]. De même on a longtemps cru que l'oxygène n'était produit que dans les couches très superficielles de l’océan, alors qu'il existe également du nanoplancton, vivant généralement à grande profondeur, photosynthétique[réf. souhaitée]. Dans les zones de dystrophisation ou dans les zones mortes de la mer, ce bilan peut être négatif.
Le flux d’énergie capté par la photosynthèse (à l’échelle planétaire) est immense, approximativement 100 térawatts[20]: qui est environ de 10 fois plus élevé que la consommation énergétique mondiale (intégrée sur un an) [Note 5]. Ce qui signifie qu'environ un peu moins du millième de l’insolation reçue par la Terre est captée par la photosynthèse et fournit toute l’énergie de la biosphère.
Notes
- traduction de l'article wikipédia allemand
- Études de l’Institut Max Planck de Francfort, de l’Université de Francfort et de l’Institut de Biologie Structurale de Grenoble (Communiqué CEA)
- Ce qui n'est gênant pour la plante que si c'est la lumière qui est le facteur limitant de son développement ; la prédominance du type C3 est un indice que ce n'est généralement pas le cas.
- On pense qu'elle permet de tamponner la concentration en O2 dans la cellule, pour éviter les oxydations, ou encore qu'elle permet la synthèse d'acides aminés tels que la sérine dans la mitochondrie des plantes concernées[réf. nécessaire]
- Qui s’élève à environ 10 à 12 térawatts.
Références
- (en) Christopher B. Field, Michael J. Randerson et Paul Falkowski, « Primary production of the biosphere: integrating terrestrial and oceanic components », dans Science, vol. 281, no 5374, juillet 1998, p. 237–40 [résumé, lien PMID, lien DOI]
- Gerald Karp, Biologie Cellulaire et Moléculaire, De Boeck Université, 2004, 872 p. (ISBN 2-8041-4537-9) [lire en ligne], p. 225
- (en) Elisabetta Collini, Cathy Y. Wong, Krystyna E. Wilk, Paul M. G. Curmi, Paul Brumer et Gregory D. Scholes, « Coherently wired light-harvesting in photosynthetic marine algae at ambient temperature », dans Nature, vol. 463, 4 février 2010, p. 644-647 [résumé, lien DOI]
- (en) Howard Gest, « History of the word photosynthesis and evolution of its definition », dans Photosynthesis Research, no 73, 2002, p. 7–10 [texte intégral, lien PMID]
- (en) Tiago Barros, Antoine Royant, Jörg Standfuss, Andreas Dreuw et Werner Kühlbrandt, « Crystal structure of plant light-harvesting complex shows the active, energy-transmitting state », dans The EMBO Journal, vol. 28, 2009, p. 298-306 [texte intégral, lien DOI]
- Jack Farineau et Jean-François Morot-Gaudry, La photosynthèse: Processus physiques, moléculaires et physiologiques, Editions Quae, 2006, 412 p. (ISBN 2-7380-1209-4 et 9782738012098) [lire en ligne]
- (en) Jon E. Keeley et Philip W. Rundel, « Evolution of CAM and C4 carbon-concentrating mechanisms », dans International Journal of Plant Sciences, vol. 164, no 3, 2003, p. 555-577 SUP (ISSN 1058-5893) [résumé, texte intégral]
- http://www.snv.jussieu.fr/vie/dossiers/eau/eaugestion/eauagriculture.html L'eau et les activités agricoles
- http://www.linternaute.com/savoir/dossier/eau-virtuelle/cereales.shtml D'après les chiffres de la FAO, 5750 m3 par hectare, légèrement plus que le blé]
- http://www.agpm.com/pages/generalites.php?PHPSESSID=cc765b3e529e276608b52a126cd34563
- (en) Mary E. Rumpho, Jared M. Worful, Jungho Lee, Krishna Kannan, Mary S. Tyler, Debanish Bhattacharya, Ahmed Moustafa et James R. Manhart, « Horizontal gene transfer of the algal nuclear gene psbO to the photosynthetic sea slug Elysia chlorotica », dans PNAS, vol. 105, no 46, 18 novembre 2008, p. 17867-17871 [lien DOI]
- (en) Oscar Khaselev et John A. Turner, « A Monolithic Photovoltaic-Photoelectrochemical Device for Hydrogen Production via Water Splitting », dans Science, vol. 280, 1998, p. 425 [résumé]
- (en) Debut of the first practical “artificial leaf” sur American Chemical Society, 27 mars 2011.
- Barbara Ngouyombo, ambassade de France au Japon, « Bulletin électronique Japon 435 »
- Photosynthèse artificielle pour réduire la pollution atmosphérique sur techno-science.net
- http://www.gurumed.org/2011/03/28/photosynthse-cration-des-premires-feuilles-artificielles-fabricables/
- http://sciencetonnante.wordpress.com/2011/04/26/la-photosynthese-artificielle-de-l%E2%80%99energie-a-partir-de-soleil-et-d%E2%80%99eau/
- (en) S. Riser et K. Johnson, « Net production of oxygen in the subtropical ocean », dans Nature, no 451, 2008, p. 323-5 [texte intégral, lien DOI] (étude conduite sur 3 ans).
- « Les océans subtropicaux aussi sont producteurs d’oxygène », dans Science et Avenir, mars 2008, p. 19
- (en) Kenneth H. Nealson et Pamela G. Conrad, « Life: past, present and future », dans Philosophical Transactions of The Royal Society of London B Biological Sciences, vol. 354, no 1392, décembre 1999, p. 1923–39 [résumé, lien PMID, lien DOI]
Voir aussi
Articles connexes
Liens externes
- [flash] (en) la photosynthèse
- [flash] Le mécanisme de la photosynthèse, exemple de la betterave à sucre (sélectionner Photosynthèse)
- Qu'est-ce que la biomasse ?
- La Photosynthèse sur le site de l'Université Pierre et Marie Curie - Très complet sur la photosynthèse.
- La Photosynthèse dans le cycle du Carbone et de l'Oxygène


Wikimedia Foundation. 2010.