- Comportement de reproduction
-
 Pour les articles homonymes, voir sexualité (homonymie).
Pour les articles homonymes, voir sexualité (homonymie). Copulation entre une femelle et un mâle chez les félidés
Copulation entre une femelle et un mâle chez les félidés
 Copulation entre une femelle et un mâle chez les canidés
Copulation entre une femelle et un mâle chez les canidés Copulation entre une femelle et un mâle chez les ursidés
Copulation entre une femelle et un mâle chez les ursidés Copulation entre une femelle et un mâle chez les félidés
Copulation entre une femelle et un mâle chez les félidésChez les animaux, le comportement de reproduction[1] correspond à l'ensemble des séquences motrices qui sont exécutées pour mettre en contact les gamètes du mâle avec celles de la femelle, ce qui permet la fécondation et la reproduction de l'espèce.
Le comportement de reproduction est contrôlé par le système nerveux. Plus le système nerveux est simple, plus le comportement est simple et stéréotypé (Insectes, Annélides …). Au contraire, plus le système nerveux est complexe, plus le comportement est élaboré (Mammifères, et surtout les Hominidés).
Chez la plupart des animaux, le comportement sexuel est un comportement de reproduction : la fécondation est le but de ce comportement. Chez les mammifères ayant un cerveau très développé (Homo sapiens, Chimpanzés, Bonobos, Orang-outan, Dauphin), le but et le contrôle cérébral du comportement sont modifiés : le comportement de reproduction devient un comportement érotique[1].
Description du comportement de reproduction
Chez les mammifères inférieurs (rongeurs, canidés, ovidés …), le comportement de reproduction, et en particulier la copulation, est relativement stéréotypé. (voir les photographies ci-contre)
Le comportement de reproduction est généralement divisé en deux phases : 1) la phase motivationnelle, qui correspond au déclenchement de l'excitation sexuelle, puis au rapprochement physique des partenaires, et 2) la phase consommatoire, qui correspond à la copulation.
Phase motivationnelle
La phase motivationnelle permet aux partenaires d'échanger des stimulations adéquates, qui permettent de déclencher l'excitation sexuelle, de reconnaître le partenaire de sexe opposé et d'engager le rapprochement physique des corps.
En général, par exemple chez le rat, on observe les séquences suivantes : le rat mâle effectue une exploration olfactive, puis éventuellement gustative de la femelle, plus particulièrement de sa région génitale. Cette exploration est d'autant plus longue que le rat est sexuellement inexpérimenté. Il se frotte parfois contre la femelle, et passe en dessous ou au-dessus d'elle. Au cours de ces frottements, on observe fréquemment un comportement apparemment de marquage de la femelle par l'urine du mâle. La rate œstrale répond aux stimulations du mâle par un comportement de saut et de fuite. Si le mâle n'est pas assez expérimenté, la femelle peut également prendre l'initiative en poussant les flancs de son partenaire. Durant ces activités, les deux partenaires peuvent émettre des ultrasons. Les stimulations du mâle pendant cette période (tentatives de monte, odeurs, vocalisations) induisent l’apparition de la lordose, la posture copulatoire de la femelle[2].
Phase consommatoire
Puis, lors de la deuxième phase du comportement de reproduction, appelée phase consommatoire, le mâle monte par l'arrière sur le dos de la femelle. Durant la monte, le mâle palpe et étreint les flancs de la femelle avec ses pattes antérieures. La femelle est active et manifeste sa réceptivité par l'adoption d'une position de lordose et par l'orientation correcte de sa région génitale, ce qui facilite l'intromission du pénis dans le vagin. Le mâle effectue des poussées pelviennes qui permettent l'intromission du pénis et l'éjaculation. Chez le mâle, l'éjaculation s'accompagne de contractions spasmodiques des muscles squelettiques.
Après la copulation, le mâle procède généralement à une toilette de sa région génitale puis entre dans une période d'inactivité. L'éjaculation est suivie chez le mâle d'une période réfractaire caractérisée par une très faible réceptivité à tout type de stimuli et par un état de veille calme proche du sommeil[2].
Facteurs à l'origine du comportement de reproduction
Une étape cruciale dans l'étude d'un comportement est d'identifier, de décrire et d'évaluer l'importance des différents facteurs qui sont : 1) à l'origine du développement de ce comportement, et 2) qui participent à son fonctionnement à l'état adulte.
Pour simplifier, on peut distinguer plusieurs types de facteurs :
- les facteurs primordiaux : ceux qui sont les plus importants et qui interviennent avant les autres.
- les facteurs innés : ceux qui existent dès l'origine dans l'organisme.
- les facteurs acquis : ceux qui sont en général appris au cours des interactions avec l'environnement.
Ces différents facteurs déterminent le développement et la dynamique du comportement de reproduction.
Facteurs innés
Chez les mammifères inférieurs, et en particulier chez les rongeurs, les recherches en neurosciences aux alentours des années 2000 ont permis d'identifier de nombreux éléments innés, tant instinctuels que physiologiques ou autonomes, qui peuvent participer directement et indirectement à la réalisation appropriée du coït vaginal fécondant :
- les hormones contrôlent le comportement de reproduction par une inhibition saisonnière (la copulation n'a lieu qu'à la saison propice), une inhibition pubertaire (la copulation n'a lieu qu'à la période de maturité des organes reproducteurs), et une inhibition œstrale (la copulation n'a lieu qu'à la période périovulatoire)[3] ;
- les phéromones permettent le déclenchement de l'excitation sexuelle et la reconnaissance du partenaire de sexe opposé ;
- les réflexes sexuels (lordose, érection, poussées pelviennes, éjaculation, ovulation réflexe lors du coït ...) permettent l'exécution motrice du coït vaginal et optimisent la fécondation ;
- les récompenses/renforcements permettent le développement de la motivation sexuelle et divers apprentissages qui optimisent la réalisation du comportement de reproduction.
Les hormones sexuelles
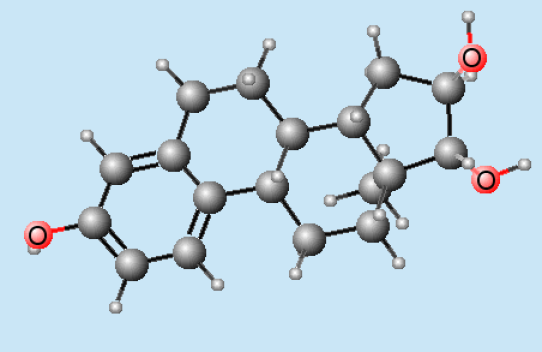
 Hormones : molécule d'estriol
Hormones : molécule d'estriol Hormones : molécule de testostérone
Hormones : molécule de testostéroneLes hormones, et en particulier les hormones sexuelles, activent, coordonnent et contrôlent la plupart des autres facteurs du comportement de reproduction[4].
Les hormones sexuelles ont deux rôles principaux : un rôle organisationnel et un rôle activationnel.
Rôle organisationnel. Au cours du développement, les hormones sexuelles provoquent la différentiation sexuelle de l'organisme en mâle et en femelle. Les différenciations sexuelles du cerveau qui influencent le comportement de reproduction sont situées principalement au niveau de l'amygdale, de la région préoptique, de l'hypothalamus, et de la strie terminale.
Rôle activationnel. À la période de maturité reproductrice, les hormones ont également un rôle d'activation et de contrôle du comportement.
- À la puberté, elles activent le comportement de reproduction (facilitation de la motivation sexuelle, facilitation des réflexes sexuels, et synthèse et émission des phéromones sexuelles)[5].
- Puis, elles exercent un contrôle temporel :
- Enfin, elles permettent la coordination du comportement de reproduction avec la physiologie de l'appareil reproducteur.
En conclusion, les hormones sont un facteur majeur du comportement de reproduction.
Les phéromones sexuelles
 Phéromones : molécule d'ectocarpene
Phéromones : molécule d'ectocarpeneChez les mammifères, on trouve à l'intérieur du nez des structures sensorielles innées, spécialisées dans la détection des phéromones sexuelles :
- l'organe voméronasal ;
- et certaines régions de l'épithélium olfactif.
Les rôles principaux des phéromones sexuelles sont :
- De déclencher l'excitation sexuelle (via le système olfactif principal et voméronasal[7],[8]
- et de permettre la reconnaissance du partenaire de sexe opposé (via le système voméronasal[9],[10],[11]).
Les phéromones permettent la réalisation de la partie initiale du comportement de reproduction, c'est-à-dire l'excitation et le rapprochement physique des partenaires, qui sont les préalables qui vont provoquer les séquences motrices de la copulation.
Remarque importante : les phéromones sont le principal mode de communication des organismes vivants[12]. Ce qui explique leur importance dans le comportement de reproduction.
Les réflexes sexuels
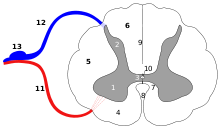 Arc réflexe inné : (12) entrée sensorielle, (2-1) interconnexions dans la moelle épinière, (11) sortie motrice.
Arc réflexe inné : (12) entrée sensorielle, (2-1) interconnexions dans la moelle épinière, (11) sortie motrice.Chez les mammifères, il existe 3 types de réflexes sexuels innés : moteurs, autonomes et neuroendocriniens.
- les réflexes moteurs :
- les réflexes autonomes :
- la lubrification vaginale chez la femelle.
- l'érection[14] et l'éjaculation[15],[16] chez le mâle.
- et un réflexe neuroendocrinien :
Les réflexes sexuels innés permettent la réalisation des séquences finales du comportement de reproduction, c'est-à-dire la copulation, qui est déclenchée par le contact physique génito-génital des corps.
Les récompenses/renforcements
Le système cérébral des récompenses/renforcements a été découvert par Olds & Milner en 1954. Il est localisé dans l'aire préoptique médiane et surtout dans le noyau accumbens. Les principaux neuromédiateurs de la récompense/renforcement sont la dopamine, les cannabinoïdes endogènes[18] et les opioïdes endogènes.
Chez le rat, un stimulus aversif (odeur répugnante), mais couplé avec l'éjaculation, devient appétitif. Ce qui montre, que déjà chez les mammifères les plus simples, les récompenses/renforcements érotiques peuvent être à l'origine de puissants conditionnements qui modifient le comportement sexuel[19].
Les récompenses/renforcements provoquent la répétition de l'activité qui a activé ces processus. Ces processus, associés à la peau poilue du corps[20], aux zones érogènes, à la copulation[21],[22] ainsi qu'à l'éjaculation et à l'orgasme[23], seraient à l'origine de la répétition de toutes les activités agréables de stimulation du corps et surtout des zones érogènes génitales. Ces processus seraient également, mais indirectement, à l'origine de l'apprentissage de la plupart des séquences non innées du comportement de reproduction[24] (voir section “Facteurs acquis” ci-dessous).
Facteurs acquis
En plus de ces facteurs innés, des recherches ont mis en évidence des éléments cruciaux, absolument nécessaires à la réalisation du comportement de reproduction, mais qui eux ne sont pas innés[25]. Ces éléments, qui sont acquis au cours de la période du développement de l'organisme, sont :
- la reconnaissance des congénères ;
- la motivation sexuelle à rechercher le contact physique génital ;
- la socialisation sexuelle ;
- la capacité au positionnement génito-génital approprié des corps.
On remarque que ces éléments du comportement de reproduction qui nécessitent un apprentissage sont en fait ceux qui seraient les plus difficiles à précabler ou à coder génétiquement. La reconnaissance des congénères, la motivation sexuelle à rechercher le contact physique génital, la socialisation sexuelle et la capacité au positionnement génito-génital approprié des corps, nécessiteraient en effet l'existence innée de représentations élaborées du corps (de l'individu et de celui du partenaire), qui ne pourraient être codées que dans les régions les plus complexes du système nerveux. En fonction des connaissances neurobiologiques actuelles, le codage inné de telles capacités cognitives est difficilement explicable[26].
Par contre, en étudiant l'environnement dans lequel se développent les jeunes mammifères, on remarque que toutes ces capacités non innées peuvent être apprises au cours du développement, lors des nombreuses interactions physiques et sociales avec la mère et les congénères[27],[28]. Les données qui montrent que ces capacités ne sont pas innées, ainsi que les arguments expérimentaux qui donnent des indications sur les conditions probables de leur apprentissage, sont détaillés dans les sections ci-dessous.
Reconnaissance des congénères
Comment un animal fait-il pour savoir qu'il doit, pour se reproduire, s'accoupler avec un partenaire de la même espèce ?
Cette capacité de reconnaissance des congénères n'est pas innée. En effet, des animaux élevés dès la naissance par une mère d'une autre espèce (chevreau par brebis et agneau par chèvre) manifestent une attraction sexuelle pour les animaux de leur espèce adoptive et pas pour ceux de leur espèce génétique[29]. Cette expérience montre qu'il n'existe apparemment pas d'informations innées relatives aux caractéristiques spécifiques des congénères chez les capridés, et, vraisemblablement, chez les autres mammifères.
On remarque d'ailleurs qu'il n'existe aucune nécessité à coder génétiquement ces informations, puisque les caractéristiques morphologiques, olfactives, auditives et visuelles des congénères sont forcément toujours apprises : sauf cas exceptionnel, un nouveau-né est toujours en contact et se développe avec des membres de son espèce. L'apprentissage des caractéristiques des congénères est donc prédéterminé par le contexte du développement et il est toujours effectué.
Socialisation sexuelle
Comment un animal fait-il pour connaître les attitudes et les signaux permettant d'engager un comportement sexuel ?
Cette capacité de socialisation sexuelle, c'est-à-dire l'ensemble des attitudes et des réactions appropriées à l'interaction sociale sexuelle avec les congénères, ne serait pas innée. En effet, on observe que des animaux élevés en isolement social dès la naissance sont incapables d'interactions sociales et sexuelles normales[30]. Il est d'ailleurs difficile d'expliquer, en fonction des connaissances neurobiologiques actuelles, de quelle manière toutes les capacités sensorielles, émotionnelles et cognitives nécessaires aux interactions sociales et sexuelles appropriées peuvent être programmées dans le système nerveux.
La socialisation sexuelle est apprise au cours des nombreux et fréquents jeux sociaux qui sont pratiqués quotidiennement par les jeunes mammifères durant toutes les années de leur développement[31]. Ces activités ludiques sont fréquemment répétées car elles procurent des récompenses, en particulier durant la période du développement[32].
Motivation sexuelle
D'où provient la motivation d'un animal à copuler ?
Cette motivation sexuelle à rechercher le contact physique génital ne serait pas innée. En effet, quels sont les processus neurobiologiques qui provoquent l'attirance réciproque d'un mâle et d'une femelle puis qui provoquent la copulation ?
En fonction des connaissances disponibles en 2010, on peut expliquer la reconnaissance du partenaire et l'état d'excitation sexuelle par les phéromones et les hormones. En schématisant, les phéromones sexuelles permettent la reconnaissance du partenaire[9],[10] et l'induction d'un état d'excitation sexuelle[33],[34],[7], par des connexions entre les récepteurs olfactifs et les neurones à gonadolibérine de l'hypothalamus[8],[35], qui contrôlent les hormones sexuelles.
Mais, une fois que le mâle et la femelle sont dans un état d'excitation sexuelle – état neural favorable au déclenchement de différentes actions physiologiques ou motrices – quels sont les processus qui vont les amener à la copulation ?
On peut envisager qu'il existerait des connaissances, des représentations ou des séquences motrices innées, spécifiques du coït vaginal. Mais aucune donnée actuelle ne permet de prouver ni même d'expliquer comment des phénomènes cognitifs aussi complexes que des connaissances ou des représentations spécifiques à la copulation peuvent être codés dans les cortex associatifs[25]. Quant aux séquences motrices innées, les seules actuellement connues sont les réflexes sexuels (lordose, intromission …) qui permettent la réalisation de la partie finale de la copulation.
En récapitulant, les processus innés connus expliquent l'excitation sexuelle des animaux, la reconnaissance du partenaire de sexe opposé, et, seulement si les animaux parviennent en contact génito-génital, les réflexes innés permettront le coït vaginal. Mais il manque une motivation innée, spécifique à entrer en contact physique approprié au niveau des zones génitales.
Néanmoins, plusieurs expériences suggèrent que cette motivation particulière est acquise durant le développement, grâce en particulier aux récompenses génitales. En effet, la stimulation corporelle, en particulier au niveau des zones génitales, procure des récompenses. Durant les mois ou les années que dure le développement, les milliers de contacts corporels postnatals avec la mère et avec les autres nouveau-nés, et en particulier le léchage génital[36],[37],[27], ainsi que les jeux sexuels avec les autres congénères, et surtout l'exploration régulière phéromone-dépendante de la région génitale des congénères[30], induisent le développement graduel d'une motivation sexuelle à rechercher des stimulations génitales.
Cette motivation sexuelle acquise est ensuite fortement potentialisée à la puberté sous l'effet des hormones sexuelles, vraisemblablement par une action modulatrice de la testostérone, dans la région préoptique, sur les enzymes contrôlant la synthèse des neuromédiateurs impliqués dans les processus sexuels[38].
Séquence motrice de la monte
Comment un mâle fait-il pour savoir qu'il faut monter sur le dos de la femelle, par l'arrière, afin de copuler ?
La capacité au positionnement génito-génital approprié des corps (la séquence de la monte chez les mammifères inférieurs) n'est pas innée. En effet, on observe que lorsque les mammifères non humains, et tout particulièrement les primates, sont mis dans des conditions où ils ne peuvent apprendre aucun élément du comportement de reproduction, de manière systématique le mâle est incapable de coïter[28],[39],[40],[27]. Cette incapacité du mâle à pratiquer le coït en dehors de toute expérience préalable a été vérifiée dans plusieurs espèces (cochon d'Inde, rat, chat, chien, macaque rhésus, chimpanzé), et elle est systématique chez tous les primates.
En variant les conditions expérimentales, il est apparu que c'est vraisemblablement la privation du contact physique, et non de la vue, des sons ou des odeurs des congénères, qui est le facteur critique à l'origine du déficit coïtal[28],[27]. On peut ainsi observer des mâles sexuellement naïfs manifester de nombreux comportements spécifiques de la reproduction (excitation, érection, contacts avec le partenaire …) mais ils ne parviennent pas à copuler. Un des principaux problèmes, bien qu'il existe également des problèmes connexes de socialisation et de peur des autres congénères[41], pourrait être lié à un déficit de construction du “schéma corporel” et se caractérise par une incapacité à positionner correctement le corps afin de réussir l'intromission[42]. L'absence d'innéité du positionnement génito-génital n'est guère surprenante. Comment cette capacité cognitive complexe, qui nécessite l'existence innée de représentations et de positions spécifiques du corps dans l'espace, serait-elle pré-programmée dans le système nerveux ? En fonction des connaissances disponibles en 2009, c'est apparemment inexplicable.
En fait, cette capacité est apprise durant les mois ou les années que dure le développement, au cours des milliers d'interactions corporelles et des jeux sexuels avec les congénères. Comme déjà indiqué, ces activités sont fréquemment répétées car elles procurent des récompenses[32]. Les premiers contacts corporels postnatals et plus particulièrement tous les types de contacts et d'explorations corporelles effectués au cours des jeux sociaux sont à l'origine de la construction d'un “schéma corporel” de l'individu et des congénères. L'apprentissage d'un schéma corporel permet à l'animal de pouvoir réaliser des ajustements posturaux adaptés – dont le positionnement génito-génital – lors des interactions corporelles spécifiques avec ses congénères (toilettage, agression, copulation …).
De plus, au cours de toutes ces interactions corporelles, toutes celles qui provoquent des contacts physiques avec les régions génitales ou qui déclenchent des réflexes sexuels peuvent ainsi amorcer des séquences sexuelles partielles (jeux sexuels), qui sont progressivement intégrés par apprentissage dans des schèmes moteurs plus globaux[42], initiant ainsi progressivement l'apprentissage d'un comportement de reproduction plus complet.
Au niveau neurobiologique, tous ces apprentissages réalisés grâce à ces activités et ces stimulations corporelles se traduisent concrètement par le développement des structures neurales impliquées dans le contrôle du comportement de reproduction (amygdale médiale, noyau préoptique médian, noyaux moteurs médullaires)[43],[37],[44].
On remarque ainsi que l'apprentissage des schèmes moteurs plus globaux correspond au développement des structures qui sont impliquées dans le contrôle du comportement de reproduction. Le contrôle que ces structures exercent sur le comportement de reproduction n'est donc pas un contrôle inné, dépendant d'une organisation génétiquement ou hormonalement “programmée” de ces structures, mais un contrôle en grande partie acquis. On observe ainsi que l'interaction entre les facteurs environnementaux (jeux sociaux et sexuels) et les facteurs innés (réflexes sexuels, phéromones, récompense) permet le développement structurel et fonctionnel du circuit neural spécifique au comportement de reproduction[45].
Modélisation du comportement de reproduction
En fonction de toutes les données présentées dans les sections précédentes, quel serait, en simplifiant, le développement et de la dynamique du comportement de reproduction des mammifères ?
Développement du comportement de reproduction
Dans la période prénatale, sous l'effet organisateur des hormones (qui sont le facteur majeur et primordial), l'organisme se différencie en mâle ou en femelle[4]. Les structures innées (réflexes sexuels, organe voméronasal, connexions olfactives …) se développent, ainsi que le système de récompense et les capacités d'apprentissage.
Puis, dès la naissance, le petit mammifère apprend au contact de sa mère et des congénères les caractéristiques sensorielles des animaux de son espèce. Au cours des nombreuses interactions physiques et sociales quotidiennes, il acquiert par les jeux physiques un schéma corporel[42], par les jeux sociaux il apprend à interagir socialement de manière appropriée[31], et, surtout pour le mâle, ses circuits neuraux, ses réflexes sexuels et sa motivation sexuelle se développent progressivement au cours des jeux sexuels[43],[37],[44].
De cette manière, tous les éléments innés et acquis permettant le comportement de reproduction deviennent fonctionnels avant la puberté.
Dynamique du comportement de reproduction à la maturité
À la puberté, les hormones sont toujours le facteur majeur. Par rapport au comportement, elles ont principalement un rôle de coordination et de modulation, en rendant actifs de manière simultanée les différents processus neurobiologiques impliqués dans le comportement de reproduction.
La mélatonine, par son action sur les neurones à gonadolibérine, active le comportement sexuel durant la saison favorable à la reproduction[6]. Les hormones sexuelles initient le comportement de reproduction à la puberté[5], lèvent les inhibitions toniques sur les réflexes sexuels, abaissent les seuils de détection des organes sensoriels[46], déclenchent la synthèse et la libération des phéromones sexuelles et potentialisent la motivation sexuelle. L'organisme est ainsi prêt pour effectuer les séquences comportementales aboutissant à la copulation.
Puis, lors de la phase motivationnelle, lorsque les animaux sexuellement naïfs sont en présence, les phéromones sont les principaux signaux primaires qui provoquent le déclenchement de la motivation sexuelle[33] et permettent la discrimination du partenaire sexuel de sexe opposé[9],[10]. L'état d'excitation sexuelle et la vue du partenaire provoquent le rappel contextuel[47] des souvenirs des jeux sexuels antérieurs, ce qui induit la motivation à entrer en contact physique génital, pour obtenir de nouvelles récompenses sexuelles.
Enfin, lors de la phase consommatoire, lorsque les animaux sont en contact physique, les stimuli corporels provoqués par chaque action constituent les stimuli déclencheurs de l'action réflexe suivante[48]. Chaque action déclenche l'action suivante : la monte déclenche la lordose[13], qui déclenche les poussées pelviennes[2] et l'intromission, qui déclenchent alors l'éjaculation[15],[16] et la libération de l'ovule[17].
Après que l'animal naïf a réalisé plusieurs copulations, différents signaux (visuels, auditifs, gustatifs, etc.) deviennent par conditionnement des signaux sexuels qui peuvent déclencher et influencer les copulations ultérieures[49]. Le comportement de reproduction peut alors être effectué malgré l'absence de certains signaux innés (comme les phéromones sexuelles) qui sont cruciaux pour l'animal sexuellement naïf[50],[51]. Puis, au fur et à mesure de l'accroissement de l'expérience sexuelle et des apprentissages concomitants, les différentes séquences motrices sont exécutées plus efficacement et le comportement de reproduction devient mature et optimisé.
Analyses phylogénétiques et fonctionnelles
À partir de toutes les données présentées dans les sections précédentes, en analysant l'organisation et la dynamique fonctionnelle des processus innées et des apprentissages qui participent à la réalisation du comportement de reproduction, il apparaît deux caractéristiques remarquables : l'existence d'un véritable comportement de reproduction, et l'existence d'un instinct partiel. De plus, les phéromones apparaissent comme étant le facteur de l'orientation sexuelle.
Orientation sexuelle & Préférences sexuelles
Orientation sexuelle. L'analyse des facteurs innés à l'origine du comportement de reproduction met en évidence que le seul facteur inné qui permet d'identifier le partenaire de sexe opposé est celui des phéromones sexuelles[9],[10]. Cette importance des phéromones dans la communication sexuelle des mammifères est en cohérence avec l'importance fondamentale des phéromones dans la communication de quasiment tous les organismes vivants (procaryotes, champignons, algues, végétaux, insectes, poissons, mammifères …)[12].
Préférences sexuelles. Il ne semble pas exister de préférences sexuelles chez les rongeurs. Des préférences, en général pour un ou plusieurs partenaires sexuels avec lesquels se forment des relations privilégiées, n'apparaissent clairement qu'à partir des primates.
Article détaillé : Préférence sexuelle.Véritable comportement de reproduction
L'analyse des facteurs innés permettant le comportement de reproduction met en évidence une caractéristique remarquable :
- on remarque qu'il existe un contrôle temporel optimisé, réalisé par les hormones. Ce contrôle est exercé à trois niveaux :
- un contrôle pubertaire : les activités sexuelles n'ont lieu qu'à la période de maturité de l'appareil reproducteur ;
- un contrôle saisonnier : les activités n'ont lieu qu'à la saison propice ;
- et un contrôle œstral : les activités n'ont lieu qu'à la période de maturité des gamètes ;
- il existe une identification du partenaire de sexe opposé, réalisé grâce aux phéromones ;
- il existe des activités réflexes innés spécifiques du coït vaginal : lordose, érection, éjaculation, etc.
- et il existe une ovulation réflexe lors du coït, qui permet une optimisation de la fécondation.
En synthèse, ces facteurs innés tendent à ce que la copulation soit hétérosexuelle, ait lieu à la saison propice et à la période de maturité de l'ovule. Ce qui signifie que dans le plan d'organisation général des mammifères, il existe un véritable comportement de reproduction[45].
Concept d' “instinct partiel”
Les données présentées ci-dessus et décrivant les facteurs innés et acquis à l'origine du comportement de reproduction montrent qu'il n'existe pas d'instinct de la reproduction (ou d'instinct sexuel), c'est-à-dire une programmation centrale innée des différentes séquences motrices nécessaires à la réalisation de ce comportement.
On observe plutôt que le comportement de reproduction émerge principalement de la coordination fonctionnelle de différents réflexes et processus sexuels innés, d'interactions avec les congénères et de différents apprentissages qui sont toujours réalisés au cours du développement dans l'environnement écologique normal.
Ces caractéristiques particulières amènent à proposer le concept d'un “instinct partiel”, c'est-à-dire un ensemble incomplet d'éléments innés, mais, parce qu'ils sont associés à des circonstances environnementales spécifiques qui existent quasiment toujours au cours du développement (par exemple les interactions mère-enfant, les stimuli sensoriels et les contacts corporels avec les congénères, etc.), les éléments manquants sont indirectement toujours acquis et le comportement “partiellement instinctuel” s'exécute correctement à sa période de maturité, sans nécessité d'apprentissages spécifiques[25].
À noter que pour un observateur extérieur, le comportement de reproduction peut apparaître comme instinctuel, dans la mesure où les apprentissages, tels ceux induits par le léchage de la région génitale, ne sont pas apparents, ne sont pas directement liés à la reproduction ou ne sont pas considérés comme “sexuels”.
Variations du comportement de reproduction
Le modèle du comportement de reproduction présenté ci-dessus correspond à un modèle général, prototypique, basé sur les connaissances en neurosciences disponibles en 2009. Le cadre général de ce modèle est valable pour tous les mammifères inférieurs.
Mais pour compléter ce modèle, certaines précisions doivent être apportées afin d'expliciter certains cas particuliers : les variations du comportement suivant les espèces de mammifères, l'existence d'activités non reproductrices, et, plus paradoxal encore, l'existence d'activités sexuelles entre espèces différentes.
Variations en fonction des espèces
Suivant les espèces de mammifères inférieurs (rats, chiens, chèvres, moutons …) certains facteurs innés (réflexes sexuels, phéromones, hormones, récompense …) n'existent pas ou plus, ou sont modifiés. Par exemple, le contrôle saisonnier ou le réflexe neuroendocrinien d'ovulation n'existent pas dans plusieurs espèces (l'ovulation réflexe existe chez le furet, la lapine, ou la chatte, mais pas chez la brebis ou la ratte).
Néanmoins, le modèle général reste toujours valable. Seule la dynamique du comportement est modifié. Par exemple, s'il n'existe pas de contrôle saisonnier, alors les copulations existent toute l'année, et des naissances peuvent avoir lieu à des saisons moins favorables. Le comportement de reproduction reste toujours fonctionnel, il est seulement moins optimisé.
Activités sexuelles non reproductrices
Le contrôle exercé par les hormones et les phéromones n'est pas toujours ni parfait, ni total. Et plus on monte dans l'échelle phylogénétique, plus le cerveau est développé, plus le contrôle hormonal s'affaiblit[4]. En particulier pour le mâle, des réflexes sexuels peuvent être exécutés avant la puberté, bien que le système reproducteur de l'animal ne soit pas mature. Il n'est donc pas “biologiquement anormal” d'observer parfois chez les mammifères inférieurs des activités sexuelles sans rapport avec la reproduction.
Activités sexuelles inter-espèces
Le comportement sexuel a priori le plus aberrant, le plus “contre-nature”, est la copulation entre deux animaux appartenant à des espèces différentes.
Néanmoins, on remarque que ce comportement existe et que ces activités inter-espèces sont d'autant plus fréquentes que la morphologie des organismes est similaire (âne/cheval, tigre/lion, etc.)[52]
Est-ce une anomalie, une pathologie, un dysfonctionnement de l'instinct sexuel ?
Comme indiqué dans les paragraphes précédents, chez les mammifères inférieurs (comme les rongeurs ou les capridés), plusieurs caractéristiques du comportement de reproduction ne sont pas innées. Par exemple, la reconnaissance du congénère est apprise durant les premières années de la vie : comme déjà indiqué, des chevreaux adoptés par des brebis et des agneaux adoptés par des chèvres copulent à l'âge adulte avec des partenaires de l'espèce adoptive et pas avec des membres de leur espèce[29]. On remarque ici que les modifications du comportement de reproduction trouvent leur origine dans des apprentissages précoces[53].
De façon plus générale, toutes les capacités apprises qui sont nécessaires à la réalisation du comportement de reproduction (reconnaissance du congénère, socialisation sexuelle, motivation sexuelle, et positionnement génito-génital du corps), sont influencées par les conditions et le contexte des apprentissages. Et ces apprentissages dépendent des caractéristiques de l'environnement et des interactions entre les animaux.
En reprenant l'exemple d'adoption croisée des chevreaux et des agneaux[29], on remarque que ce qui apparaît a priori comme le plus aberrant et contre-nature, est en fait “biologiquement normal” : les nouveau-nés interagissent avec leurs mères adoptives, et durant toutes les années de leur développement, au cours des milliers d'interactions physiques et sociales, les chevreaux apprennent les différentes caractéristiques sensorielles, comportementales et sociales des brebis, et les agneaux apprennent celles des chèvres. Les chevreaux et les agneaux apprennent les caractéristiques de leurs mères adoptives, apprennent les codes de socialisation de leurs “congénères” adoptifs, ont des jeux sociaux et sexuels avec leurs “congénères” adoptifs, et ainsi il est “biologiquement normal” qu'à l'âge adulte ils copulent avec un partenaire de leur espèce adoptive, et non de leur espèce génétique qui leur est complètement étrangère.
De plus, on observe expérimentalement que lorsqu'un animal sexuellement expérimenté est excité, la force de l'excitation sexuelle est telle que la vue ou le contact avec tout animal ou objet de forme similaire suffit à déclencher la copulation[54]. L'élevage en commun n'est donc parfois même pas nécessaire.
Pour toutes ces raisons, il est “biologiquement normal” que des animaux qui vivent dans certains types d'environnements, par exemple élevés ensemble dans une ferme ou un zoo, puissent avoir des interactions sexuelles (pour exemple, voir les vidéos : lapin avec chat, chien avec poule, chien avec chat, chien avec lion, chien avec chat, tigre avec chien).
Toujours pour ces raisons, il n'est pas “biologiquement anormal” que des animaux domestiques (surtout les chiens) ou élevés dans des zoos (et tout particulièrement les primates) tentent parfois d'avoir des interactions sexuelles avec des humains[52] (et vice-versa d'ailleurs[55]).
En conclusion, comme il n'existe qu'un instinct partiel du comportement de reproduction, tous les types d'activités sexuelles qui n'ont aucun rapport avec la reproduction s'expliquent par l'existence des apprentissages cruciaux (reconnaissance du congénère, positionnement génito-génital du corps, socialisation et motivation sexuelle). Ces apprentissages cruciaux sont influencés par les caractéristiques de l'environnement, et dans certaines circonstances, des apprentissages précoces peuvent induire des modifications comportementales importantes.
Évolution du comportement de reproduction : le comportement érotique
Article principal : Comportement érotique.On remarque qu'au cours de l'évolution, l'importance et l'influence des hormones[56],[57] et des phéromones[58],[59] sur le comportement de reproduction a diminué. Au contraire, l'importance de la récompense/renforcement est devenu majeur[24].
Chez tous les mammifères qui ont un cerveau très développé (Homme, Chimpanzé, Orang outan et Dauphin), le but du comportement sexuel n'est plus le coït vaginal mais la recherche du plaisir érotique, procuré par la stimulation du corps et des zones érogènes.
Le comportement de reproduction devient graduellement un comportement érotique[45].
Les références et les explications détaillées sont présentées dans l'article Comportement érotique.Notes et références
- Les références et les explications détaillées sont données dans cet article et dans l'article Comportement érotique. Les distinctions entre “comportement sexuel”, “comportement de reproduction” et “comportement érotique” ont été proposées par les auteurs Martin H. Johnson et Barry J. Everitt dans leur ouvrage Reproduction, 5e edition, publié chez De Boeck Université en 2001, afin de tenir compte des différences neurobiologiques, psychologiques et comportementales du comportement sexuel entre les espèces. L'ouvrage qui présente le plus de vérifications expérimentales de cette distinction est Functional and dysfunctional sexual behavior du neurobiologiste Anders Agmo.
- MEISEL Robert L. , SACHS Benjamin D. : The physiology of male sexual behavior. in KNOBIL Ernest , NEILL Jimmy D. The physiology of reproduction, Raven Press, 2nd edition, 1994
- NB: l'inhibition saisonnière et l'inhibition œstrale n'existent pas dans certaines espèces. Mais quelles que soient les variations entre les espèces, les hormones restent le facteur primordial du comportement de reproduction.
- THIBAULT Charles ; LEVASSEUR Marie-claire : La reproduction chez les mammifères et l'homme, INRA Ellipse 2001
- SISK C. L. , FOSTER D. L. The neural basis of puberty and adolescence, Nature Neuroscience, 7(10):1040-1047, 2004
- ALEANDRI V. , SPINA V. , MORINI A. The pineal gland and reproduction, Hum. Reprod. Update., 2(3):225-235, 1996
- MONCHO-BOGANI J. , LANUZA E. , HERNANDEZ A. , NOVEJARQUE A. , MARTINEZ-GARCIA F. Attractive properties of sexual pheromones in mice: innate or learned? Physiology & Behavior, 77(1):167-176, 2002
- YOON H. , ENQUIST L. W. , DULAC C. Olfactory inputs to hypothalamic neurons controlling reproduction and fertility, Cell, 123(4):669-682, 2005
- (en) STOWERS L. , HOLY T. E. , MEISTER M. , DULAC C. , KOENTGES G. Loss of sex discrimination and male-male aggression in mice deficient for TRP2, Science, 295(5559):1493-1500, 2002
- DULAC C. , TORELLO A. T. Molecular detection of pheromone signals in mammals: from genes to behaviour, Nat. Rev. Neurosci., 4(7):551-562, 2003
- (fr) CIOFI Philippe, La bisexualité est-elle innée ? Cerveau & Psycho, 25:50-52, 2008
- DESCOINS C., Phéromones, Encyclopédia Universalis, v11.0, 2006
- PFAFF Donald W. , SCHWARTZ-GIBLIN Susan , MACCARTHY Margareth M. , KOW Lee-Ming : Cellular and molecular mechanisms of female reproductive behaviors, in KNOBIL Ernest , NEILL Jimmy D. : The physiology of reproduction, Raven Press, 2nd edition, 1994
- GIULIANO F. , RAMPIN O. Neural control of erection, Physiology & Behavior, 83(2):189-201, 2004
- ALLARD J. , TRUITT W. A. , MCKENNA K. E. , COOLEN L. M. Spinal cord control of ejaculation, World J. Urol., 23(2):119-126, 2005
- COOLEN L. M. Neural control of ejaculation, J. Comp Neurol., 493(1):39-45, 2005
- SPIES H. G. , PAU K. Y. , YANG S. P. Coital and estrogen signals: a contrast in the preovulatory neuroendocrine networks of rabbits and rhesus monkeys, Biol. Reprod., 56(2):310-319, 1997
- MAHLER S. V. , SMITH K. S. , BERRIDGE K. C. Endocannabinoid Hedonic Hotspot for Sensory Pleasure: Anandamide in Nucleus Accumbens Shell Enhances 'Liking' of a Sweet Reward, Neuropsychopharmacology, 2007
- PFAUS J. G. , KIPPIN T. E. , CENTENO S. Conditioning and sexual behavior: a review, Hormones and Behavior, 40(2):291-321, 2001
- (en) OLAUSSON H. , LAMARRE Y. , BACKLUND H. , MORIN C. , WALLIN B. G. , STARCK G. , EKHOLM S. , STRIGO I. , WORSLEY K. , VALLBO A. B. , BUSHNELL M. C. Unmyelinated tactile afferents signal touch and project to insular cortex, Nature Neuroscience, 5(9):900-904, 2002
- CAGGIULA A. R. , HOEBEL B. G. “Copulation-reward site” in the posterior hypothalamus, Science, 153(741):1284-1285, 1966
- CAGGIULA A. R. Analysis of the copulation-reward properties of posterior hypothalamic stimulation in male rats, J. Comp Physiol Psychol., 70(3):399-412, 1970
- HOLSTEGE G. , GEORGIADIS J. R. , PAANS A. M. , MEINERS L. C. , VAN DER GRAAF F. H. , REINDERS A. A. Brain activation during human male ejaculation, The Journal of Neuroscience, 23(27):9185-9193, 2003
- (en) AGMO Anders Functional and dysfunctional sexual behavior
- WUNSCH Serge , BRENOT Philippe. Does a sexual instinct exist ? Sexologies, 13(48):30-36, 2004
- Voir les explications détaillées dans l'article Instinct
- WARD Ingeborg L. Sexual behavior : the product of perinatal hormonal and prepubertal social factors, in GERAL Arnold A. , MOLTZ Howard , WARD Ingeborg L. (Ed), Sexual differentiation, vol 11, Handbook of behavioral neurobiology, Plenum Press, NY, 1992
- GRUENDEL A. D. , ARNOLD W. J. Effects of early social deprivation on reproductive behavior of male rats, J. Comp Physiol Psychol., 67(1):123-128, 1969
- KENDRICK K. M. , HINTON M. R. , ATKINS K. , HAUPT M. A. , SKINNER J. D. Mothers determine sexual preferences, Nature, 395(6699):229-230, 1998
- SPEVAK A. M. , QUADAGNO D. M. , KNOEPPEL D. The effects of isolation on sexual and social behavior in the rat, Behav. Biol., 8(1):63-73, 1973
- VANDERSCHUREN L. J. , NIESINK R. J. , VAN REE J. M. The neurobiology of social play behavior in rats, Neuroscience and Biobehavioral Reviews, 21(3):309-326, 1997
- DOUGLAS L. A. , VARLINSKAYA E. I. , SPEAR L. P. Rewarding properties of social interactions in adolescent and adult male and female rats: impact of social versus isolate housing of subjects and partners, Dev. Psychobiol., 45(3):153-162, 2004
- MONCHO-BOGANI J. , MARTINEZ-GARCIA F. , NOVEJARQUE A. , LANUZA E. Attraction to sexual pheromones and associated odorants in female mice involves activation of the reward system and basolateral amygdala, Eur. J. Neurosci., 21(8):2186-2198, 2005
- MONCHO-BOGANI J. , LANUZA E. , LORENTE M. J. , MARTINEZ-GARCIA F. Attraction to male pheromones and sexual behaviour show different regulatory mechanisms in female mice, Physiology & Behavior, 81(3):427-434, 2004
- BOEHM U. , ZOU Z. , BUCK L. B. Feedback loops link odor and pheromone signaling with reproduction, Cell, 123(4):683-695, 2005
- MOORE C. L. The role of maternal stimulation in the development of sexual behavior and its neural basis, Annals of the New York Academy of Sciences, 662:160-177, 1992
- BAUM M. J. , BRESSLER S. C. , DAUM M. C. , VEIGA C. A. , MCNAMEE C. S. Ferret mothers provide more anogenital licking to male offspring: possible contribution to psychosexual differentiation, Physiology & Behavior, 60(2):353-359, 1996
- DU J. , HULL E. M. Effects of testosterone on neuronal nitric oxide synthase and tyrosine hydroxylase, Brain Research, 836(1-2):90-98, 1999
- MISSAKIAN E. A. Reproductive behavior of socially deprived male rhesus monkeys (Macaca mulatta), J. Comp Physiol Psychol., 69(3):403-407, 1969
- TURNER C. H. , DAVENPORT R. K. , ROGERS C. M. The effect of early deprivation on the social behavior of adolescent chimpanzees, The American journal of psychiatry, 125(11):1531-1536, 1969
- GOLDFOOT D. A. Rearing conditions which support or inhibit later sexual potential of laboratory-born rhesus monkeys: hypothesis and diagnostic behaviors, Lab Anim Sci., 27(4):548-556, 1977
- HARD E. , LARSSON K. Climbing behavior patterns in prepubertal rats. Effects of hormones, rearing conditions, and sensory cues, Brain, Behavior and Evolution, 4(2):151-161, 1971
- MOORE C. L. Maternal contributions to the development of masculine sexual behavior in laboratory rats, Dev. Psychobiol., 17(4):347-356, 1984
- COOKE B. M. , CHOWANADISAI W. , BREEDLOVE S. M. Post-weaning social isolation of male rats reduces the volume of the medial amygdala and leads to deficits in adult sexual behavior, Behavioural Brain Research, 117(1-2):107-113, 2000
- (fr) WUNSCH Serge, Thèse de doctorat sur le comportement sexuel [PDF] EPHE-Sorbonne, Paris, 2007.
- GANDELMAN R. : Gonadal hormone and sensory function, Neurocience and biobehavioral reviews, 7:1-18, 1983
- Rappel contextuel : rappel d'un souvenir provoqué par la perception d'un contexte similaire au contexte de mémorisation de ce souvenir. GERRIG Richard , ZIMBARDO Philip. Psychologie, Pearson education, 18e édition, 2008
- BALTHAZART J. , FABRE-NYS C. Le comportement sexuel, in THIBAULT C. , LEVASSEUR M.-C. (Eds). La reproduction chez les mammifères et l'Homme, INRA Ellipse, 2(27):611-637, 2001
- WOODSON James C. Including 'learned sexuality' in the organization of sexual behavior, Neuroscience and Biobehavioral Reviews, 26(1):69-80, 2002
- MEREDITH Michael : Sensory processing in the main and accessory olfactory systems : comparisons and contrasts, Journal of steroid biochemistry and molecular biology, 39/4B:601-614, 1991
- SIGNORET Jean-Pierre , BALTHAZART Jacques : Le comportement sexuel, in THIBAULT Charles ; LEVASSEUR Marie-claire : La reproduction chez les mammifères et l'homme, INRA Ellipse 1991
- (en) FORD Clellan S. , BEACH Frank A. : Patterns of sexual behavior, Methuen & Co, London, 1965. Le livre existe en français, mais il est plus difficile à trouver : (fr) Le comportement sexuel chez l'homme et l'animal, R. Laffont, 1970
- L'importance du vécu et des apprentissages était déjà vérifié expérimentalement dès les années 1950 : « L'expérience et l'apprentissage influencent la tendance des mammifères inférieurs à avoir des réponses sexuelles pour des partenaires d'une autre espèce. » FORD Clellan S. , BEACH Frank A. : Patterns of sexual behavior, Methuen & Co, London, 1965
- « Les mammifères mâles qui sont habitués à recevoir des femelles réceptives dans une même cage, souvent deviennent si excité par les caractéristiques de cet environnement qu'ils vont tenter de s'accoupler avec un animal d'une autre espèce. Les lapins mâles qui copulent régulièrement dans leur cage vont essayer de copuler avec des rats, des petits chats, et même des objets inanimés s'ils sont présentés de la même manière que la femelle réceptive. Les rats mâles qui ont copulé avec une femelle dans une cage d'observation essayent de copuler avec quasiment tout animal d'une taille appropriée qui est rencontré dans cette cage expérimentale. » FORD Clellan S. , BEACH Frank A. : Patterns of sexual behavior, Methuen & Co, London, 1965
- KINSEY Alfred C., POMEROY Wardell B., MARTIN Clyde E. Le comportement sexuel de l'homme. Rayonnement de la pensée, 1948
- SIGNORET Jean-Pierre, Sexuel (Comportement), Encyclopædia Universalis. v. 11.0, 2006.
- BUVAT J. : Hormones et comportement sexuel de l'Homme : données physiologiques et physiopathologiques, Contracept. Fertil. Sex., 24/10:767-778, 1996
- (en) ZHANG J. , WEBB D. M. Evolutionary deterioration of the vomeronasal pheromone transduction pathway in catarrhine primates, Proceedings of the National Academy of Sciences of the United States of America, 100(14):8337-8341, 2003
- FOIDART A. , LEGROS J.J. , BALTHAZART J. : Les phéromones humaines : vestige animal ou réalité non reconnue, Revue médicale de Liège, 49/12:662-680, 1994
Bibliographie
- (fr) C. Thibault, M.-C. Levasseur (Eds). La reproduction chez les mammifères et l'Homme, INRA Ellipse, 2001
- (en) Donald Pfaff D., Arthur ARNOLD. Hormones, Brain and Behavior, Academic Press 2002
- (fr) Serge Wunsch, Thèse de doctorat sur le comportement sexuel [PDF] EPHE-Sorbonne, Paris, 2007.
- (en) Anders Agmo Functional and dysfunctional sexual behavior Elsevier 2007
- (en) M. Breedlove, M. Rosenzweig, N Watson. Biological Psychology, an introduction to behavioral, cognitive, and clinical neuroscience, 5e edition, Sinauer Associates, 2007
Annexes
Articles connexes
- Sexualité, l'essentiel en un article
- Comportement érotique, les bases neurobiologiques de la sexualité humaine
- Comportement sexuel humain, les comportements sexuels de l'être humain
- Sexualité humaine, les phénomènes culturels de la sexualité humaine
- Sexualité infantile, le développement sexuel chez l'être humain
- Sexualité féminine, les caractéristiques de la sexualité féminine
- Reproduction humaine, la physiologie de la reproduction humaine
- Comportement sexuel, les comportements sexuels de l'animal
- Sexualité animale, l'émergence des phénomènes culturels chez l'animal
- Sexualité (reproduction), la reproduction sexuée chez les organismes vivants
Liens externes
L'Université de Berlin, la plus importante base de données sur la sexualité humaine (Documents en français / Accueil principal)
 Portail de la sexualité et de la sexologie
Portail de la sexualité et de la sexologie Portail des neurosciences
Portail des neurosciences






Wikimedia Foundation. 2010.