- Dopamine
-
Dopamine 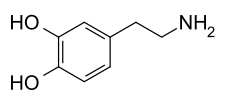

Molécule de dopamineGénéral Nom IUPAC 4-(2-aminoéthyl)benzène-1,2-diol Synonymes oxytyramine
hydroxytyramine
intopine
3,4-dihydroxyphényléthylamineNo CAS No EINECS Code ATC C01 DrugBank PubChem ChEBI SMILES InChI Apparence poudre blanche à odeur distinctive Propriétés chimiques Formule brute C8H11NO2 [Isomères] Masse molaire[1] 153,1784 ± 0,008 g·mol-1
C 62,73 %, H 7,24 %, N 9,14 %, O 20,89 %,Précautions Directive 67/548/EEC 
XiPhrases R : 36/37/38, Phrases S : 26, 36/37, Unités du SI & CNTP, sauf indication contraire. La dopamine est un neurotransmetteur appartenant aux catécholamines et donc issue de l'acide aminé tyrosine. Dans le système nerveux central, elle agit comme neurotransmetteur en activant les récepteurs dopaminergiques postsynaptiques. La dopamine est principalement produite dans la substance noire et dans l'aire tegmentale ventrale[2], situées dans le mésencéphale (partie supérieure du tronc cérébral). Bien que la dopamine, avec la noradrénaline et la sérotonine, soient très minoritaires dans le cerveau, puisqu'ensemble, elles concernent moins de 1 % des neurones[3], elles jouent un rôle modulateur final des sorties motrices et psychiques essentiel.
La dopamine est également une neurohormone produite par l'hypothalamus. Sa principale fonction en tant qu'hormone est d'inhiber la libération de prolactine par le lobe antérieur de l'hypophyse.
La dopamine est le précurseur de l'adrénaline et de la noradrénaline.
Sommaire
Historique
Sa fonction de neurotransmetteur a été découverte en 1958 par Arvid Carlsson et Nils-Åke Hillarp au Laboratoire de pharmacologie du Conseil national cardiologique de Suède. Elle a été nommée dopamine car il s'agit d'une monoamine dont le précurseur direct est le 3,4-dihydroxyphénylalanine (L-DOPA). Arvid Carlsson a reçu en 2000 le prix Nobel de physiologie ou médecine pour avoir démontré que la dopamine n'est pas seulement un précurseur de la norépinéphrine (noradrénaline) et de l'épinéphrine (adrénaline), mais également un neurotransmetteur.
La dopamine avait été synthétisée pour la première fois en 1910 par George Barger et James Ewens au Wellcome Laboratories à Londres, en Angleterre.
Métabolisme de la dopamine
 Métabolisme de la dopamine
Métabolisme de la dopamine
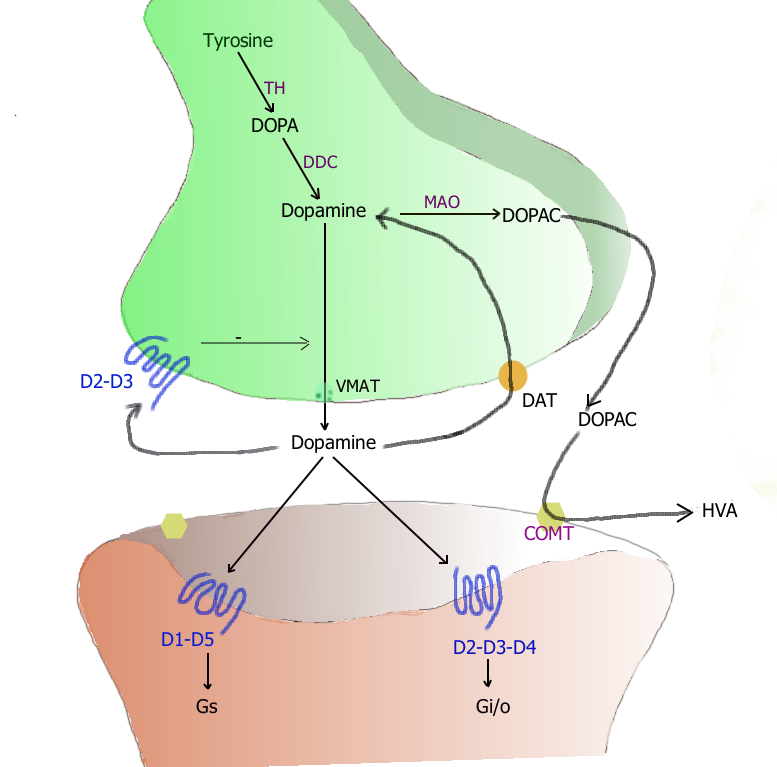
 Synapse dopaminergique (d'après Landry[4])
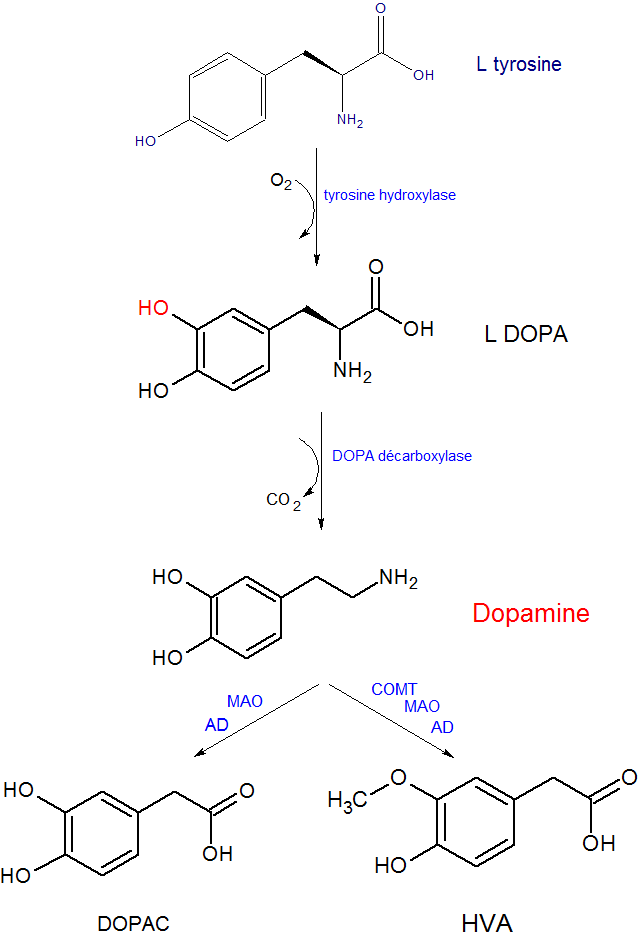
Synapse dopaminergique (d'après Landry[4])La dopamine est synthétisée dans les neurones à partir de la tyrosine d'origine circulante. La réaction est assurée par deux enzymes : la tyrosine hydroxylase (TH), une enzyme limitante contrôlant la production de L-DOPA, puis par la DOPA-décarboxylase (DDC)[4] assurant la décarboxylation de cette dernière pour donner la dopamine.
Après avoir été produite dans le cytoplasme des terminaisons présynaptiques, la dopamine est chargée dans des vésicules synaptiques par un transporteur VMAT-2. À l'arrivée d'un potentiel d'action, les vésicules libèrent leur contenu dans la fente synaptique par exocytose.
La dopamine libérée dans la fente est en partie captée par des récepteurs se trouvant sur la cellule postsynaptique et transmet ainsi le signal neuronal par transduction. Environ 80 % de la dopamine libérée est recaptée par les neurones dopaminergiques présynaptiques par des transporteurs sélectifs DAT (dopamine active transporter). Sauf au niveau du cortex préfrontal, où l'expression des DAT est très faible et où la dopamine est recaptée par les neurones noradrénergiques via le transporteur NET[5].
La dégradation de la dopamine s'opère soit dans la fente synaptique par une ectoenzyme, la catécholamine-O-méthyltransférase (COMT) soit à l'intérieur du neurone par des enzymes mitochondriales, les monoamine-oxydases A et B (MAO). La première voie produit de l'acide homovanillique (HVA) et la seconde, de l'acide dihydroxyphénylacétique (DOPAC). La mesure du taux de ces deux métabolites dans le liquide céphalorachidien sert à indiquer l'activité des neurones dopaminergiques centraux.
Les récepteurs dopaminergiques
On connait actuellement 5 types de récepteurs de la dopamine, codés par 5 gènes différents. Ce sont tous des récepteurs couplés aux protéines G (RCPG), formés par des protéines comportant 7 hélices transmembranaires. Ils sont classés en deux familles suivant la nature de la protéine G à la quelle ils sont couplés :
- la famille des récepteurs de type D1, comprenant les sous-types D1 et D5, couplés à la protéine Gs qui augmente l'adénylate cyclase
- la famille des récepteurs de type D2, comprenant les sous-types D2, D3, D4, couplés à la protéine Gi qui diminue l'adénylate cyclase
Contrairement aux récepteurs ionotropes, qui sont rapides, les récepteurs couplés aux protéines G réagissent lentement et le plus souvent ne produisent pas de courant postsynaptiques mesurables (du moins in vitro). Ils sont là pour moduler l'activité des neurones postsynaptiques et modifiant leur mode de traitement de l'information[6].
Localisation des récepteurs dopaminergiques (d'après Landry et Gies[4] 2009) Famille de type D1 Famille de type D2 D1 D5 D2 D3 D4 Protéine G Gs Gs Gi Gi Gi Localisation centrale striatum, noyau accumbens, tubercules olfactifs, cortex hippocampe, hypothalamus striatum, substance noire, noyau accumbens, tubercules olfactifs, cortex tubercules olfactifs, noyau accumbens amygdale, cortex, hippocampe Localisation périphérique artères, rein, tractus digestif artères, rein, tractus digestif terminaisons synaptiques, système nerveux entérique, area postrema, hypophyse rein, area postrema rein, coeur
Lorsque les récepteurs de sous-types D1 et D5 de la première famille sont activés par la liaison d'un agoniste, ils induisent la formation d'AMP cyclique par activation de l'adénylate cyclase par l'intermédiaire de la protéine Gs. La cascade de réactions qui s'en suit aboutit à une dépolarisation et donc à une augmentation de la fréquence d'émission des potentiels d'action.Ces récepteurs ont une localisation somato-dendritique dans le système nerveux central. Ils sont fortement exprimés par le striatum, le noyau accumbens, les tubercules olfactifs, le cortex cérébral, l'hypothalamus, le thalamus, et la pars reticulata de la substance noire. Le récepteur D5 dont l'affinité pour la dopamine est 10 fois supérieure à celle du récepteur D1 se trouve essentiellement dans l'hippocampe et l'hypothalamus.
Au niveau périphérique, l'activation des récepteurs de la famille D1, présents dans les cellules musculaires lisses vasculaires, entraînent une vasodilatation.
Les récepteurs dopaminergiques de type D2 comportent les sous-types D2, D3, D4 et leurs isoformes. Ils réagissent différemment à la liaison de la dopamine, puisque la protéine Gi à laquelle ils sont couplés induit une diminution du taux d'AMP cyclique et donc de l'activité PKA. Cette voie de signalisation aboutit à une hyperpolarisation au niveau postsynaptique et à une diminution de la libération de neuromédiateur (par inhibition de l'exocytose) au niveau présynaptique.
Au niveau central, ces récepteurs sont principalement localisés dans le striatum, la pars compacta de la substance noire, le noyau accumbens, les tubercules olfactifs et le cortex cérébral.
Les voies de projections dopaminergiques
Dans le système nerveux central, la dopamine joue un rôle complexe et intervient dans diverses fonctions importantes, telles que le comportement, la cognition, les fonctions motrices, la motivation, les récompenses, le sommeil ou la mémorisation. La dopamine est principalement synthétisée et libérée par des populations de neurones très restreintes situées dans la substance noire (SN) et dans l’aire tegmentale ventrale (VTA) qui ont pour cible majeure diverses structures cérébrales appartenant au système des noyaux de la base. Les noyaux de la base sont des structures sous-corticales comprenant différents noyaux (striatum, globus pallidus, noyau sous-thalamique). Le striatum qui représente le premier relais du système et la cible principale des projections dopaminergiques se divise en deux composantes: ventrale et dorsale.
De nombreuses études[7]ont mis en évidence trois grandes voies de projection ascendantes du mésencéphale.
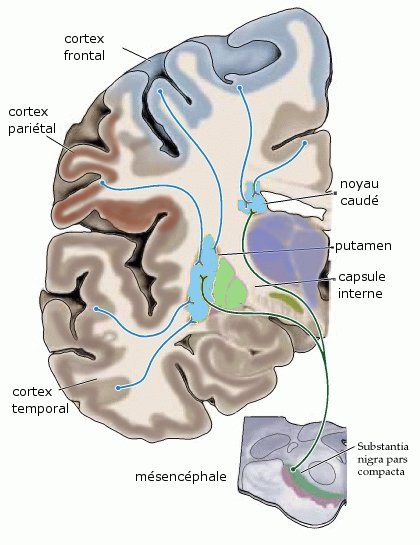
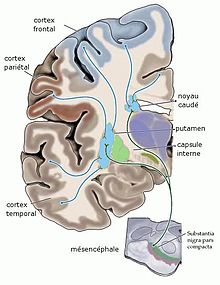 Voie nigrostriée dopaminergique, à la différence de la connexion cortico striée glutamatergique excitatrice (d'après Purves et al Neurosciences)
Voie nigrostriée dopaminergique, à la différence de la connexion cortico striée glutamatergique excitatrice (d'après Purves et al Neurosciences)1. La voie nigro-striatale ou nigrostriée
SN pars compacta → Striatum dorsal (N.C., putamen) Les neurones dopaminergiques de la pars compacta de la substance noire projettent majoritairement dans la partie supérieure du striatum, constituée du noyau caudé et du putamen. La voie nigrostriée représente 80 % des neurones dopaminergiques centraux[4]. Ce réseau constitue un système modulateur des aires corticales motrices et intervient dans le phénomène de contrôle des fonctions motrices[8]. La maladie de Parkinson est une maladie dont la cause est la dégénérescence du groupe de neurones produisant la dopamine dans la substance noire. L'administration du précurseur L-DOPA, qui contrairement à la dopamine peut traverser la barrière hémato-encéphalique, ralentit la progression de la maladie, car le cerveau transforme cette substance en dopamine.
2. La voie méso-limbique (mésencéphale → système limbique)
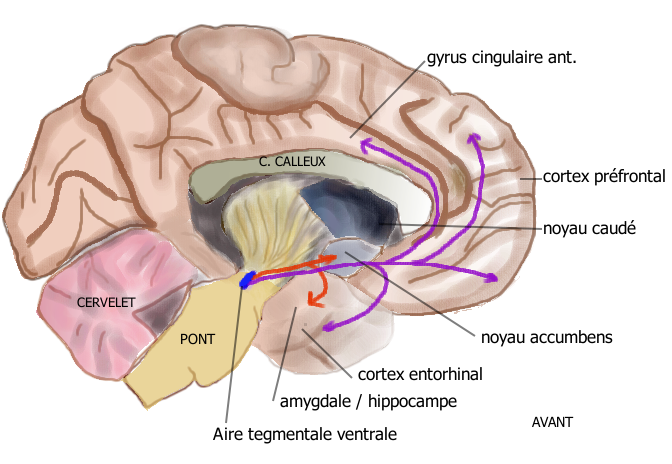
ATV → striatum ventral (N. Accumbens), septum, amygdale, hippocampe 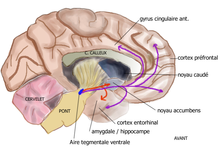 Voie mésolimbique (en rouge) et voie mésocorticale (en mauve), sur une coupe sagittale médiane du cerveau humain
Voie mésolimbique (en rouge) et voie mésocorticale (en mauve), sur une coupe sagittale médiane du cerveau humainLes projections dopaminergiques qui trouvent leur origine dans l'aire tegmentale ventrale (ATV) du mésencéphale ont pour cible la région ventrale du striatum, appelée également noyau accumbens ou nucleus accumbens, la stria terminalis, les tubercules olfactifs, le septum, l'amygdale, l'hippocampe. Ce réseau neuronal, qui représente le système de récompense / renforcement participe au contrôle des processus motivationnels et de récompense et est impliqué dans les phénomènes de dépendances et d'addiction[9]. Par exemple, la cocaïne provoque une augmentation de dopamine dans les fentes synaptiques au niveau du noyau accumbens en inversant le fonctionnement du système de recapture de la dopamine[10]. La nicotine provoque aussi une augmentation de la transmission dopaminergique. Toutes les substances psychoactives (cocaïne, amphétamine, alcool, opiacés) provoquent une augmentation des taux extracellulaires de dopamine dans le noyau accumbens (Di Chiara et al[11] 1988). D'une manière générale, son dysfonctionnement est lié aux comportement d'addiction. La dopamine est aussi impliquée dans la zone cérébrale non incluse dans la barrière hématoencéphalique responsable du réflexe de vomissement, ce qui explique l'effet anti-émétique des neuroleptiques (antagonistes dopaminergiques).
3. La voie mésocorticale
ATV → cortex frontal (gyrus cingulaire), temporal (gyrus entorhinal) Cette voie est formée de neurones dopaminergiques de l'aire tegmentale ventrale (ATV) dont les axones projettent sur le cortex frontal et ventral, en particulier sur le gyrus cingulaire antérieur, l'aire entorhinale et le cortex préfrontal. Elle joue un rôle dans la concentration et des fonctions exécutives comme la mémoire de travail[12].
Dans le système nerveux périphérique, elle joue le rôle d'analeptique circulatoire (stimulant des fonctions assurant la circulation sanguine).
La dopamine joue aussi un rôle de neuro-hormone au niveau de l'axe hypothalamo-hypophysaire.
La voie tubéro-infundibulaire a son origine dans l'hypothalamus et projette sur l'éminence médiane où la dopamine est relâchée dans la veine porte antéhypophysaire. Elle inhibe la libération de prolactine par l'hypophyse antérieure.La pratique régulière d'un sport permet d'augmenter la sécrétion naturelle de dopamine[13].
Le phénomène de frisson ressenti lors de l'écoute de musique est également dû à la sécrétion de dopamine[14].
Elle est aussi impliquée dans le trouble de déficit de l'attention/hyperactivité (TDAH), dont la cause est un problème de recapture de la dopamine par les synapses.[réf. nécessaire]
Agonistes et antagonistes
La dopamine ne passant pas la barrière hémato-encéphalique, n'exerce que des effets périphériques lorsqu'elle est administrée par voie générale.
L'apomorphine, un dérivé alcaloïdique de la morphine, sans action narcotique, est un puissant agoniste (activateur) des récepteurs dopaminergiques de type D1 et D2, qui a l'avantage de passer la barrière hémato-encéphalique. Elle est indiquée :
- dans le traitement de la maladie de Parkinson, par voie sous-cutanée. Aponikon® par perfusion sous-cutanée continue exerce une action antiparkinsonnienne en activant les récepteurs de type D2 striataux
- dans le traitement des dysfonctions érectiles, par voie sublinguale
- dans le traitement de certaines intoxications, comme émétique, par voie sous-cutanée (Aponikon®).Agonistes et antagonistes des récepteurs dopaminergiques (d'après Dunlop et al[5]) Famille de type D1 Famille de type D2 D1 D5 D2 D3 D4 Agoniste apomorphine, SKF38893,fénoldopan apomorphine, chloro-PB apomorphine, bromocriptine, pergolide, ropinirole, rotigotine apomorphine, cabergoline, pergolide, pramipexole apomorphine, PD 168077, quinpirole Antagoniste SCH 23390 SCH 23390 chlorpromazine, sulpiride chlorpromazine clozapine
L'antagoniste SCH 23390 a permis de distinguer expérimentalement l'activation des récepteurs de type D1 de ceux de type D2. Le chlorpromazine à l'inverse, est plus affine pour les récepteurs de type D2.La chlorpromazine (Largactil®) fut le premier médicament anti-psychotique, étudié et utilisé avec succès en 1951 par Henri Laborit.
Utilisation médicale
En perfusion continue, c'est un tonique qui accélère le rythme cardiaque. Elle permet de maintenir la pression artérielle et le débit cardiaque chez les patients dans certains cas états de choc.
À dose modérée, elle entraîne une vasoconstriction veineuse aidant au maintien de la pression artérielle. Elle entraîne également une vasodilatation des artères rénales permettant de conserver une diurèse.
À plus forte dose, les effets sur la contraction cardiaque sont contrebalancés par la survenue d'une tachycardie, voire de troubles du rythme cardiaque. Un dérivé, la dobutamine, présentant de manière atténuée ces effets secondaires, est alors préférentiellement utilisé.
À dose contrôlée, l'apport d'un agoniste de la dopamine améliore plusieurs maladies dont la maladie de Parkinson et le syndrome des jambes sans repos.
Traitements
Pour traiter des maladies à déficit dopaminergique, on a recours à plusieurs types de traitements.
Les molécules agissant sur le phénomène de recapture de la dopamine
On classe dans ce groupe les tricycliques, qui inhibent les transporteurs d'amines de la membrane pré-synaptique. Les tricycliques ne sont pas spécifiques du transporteur de dopamine (DAT), ils inhibent aussi les transporteurs de la sérotonine, la noradrénaline, etc...
Les molécules agissant sur le phénomène de dégradation synaptique de la dopamine
On classe dans ce groupe les IMAO, inhibiteurs de la monoamine oxydase (MAO).
- 2007. L'étude des mésanges charbonnières vient encore une fois confirmer les études de Bart Kempenaers, qui évoquait le fait qu'une augmentation de concentration en récepteur D4 de la dopamine dans le cerveau allait de pair avec un accroissement de curiosité chez l'animal[15].
Notes et références
- Masse molaire calculée d’après Atomic weights of the elements 2007 sur www.chem.qmul.ac.uk.
- Fallon J.H. and Moore R.Y. (1978), Catecholamine innervation of the basal forebrain. IV. Topography of the dopamine projection to the basal forebrain and neostriatum, J. Comp. Neurol., Volume 180, Issue 3, p. 545-80
- Abrous, Nora et als, « Tabac : comprendre la dépendance pour agir
chapr 5 : Effets de la nicotine sur la neurotransmission cérébrale », dans INSTITUT NATIONAL DE LA SANTE ET DE LA RECHERCHE MEDICALE, janv 2004 [texte intégral] - Yves Landry, Jean-Pierre Gies, Pharmacologie Des cibles vers l'indication thérapeutique, Dunod, 2009 (2e édition)
- Dunlop BW, Nemeroff CB., « The role of dopamine in the pathophysiology of depression. », dans Arch Gen Psychiatry., vol. 64, no 3, 2007, p. 327-37 [texte intégral]
- Missale C, Nash SR, Robinson SW, Jaber M, Caron MG., « Dopamine receptors: from structure to function. », dans Physiol Rev., vol. 78, no 1, 1998, p. 189-225
- Bertha Madras, C. Colvis, J.D. Pollock, JL Rutter, D. Shurtleff, M. von Zastrow, The cell biology of addiction, John Inglis, 2006
- Houk J.C. and Wise S.P. (1995), Distributed modular architectures linking basal ganglia, cerebellum, and cerebral cortex: their role in planning and controlling action, Cereb. Cortex. 5, 95-110.
- Nestler E.J. (2001), Molecular basis of long-term plasticity underlying addiction, Nat. Rev. Neurosci. 2, 119-128.
- http://www.lanutrition.fr/A-quoi-sert-la-dopamine-a-2003-152.html La Nutrition.fr : A quoi sert la dopamine ?
- DICHIARA G, IMPERATO A., « Drugs abused by humans preferentially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats », dans Proc Natl Acad Sci USA, vol. 85, 1988
- Charles B. Nemeroff, « The role of dopamine in pathophysiology of depression », dans Arch Gen Psychiatry, vol. 64, 2007
- Santé Magazine : Vous êtes fatigué ? Bougez !
- Pourquoi frissonne-t-on parfois en écoutant de la musique ? sur maxisciences.com. Consulté le 11 janvier 2011
- Drd4 gene polymorphisms are associated with personality variation in a passerine bird (Proc Biol Sci. 2007 July 22; 274(1619): 1685–1691)
Voir aussi
Catégories :- Produit chimique irritant
- Hormone de l'hypothalamus
- Neurotransmetteur
- Catécholamine
- Monoamine


Wikimedia Foundation. 2010.