- Agrobacterium tumefaciens
-
 Agrobacterium tumefaciens
Agrobacterium tumefaciens
Galle du collet provoquée par A. tumefaciens Classification Règne Bacteria Embranchement Proteobacteria Classe Alpha Proteobacteria Ordre Rhizobiales Famille Rhizobiaceae Genre Agrobacterium Nom binominal Agrobacterium tumefaciens
(Smith & Townsend 1907) Conn 1942Agrobacterium tumefaciens est une bactérie gram négative trouvée dans les sols. C'est un pathogène des végétaux responsable d'une maladie appelée galle du collet (ou crown-gall en anglais). A. tumefaciens a été identifiée à partir de galles en 1907[1].
Le mécanisme de formation des galles s'apparente à une transformation génétique, il est dû à un plasmide bactérien appelé plasmide Ti qui rend les bactéries virulentes. Un fragment d'ADN du plasmide Ti, l'ADN-T est transféré de la bactérie vers la plante, puis intégré dans le génome végétal où il induit la formations des galles caractérisés par la multiplication anarchique des cellules végétales. Agrobacterium tumefaciens infecte essentiellement les dicotylédones à la faveur d'une blessure.
Cette observation constitue le fondement d'une des techniques les plus utilisées en ingénierie génétique des végétaux pour produire des organismes génétiquement modifiés.
Sommaire
Génome
Le génome d' Agrobacterium tumefaciens (souche de référence C58, séquencée en 2001) se compose de deux chromosomes, l'un linéaire de 2,8 Mb contenant une origine de réplication plasmidique et l'autre circulaire de 2 Mb portant une origine de réplication de type cori. A. tumefaciens C58 porte également deux plasmides AtC58 et TiC58 (le plasmide Ti) respectivement de 540 Kb et 214 Kb[2].
Cycle infectieux
La bactérie Agrobacterium tumefaciens infecte les végétaux (essentiellement des dicotylédones) à la faveur d'une blessure. Des composés phénoliques produits par la plante - généralement bactériostatiques ou antibiotiques - attirent au contraire Agrobacterium vers le site de la blessure. La bactérie s'attache aux cellules végétales. Sous l'action de ces composés phénoliques, Agrobacterium met en place un système de transfert d'un fragment de son ADN, vers la cellule blessée. Cet ADN, dit ADN-T (ou T-DNA en anglais), est porté par le plasmide Ti (tumor-inducing) et s'intègre au génome nucléaire de la cellule végétale. Les gènes portés par l'ADN-T s'expriment dans le végétal et conduisent:
- d’une part à la synthèse d'hormones de croissance végétales, l'auxine, et une cytokinine, dont la surproduction entraîne une multiplication anarchique des cellules végétales, d'où formation de la tumeur.
- d'autre part, à la synthèse par la cellule végétale de composés absents habituellement de ces cellules, appelés opines.
Les opines sont spécifiquement utilisées par les agrobactéries qui ont induit la formation de la tumeur. Cette spécificité est liée au fait que les gènes déterminant l'utilisation des opines sont portés par le plasmide Ti. De plus, certaines opines induisent le transfert du plasmide Ti d'une agrobactérie vers une autre par conjugaison. Les opines sont donc des médiateurs chimiques clefs de l'interaction Agrobacterium - plante, dont la présence dans la tumeur favorise la croissance des pathogènes et concourt à leur dissémination.
L'interaction Agrobacterium - plante peut donc être vue comme une manipulation génétique naturelle au cours de laquelle la bactérie détourne à son profit l'activité métabolique de la plante.Étapes de la transfection
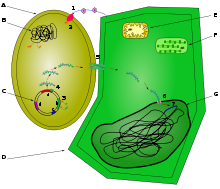 A: Agrobacterium tumefaciens
A: Agrobacterium tumefaciens
B: chromosomes d'Agrobacterium
C: Plasmide Ti : a: T-DNA , b: genes vir , c: origine de réplication , d: génes du catabolisme des opines
D: cellule végétale
E: Mitochondrie
F: Chloroplaste
G: NoyauLa bactérie perçoit les signaux phénoliques émis par la plante grâce à une protéine transmembranaire codée par le gène virA (1). La protéine VirA, à activité kinase, s'autophosphoryle et transfère le phosphate à une autre protéine, cytosolique cette fois et codée par virG (2). Cette dernière active la transcription des gènes de virulence (3).
La plante blessée émet également des signaux glucidiques, qui sont captés par une protéine codée par chvE. Celle-ci active VirA et la rend réceptive à des concentrations phénoliques faibles. L'augmentation du pH du milieu est captée par ChvI, qui active également les gènes de virulence.
Les gènes "vir" synthétisent plusieurs protéines dont une, VirD2, reconnaît les séquences double brin spécifiques des bordures droite et gauche de l'ADN-T. VirD1 ouvre les doubles brins, qui sont restreints (coupés) par VirD2; l'ADN-T est donc excisé du plasmide (4). La protéine VirD2 reste fixée sur la bordure droite de l'ADN-T, ce qui l'oriente pour sortir de la bactérie. Cette sécretion s'effectue par un pilus protéique reliant la bactérie à la cellule végétale (5). Cet appareil sécrétoire (dit de type 4) est essentiellement codé par les gènes de l'opéron virB, et par virD4 Avant ou après son entrée dans la cellule (selon les auteurs), l'ADN-T est recouvert de plusieurs monomères de la protéine VirE2. Celle-ci est également d'origine bactérienne, codée par le gène virE2, et permet de protéger l'ADN-T monocaténaire de la dégradation dans la cellule végétale. VirD2 et VirE2 possèdent des séquences signal de localisation nucléaire dites NLS (pour Nuclear Localisation Signal) qui sont reconnues par la cellule végétale, permettant d'adresser le complexe nucléoprotéique ADN-T/VirE2/VirD2 vers le noyau de la cellule végétale (6). Ce complexe pénètre donc dans le noyau, où il s'intègre à l'ADN végétal. Les ADN polymérases du végétal l'utilisent comme matrice, et le transforment en ADN double brin (7).
Les gènes codés par l'ADN-T ne sont généralement pas exprimés dans une cellule procaryote, donc chez Agrobacterium. Le génie génétique végétal repose en grande partie sur l'utilisation d' Agrobacterium comme vecteur naturel de gènes. Les biotechnologistes utilisent le plus souvent des plasmides proches du plasmide Ti, dit plasmides désarmés, car leur ADN-T ne porte plus les gènes responsables du pouvoir pathogène. Sur ces plasmides désarmés, les gènes tumoraux sont en effet remplacés par un (ou plusieurs) gène d’intérêt agronomique (par exemple le gène codant la toxine de Bacillus thuringiensis dit gène Bt, ou des gènes de résistance à l'herbicide non sélectif glyphosate) et par un ou plusieurs gènes marqueurs permettant de sélectionner les cellules transformées, puis de les multiplier sur des milieux de culture, in vitro. Des plantes entières sont ensuite régénérées par des techniques classiques de culture in vitro, faisant intervenir les hormones végétales auxine et cytokinine.
Lien avec LUCA
Article détaillé : Acidocalcisome.Une étude réalisée en 2003 montre que Agrobacterium tumefaciens présente les caractéristiques d'un acidocalcisome dans sa structure, organite que l'on pensait n'exister que chez les archées et les eukaryotes. Cette découverte est la première à montrer une caractéristique commune à tous les règnes du vivant.
Notes et références
- Smith EF, Townsend CO, « A PLANT-TUMOR OF BACTERIAL ORIGIN », dans Science, vol. 25, no 643, avril 1907, p. 671–3 [lien PMID, lien DOI]
- Goodner B, Hinkle G, Gattung S, et al., « Genome sequence of the plant pathogen and biotechnology agent Agrobacterium tumefaciens C58 », dans Science, vol. 294, no 5550, décembre 2001, p. 2323–8 [lien PMID, lien DOI]
Voir aussi
Liens externes
- Référence Catalogue of Life : Agrobacterium tumefaciens (en)
- Référence NCBI : Agrobacterium tumefaciens (en)
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire Portail de la microbiologie
Portail de la microbiologie Portail de la botanique
Portail de la botanique
Catégories :- Bactérie (nom scientifique)
- Rhizobiales
- Biotechnologie
- Génétique
- Gram négatif
- Bactérie phytopathogène


Wikimedia Foundation. 2010.