- Auxine
-
L’auxine est, en physiologie végétale, une phytohormone de croissance végétale qui est indispensable au développement des plantes. Le terme d’auxine a été étendu à un ensemble de substances naturelles aux propriétés analogues ainsi qu’à des hormones de synthèse.
Le terme a été formé sur le grec ancien auxein, croître.
Sommaire
Histoire
Cette découverte avait été pressentie par Francis Darwin aidé de son père en 1880 en observant la courbure des coléoptiles d’avoine vers la lumière (phototropisme). Couper le bout du coléoptile ou le recouvrir par un capuchon opaque empêchait toute courbure. En 1910, Boysen-Jensen sépare le bout du coléoptile de la tige par un bloc de gélose. Le coléoptile se courbant toujours vers la lumière, il en déduit que la substance induisant la courbure a migré à travers le bloc. En 1919 Paal puis en 1923 Soding réalisent des expériences complémentaires. Ils reprennent l'idée de Boysen-Jensen en remplaçant le bloc de gélose par des blocs de mica et de platine puis par du beurre de cacao. La plante ne se courbant pas vers la lumière, ils en déduisent que le signal n'est ni électrique ni liposoluble mais hydrosoluble.
La molécule d’auxine naturelle fut finalement découverte en 1926 par Went. Il dépose des bouts de coléoptiles sur des blocs de gélose afin que l'auxine migre dans ceux-ci. Il s'assure de la migration en déposant ces blocs sur l'un des côtés du bout de tiges qu'il a préalablement décapitées. Malgré une obscurité totale, ces tiges se courbent par une croissance du côté ou le bloc a été déposé. Enfin la structure chimique de l'auxine est définitivement identifiée en 1931 par Kogl et Haagen-smit.
Elle fut la seconde hormone végétale à être caractérisée (elle succède aux gibbérellines).
Biosynthèse
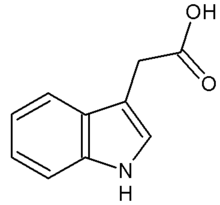 Acide indole 3-acétique.
Acide indole 3-acétique.
Sur le plan chimique, c’est l’acide indole 3-acétique ou AIA, un acide faible qui peut facilement se dissocier.
L’auxine est synthétisée majoritairement à partir du tryptophane mais aussi à partir du chorismate à l’extrémité des tiges (dans l’apex), et dans le méristème des bourgeons terminaux.
Synthèse à partir du tryptophane
Il y a deux voies de biosynthèse comportant chacune deux étapes.
Première possibilité :
- décarboxylation : tryptophane → tryptamine + CO2 (tryptophane décarboxylase)
- désamination : tryptamine + O → acide indole-acétaldéhyde + NH3 (tryptamine oxydase)
Deuxième possibilité :
- désamination : tryptophane + O → acide indole-pyruvique + NH3 (transaminase)
- décarboxylation : acide indole-pyruvique → acide indole-acétaldéhyde + CO2 (indole-pyruvique décarboxylase)
Puis l’acide indole-acétaldéhyde est transformé en acide indole-acétique (AIA, auxine) (indole-acétaldéhyde déshydrogénase)
Synthèse à partir du chorismate
Le chorismate est transformé en acide indole-pyruvique, puis en acide indole-acétique comme dans la voie de biosynthèse à partir du tryptophane.
Effets
 Plante sauvage (gauche), mutant au niveau de la transduction de signaux d’auxine (axr2, droite) (Gray, 2004).
Plante sauvage (gauche), mutant au niveau de la transduction de signaux d’auxine (axr2, droite) (Gray, 2004).Les rôles de l’auxine sont nombreux. Son action dépend très fortement à la fois de sa concentration et du tissu sur lequel elle agit. Par exemple, une même concentration peut inhiber le développement d’un bourgeon alors qu’elle favorisera l’élongation d’une tige.
Selon les plantes, une même concentration sur un même organe peut entraîner des conséquences différentes. Par exemple, l’auxine stimule la croissance du limbe des monocotylédones alors qu’elle inhibe celle des dicotylédones.
L’auxine a aussi une action cambiostimulante et est responsable du phototropisme.
Élongation cellulaire
Elle favorise la croissance en longueur en agissant sur l’élongation cellulaire ou auxèse.
Ce mode d’action de l’auxine est relativement bien connu.
L’auxine ne pénètre pas dans les cellules mais agit via une protéine G qui par l’activation de l’adénylate cyclase fait augmenter la concentration intracellulaire en ions Ca2+ et en AMPc ce qui active une cascade de réactions (notamment celle des MAP, Mitogen Activating Protein) stimulant la division cellulaire mais aussi l’activation et la production de pompes à protons. Ces pompes expulsent des protons dans le milieu extracellulaire ce qui fait diminuer le pH dans la paroi et augmenter le potentiel de membrane (la tension entre les deux côtés de la membrane est plus forte). Il s’agit de l’hypothèse « de la croissance acidodépendante ».
L’acidification de la paroi a pour conséquence d’activer les expansines (enzymes) qui coupent les liaisons hydrogène entre les microfibrilles de cellulose et d’autres composants de la paroi cellulaire. L’armature de cellulose se relâche et les polysaccharides de connexion sont séparés, grâce aux enzymes de la paroi cellulaire qui peuvent y accéder plus facilement. La paroi devient plus extensible. L’efflux des protons favorise aussi l’entrée d’ions potassium qui vont, par un mécanisme d’osmose, induire l’entrée d’eau dans la cellule, d’où une augmentation de la pression de turgescence qui s’applique sur la totalité de la paroi par l’intermédiaire du cytoplasme. La cellule peut alors « s’étirer ».
Tous ces facteurs concourent à l’allongement cellulaire lorsque la pression de la turgescence dépasse l’élasticité de la paroi végétale.
Phototropisme
L’auxine joue également un rôle dans le phototropisme positif des tiges. Un éclairement dissymétrique de la tige entraine une migration latérale de l’auxine du côté éclairé vers le côté sombre. Celle-ci favorisant la croissance, le côté sombre grandit plus vite et la tige se tourne alors vers la lumière, d’où le qualificatif de phototropisme positif.
Contrôle de la dominance apicale
En synergie avec les cytokinines, elle participe à la néoformation des bourgeons. En revanche elle s’oppose à leur débourrement : c’est le principe de la dominance apicale.
Le bourgeon apical profite de sa position haute pour dominer les bourgeons latéraux. Il synthétise de l’auxine (apex) qu’il évacue via le phloème. Les bourgeons sous-jacents subissent alors des concentrations en auxine trop fortes qui sont inhibitrices.
Formation des racines latérales
L’auxine a aussi des rôles dans l’organogénèse. Elle agit aussi à forte concentration (de l’ordre de 10-5 g/L) sur la rhizogenèse, favorisant l’apparition de racines sur les boutures.
Une forte concentration en auxine permet la mise en route des gènes impliqués dans l’initiation des méristèmes racinaires latéraux. Toutefois, si la teneur en auxine reste forte, la croissance racinaire sera ralentie.
Ainsi, une faible concentration en auxine favorise l’élongation d’une tige alors qu’une plus forte concentration inhibe cette croissance et favorise cette fois la rhizogenèse.
Développement des fruits
Si on retire une partie des akènes en développement alors le fruit se développe mal. L’ajout d’auxine rétablit un développement normal du fruit. Exemple : la fraise.
L’action de l’auxine est très importante dans l’induction florale.
L’action de cette hormone issue des apex, bourgeons terminaux, est contrecarrée par les gibbérellines venues des racines.
Un rameau fructifère est un végétal où l’action de l’auxine est suffisamment forte pour produire l’induction florale ou pour l’avoir produite. Le cas du greffon de pêcher qui fleurit l’année de la greffe s’explique ainsi.
Les plants de semis, vigoureux, tout en bois et racines, ont une physiologie où dominent gibbérellines et cytokinines.
Utilisation
L’auxine de synthèse la plus utilisée est l’acide naphtalène acétique (ANA). L’utilisation principale est le traitement local des boutures, on la trouve sous la dénomination hormone de bouturage. Elle a remplacé petit à petit l'utilisation de l'eau de saule.
L’auxine a un rôle fondamental dans les biotechnologies végétales, elle permet par exemple le développement des fruits sans fécondation (comme la banane qui est un fruit parthénocarpique).

Wikimedia Foundation. 2010.