- Diatomée
-
Bacillariophyta
Bacillariophyta
ou Diatomées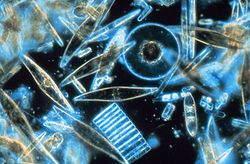
Diatomées marines vues au microscope Classification classique Domaine Eukaryota Règne Chromalveolata Division Stramenopiles Embranchement Ochrophyta Sous-embr. Diatomeae Super-classe Bacillariophyta
Engl. & Gilg, 1924Classes de rang inférieur Position phylogénétique - Opalinata
- Labyrinthulomycetes
- Bicosoecida
- Sloomycetes
- Hyphochytriomycetes
- Oomycota
- Ochrophyta
- Diatomeae
- Bolidophyceae
- Bacillariophyta
- Parmophyceae
- Hypogyristea
- Chrysista
- Diatomeae
Groupe frère : Bolidophyceae Parcourez la biologie sur Wikipédia : 





Les Bacillariophyta (diatomées) sont des microalgues unicellulaires planctoniques (de 2 μm à 1 mm) présentes dans tous les milieux aquatiques (avec une préférence pour les eaux froides) et enveloppées par un squelette externe siliceux. Elles peuvent vivre isolées ou en colonie, être libres ou fixées. Les formes pélagiques appartiennent au phytoplancton, les formes benthiques appartiennent au microphytobenthos. Les diatomées sont un constituant majeur du phytoplancton et jouent donc un rôle primordial dans la vie des écosystèmes marins, à l'origine des réseaux alimentaires de nombreuses espèces.
Sommaire
Description
 Différentes sortes de diatomées.
Différentes sortes de diatomées.
Les diatomées sont des algues jaunes et brunes unicellulaires caractérisées par le fait qu'elles sont les seuls organismes unicellulaires à posséder une structure externe siliceuse enveloppant totalement la cellule. Transparente et rigide, cette enveloppe, appelée frustule, est associée à des composants organiques et est formée de deux thèques emboîtées à symétrie remarquable ; la silice qui la compose est faiblement cristallisée (semblable à du verre).
Comme toute cellule végétale, la cellule d'une diatomée possède des chloroplastes pour sa photosynthèse. Les échanges avec le milieu extérieur se font aux travers de nombreux orifices très fins qui traversent la frustule et qui sont disposés en lignes (droites ou courbes) ou en réseau selon un motif propre à l'espèce. Ces ornementations sont ainsi utilisées pour la classification.
Le frustule
Le frustule est composé de deux parties ou thèques s'emboîtant l'une dans l'autre, à la manière d'une boîte. Chacune de ces deux parties est elle-même composée de deux éléments. Le premier, la valve, plus ou moins bombée (rarement plate), correspond à la face du « couvercle » ou du « fond » de la « boîte » ; à sa périphérie peut se situer une zone oblique ou verticale, le manteau, qui fait la liaison avec le second élément. Ce second élement, le cingulum, est une paroi verticale entourant la valve qui peut être formé d'une simple bande siliceuse ou comporter plusieurs bandes ou segments cingulaires. On parle d'épithèque ou d'épivalve ou d'hypovalve pour désigner la partie du frustule correspondant au « couvercle » et d'hypothèque ou d'épicingulum ou d'hypocingulum pour désigner le « fond ». Le volume interne du frustule, occupé par la cellule, peut être partiellement subdivisé par des cloisons, perforées ou incomplètes, portés par les valves ou par les bandes cingulaires.
Chez de nombreuses diatommées pennales, une fente, de longueur variable, parcourant les deux valves ou une seule, souvent en leur milieu, est appelée raphé et constitue un canal de communication avec l'extérieur et sert à la locomotion par excrétion du mucilage. Le raphé est interrompu en son milieu par un épaississement siliceux, le nodule central, et possède un nodule terminal à chaque extrémité. Si le raphé est en position médiane, il est situé dans une zone sans ornementations, l'aire longitudinale. Le nodule central est, lui, localisé dans l'aire centrale. Les diatomées dites monoraphidées sont dépourvues de raphé sur la valve supérieure et présentent une ornementation différente sur les deux valves. Les diatomées centrales[1] et certaines diatomées pennales sont dites araphidées car elles ne possèdent aucun raphé.
C'est notamment dans les familles des Surirellaceae et des Nitzschiaceae que l'évolution du raphé a abouti à la création d'une structure en forme de tuyau, le canal-raphé. À l'opposé du côté où s'ouvre le raphé, la paroi du tube est percée d'orifices débouchant à l'intérieur du frustule (pores internes) et séparés par des piliers siliceux, les fibules. Souvent, le canal-raphé est situé au faîte d'une carène valvaire plus ou moins développée.
 Plusieurs exemples de frustule de diatomées.
Plusieurs exemples de frustule de diatomées. Biddulphia pulchella, S. F. Gray, 1821. Diatomée des eaux chaudes à tempérées, présente en Méditerranée
Biddulphia pulchella, S. F. Gray, 1821. Diatomée des eaux chaudes à tempérées, présente en Méditerranée Cocconeis krammerii
Cocconeis krammeriiLa paroi des valves est constituée d'une seule couche siliceuse ou de deux couches superposées entre lesquelles se trouvent des espaces délimités par des cloisons transversales. Les diatomées pennales tiennent leur nom du fait que les ornements de leurs valves sont souvent disposés à la manière des dents d'un peigne alors qu'ils sont souvent radiaux chez les diatomées centrales. Ces ornementations sont propres à chaque espèce et sont donc gouvernés par les gènes. Les bandes cingulaires sont souvent dépourvues d'ornements. Ces ornementations de la frustule, qui apparaissent comme des stries, des côtes, des perles ou une dentelle, présentent une esthétique d'une extrême finesse dont la beauté attire de nombreux diatomistes.
Ces « ornements » correspondent en fait à des regroupements de fines perforations ou aréoles avec un diamètre de l'ordre du micromètre. Ces perforations ont souvent une extrémité (du côté soit externe soit interne de la paroi valvaire), partiellement obturé par une fine dentelle siliceuse qualifiée de crible. Ces orifices servent aux échanges entre la cellule et le milieu extérieur. Chez certaines diatomées, il existe des pores très fins (porelles) regroupés dans des structures limitées par un anneau marginal épaissi, les ocelles. Les diatomées pennales peuvent posséder des pores de plus gros diamètre à l'extrémité des valves permettant l'écoulement de substances mucilagineuses secrétées par la cellule.
À la surface des valves se trouvent de petites structures silicifées tubulaires : les « processus ». Il en existe quatre ou cinq types. C'est à leur niveau que certaines espèces extrudent des filaments de chitine.
- ↑ Les diatomées centrales sont parfois appelées diatomées centriques.
Cytologie
Les diatomées sont des plantes unicellulaires. Elles sont donc autotrophes et utilisent l'énergie de la lumière grâce à la chlorophylle a et la chlorophylle c contenues dans leurs chloroplastes par photosynthèse. Les chlorophylles sont situés dans les chloroplastes, qui peuvent être présentés comme de petites granules dispersés dans la cellule ou avoir une forme de plaque ou de bande ou d'étoile et sont d'un nombre variable (entre un et plusieurs dizaines). Par la présence de pigments de carotènes et de xanthophylles (dont la fucoxanthine), les chloroplastes sont jaunes, verts olive ou bruns.
Certaines espèces peuvent malgré tout présenter une hétérotrophie plus ou moins importante. Celle-ci peut être facultative et temporaire lorsque la diatomée est en présence de molécules organiques assimilables et que l'éclairement est insuffisant. Quelques rares espèces (notamment en eaux eutrophes et peu éclairées des zones littorales et estuariennes) ne sont cependant pas douées de photosynthèse et ont donc une hétérotrophie obligatoire.
Comme toute cellule, les diatomées possèdent une membrane organique périphérique à leur cytoplasme, un noyau, et les différents organites nécessaires à son fonctionnement (mitochondries, systèmes membranaires et tubulaires intracytoplasmique… ). La cellule des diatomées centrales est en partie occupée par une vacuole dont la densité du volume est proche de celle de l'eau ambiante. La plupart des diatomées pennales possèdent deux vacuoles situées de part et d'autre de la zone cytoplasmique médiane entourant le noyau. Des globules lipidiques sont en outre présents en nombre variable, ceux-ci servent de réserve mais peuvent aussi refléter un mauvais état physiologique de la cellule ; les lipides représentent généralement 4 à 8 % du poids sec mais peuvent parfois en représenter 40 % chez certaines espèces ou dans certaines situations.
Les diatomées excrètent des substances mucilagineuses à travers des pores spéciaux. Ces excrétions jouent un rôle important dans leur locomotion, leur flottaison (pour les espèces planctoniques), leur reproduction et dans la constitution et la protection des colonies. Certaines peuvent émettre des filaments de chitine. Les diatomées peuvent parfois secréter des molécules susceptibles d'agir sur d'autres organismes (bactéricides, antibiotiques, substances toxiques pour les œufs d'invertébrés marins, voir des toxines nocives pour l'Homme).
Locomotion
À moins de posséder un raphé sur la valve en contact avec le substrat, les diatomées pennales peuvent se mouvoir de manière autonome. Elles sont notamment attirées par la lumière sauf si cette dernière est trop intense. Les mécanismes permettant cette locomotion ne sont pas totalement élucidés mais il semble néanmoins qu'une substance mucilagineuse adhère au substrat et s'écoule à travers le raphé dans la direction opposée au mouvement. Les diatomées les plus rapides peuvent atteindre des vitesses de 20 μm/s soit 7,2 cm/h. Sans avant ni arrière, les diatomées inversent périodiquement le sens du mouvement généralement.
Les diatomées centrales n'ont pas de raphé et ne peuvent donc se déplacer sur un support.
Multiplication végétative
La multiplication cellulaire végétative est la principale méthode de multiplication des diatomées ; elle ne fait pas intervenir de processus sexué. Lorsque les conditions sont favorables à leur prolifération, les diatomées se multiplient par bipartition (la cellule mère donne deux cellules filles), ce qui peut se faire de manière très rapide.
Cette bipartition est particulière : chaque diatomée fille conserve une des thèques de la diatomée mère comme épithèque propre, et reconstitue la thèque manquante. Les deux thèques n'ayant pas la même taille, les diatomées filles sont de tailles différentes : celle qui est issue de la grande thèque a la même taille que la diatomée mère, tandis que celle issue de la petite thèque est légèrement plus petite. Ce processus se reproduit à chaque cycle.
Il y a d'abord formation de l'hypocingulum de chacune des deux nouvelles thèques. La division du noyau et du cytoplasme est suivie par la formation de deux hypovalves qui sont totalement silicifiées dans les dix à vingt minutes suivant la division. Une fois les deux frustules achevés, les deux cellules filles se séparent, ou restent associées chez les espèces coloniales.
La manière dont la paroi siliceuse est élaborée n'est pas complètement élucidée mais on peut schématiquement l'expliquer ainsi. La cellule extrait les traces d'acide silicique du milieu. Un système membranaire particulier forme des vésicules sous la membrane cellulaire dans lesquelles est polymérisé l'acide silicique et où la silice s'accumule. Leur nombre va croissant au fur et à mesure de la quantité de silice produite, les vésicules finissent par fusionner et les néoformations du frustule sont alors transférés à l'extérieur. À la fin de la formation du frustule, une partie du cytoplasme reste associé à la partie siliceuse pour former une mince couche organique, tandis que se reforme en dessous une nouvelle membrane cellulaire.
Reproduction
Lors de la multiplication asexuée, chaque nouvelle thèque des deux cellules filles étant une hypothèque, une des cellules filles est plus petite que la cellule mère induisant une diminution progressive de la taille des générations successives de diatomées (la taille est donc un critère d'identification à utiliser prudemment). À partir d'une taille minimale (30 % de la taille initiale), la reproduction intervient entre deux diatomées afin de générer des individus de taille normale, trois ou quatre fois, voire huit à dix fois plus grands. Le processus est complexe et diffère chez les diatomées centrales et les pennales.
Chez les diatomées centrales, les cellules se transforment les unes en gamètes femelles, les autres en gamètes mâles. Les gamètes mâles sont munis d'une flagelle avec laquelle il s'introduisent dans les diatomées gamètes femelles. Chez les diatomées pennales, la fécondation se fait par cystogamie : il y a d'abord adhésion des deux diatomées, facilitée par du mucilage. Chacune évolue alors en un ou deux gamètes actifs (mâles, sans flagelle) ou passifs (femelles) qui fusionnent ensuite par divers moyens.
Dans tous les cas, l'œuf résultant de la fusion des gamètes, appelé auxospore, s'entoure d'une épaisse paroi mucilagineuse et grossit considérablement avant de secréter un frustule et donc devenir une nouvelle diatomée de grande taille.
Hypnospores et microspores
Si les conditions environnementales deviennent défavorables (diminution de l'éclairement, de la température, de la teneur en sels nutritifs… ), de nombreuses espèces de diatomées centrales (surtout planctoniques) et quelques diatomées pennales, forment des structures de résistance, les hypnospores ou spores de résistance, qui peuvent tenir un état de vie ralenti durant quelques semaines.
Ces structures sont formées par contraction du cytoplasme en une masse dense, de couleur foncée, qui secrète une membrane silicifiée, constituée de deux valves identiques ou différentes. Ces spores peuvent rester dans le frustule parental ou non et peuvent ressembler à la diatomée initiale ou avoir une morphologie différente. Lors de la germination, le spore réinvestit le frustule parental ou en secrète un nouveau.
Des cellules de très petite taille peuvent se former à l'intérieur du frustule de certaines diatomées centrales, au nombre de 8 à 128. La nature de ces microspores est inconnue et fait l'objet de plusieurs théories.
Colonie
De nombreuses espèces vivent isolées, soit libres soit fixées par du mucilage à un support (inerte ou vivant) mais il existe également des espèces coloniales aussi bien chez les diatomées centrales que pennales. À la suite de la multiplication végétative, les cellules filles restent associées, soit grâce à des substances mucilagineuses ou à des filaments de chitine, soit par l'intermédiaire de dents, d'épines ou de soies, afin de former progressivement une colonie. Selon l'espèce et le mode de liaison, la colonie peut prendre une multitude de formes (ruban, étoile, chaînette, éventail, « arbuste », zigzag ou tube muqueux à l'intérieur duquel les diatomées sont mobiles). Chaque diatomée de la colonie reste autonome et peut survivre si la colonie est fragmentée.
Plusieurs hypothèses sont avancées quant à l'avantage de ces colonies : influence sur la flottabilité des espèces planctoniques et sur l'absorption des sels nutritifs, défense contre le broutage et le zooplancton.
L'aptitude de certaines diatomées à former des constructions pluricellulaires à amener certains diatomistes du XIXe siècle (par exemple, Jean Deby en 1888) à proposer que les diatomées étaient fondamentalement des organismes pluricellulaires.
Écologie
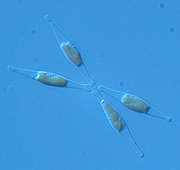 Phaeodactylum tricornutum
Phaeodactylum tricornutumIl existe environ 100 000 espèces connues de diatomées, mais d'après certains auteurs, il en existerait près d'un million. Elles sont répandues dans tous les milieux aquatiques (que ce soit en eau douce, salée ou saumâtre) ou même seulement humides et sont soit planctoniques (vivent en suspension dans le milieu liquide) soit benthiques (vivent sur le fond ou sur des supports variés). La mer contient de nombreuses espèces planctoniques de diatomées, isolées ou en colonie, et surtout centrales. En eau douce, elles sont essentiellement benthiques et sont plutôt des diatomées pennales et coloniales. En général, les espèces benthiques sont d'ailleurs souvent pennales.
La présence des diatomées dans un milieu est liée à plusieurs paramètres physico-chimiques (lumière, sels minéraux, mais aussi le pH, la salinité et les teneurs en oxygène et en matière organique). Elles sont surtout présentes dans les mers froides où des courants ascendants ramènent en surface les nitrates et les phosphates dissous, alors qu'elles se font plus rares dans les mers tropicales, pauvres en éléments minéraux. Elles abondent également dans les zones où ont lieu des remontés d'eau froides (« upwellings ») riches en sels minéraux. Des variations à long terme se superposent parfois aux variations saisonnières (entre deux et plusieurs années selon le lieu) dues à des fluctuations de la température, des courants et des paramètres chimiques de l'eau à cause des évolutions climatiques. Ces variations peuvent avoir des conséquences sur les population de poissons et donc sur les pêches.
Au printemps, sous l'effet de la température et de la chaleur, elles peuvent se développer de façon abondante formant des efflorescences dans les lacs. Les petites diatomées sont présentes surtout au printemps et en automne. Les diatomées plus grosses sont ubiquitaires (elles sont présentes dans tous les océans). Elles se déposent sur les fonds et objets immergés formant un amas, plus ou moins gélatineux, de couleur brune. En eaux douces, dormantes ou courant doucement, les diatomées centrales peuvent constituer d'épais entrelacs de longs et fins filaments bruns (à la différence des algues aquatiques, de couleur verte). Il existe des espèces capables de coloniser les milieux les plus inhospitaliers (mares temporaires, mousse humide sur les arbres, glace, effluents pollués). Une température de 0°C n'est pas mortelle tant que l'eau (salée) ne gèle pas.
Les diatomées ayant besoin de lumière, elles se situent près de la surface de l'eau. À la surface même, l'intensité lumineuse est trop élevée et de nombreuses espèces se limitent à une profondeur où seul un tiers de la lumière est filtrée (entre 3 et 40 mètres). Comme le reste du phytoplancton, chaque espèce de diatomée a un besoin d'énergie lumineuse pour compenser l'énergie dépensée pour survivre, lorsqu'une certaine profondeur dite de compensation est franchie, la cellule dépérit. Cette profondeur varie selon la nébulosité et les saisons. Malgré leur densité légèrement supérieure à celle de l'eau, les diatomées ne s'enfoncent pas car elles possèdent des mécanismes adaptatifs morphologiques et physiologiques pour faire varier leur densité (donc leur flottabilité). Les mécanismes morphologiques possibles peuvent être un amincissement de la frustule, l'allongement de sa forme en baguette fine, la présence de mucus, de soies, de filaments et d'épines qui jouent un rôle de parachute, voire l'organisation coloniale en rubans ou en chaînettes. Les mécanismes physiologiques influent sur le métabolisme : échanges sélectifs des ions (les ions chlorure et potassium sont plus légers que les ions sulfate et magnésium) et production de mucus dont la densité est plus faible que celle de la cellule.
La diversité remarquable des diatomées est d'ailleurs utilisée pour indiquer la qualité des cours d'eau : les diatomées constituent un excellent bioindicateur. Elles sont utilisées en routine dans tous les pays d'Europe, mais également aux États-Unis, au Canada, etc. En France, l'IPS -Indice de Polluosensibilité Spécifique- (Coste, 1982) ainsi que l'IBD -Indice Biologique Diatomées- (Afnor, 2000) sont utilisés. La pollution croissante de l'eau ainsi que l'utilisation intempestive d'engrais chimiques et d'herbicides ont transformé et appauvri les populations de diatomées (H. Germain, 1981). Si l'impact sur l'écosystème côtier n'est pas encore totalement saisi, il semblerait que les littoraux en subissent les conséquences par la modification de la composition alguale du plancton, provoquant des blooms de phytoplancton toxique. Les dinoflagellés sont les principaux concernés mais depuis quelque temps, certaines espèces de diatomées sont également mises en cause. La prolifération d'une espèce de diatomées, même si celle-ci n'est pas toxique, peut être responsable de mortalité élevée chez les invertébrés marins (entre autres dans les zones d'élevages conclylicoles) en consommant tout l'oxygène du milieu et surtout, à la fin du bloom, par l'accumulation et la décomposition de leur matière organique qui provoque une désoxygénation des eaux profondes.
Place actuelle dans la biosphère
Notamment par la masse qu'elles représentent, les diatomées constituent le groupe d'algues le plus important du phytoplancton marin (constitué par ailleurs de dinoflagellés, de cocolithophoridés et d'algues flagellés). Dans les mers et les océans, elles assurent 20 à 25 % de la production phytoplanctonique, jouant un rôle majeur dans la vie des écosystèmes marins. Elles sont à la base des réseaux alimentaires qui aboutissent aux poissons, aux crustacés et à plusieurs espèces de coquillage économiquement importants. À part quelques exceptions, ce sont typiquement des « algues fourrages », contrairement à d'autres algues comme les dinoflagellés qui sont toxiques pour les vertébrés et les invertébrés.
En effet, comme l'ensemble des éléments phytoplanctoniques, elles sont des producteurs de matière organique alors que les autres êtres vivants, dépourvus de chlorophylle, ne sont que des transformateurs de la matière produite par le phytoplancton. Au printemps, dans les eaux arctiques, la masse des diatomées est de 2 à 20 grammes par mètre cube d'eau. Chaque année, 200 à 600 grammes de carbone, sous forme de dioxyde de carbone, sont incorporés dans la matière vivante par mètre carré d'océan. La production annuelle de matière organique (sèche) dans les océans est estimée à environ 25 milliards de tonnes.
Fabriquants de la matière vivante à partir d'éléments minéraux, les diatomées sont à la base des chaînes et des réseaux alimentaires. Elles sont consommées par les animaux herbivores du plancton : petits crustacés (copépodes et euphausiacés), larves de crustacés, alevins de poissons, et quelques poissons ayant un régime herbivore (anchois, sardines… ). Ces espèces sont ensuite elles-mêmes consommées par des prédateurs supérieurs. En mer tempérée, on estime que pour 1 kg de plancton végétal consommé, 2,5 g de poisson carnivore est produit.
En plus de la production de matière organique, l'activité photosynthétique des diatomées et des autres algues phytoplanctoniques a deux effets importants sur le fonctionnement de la biosphère. D'une part, elle réduit le dyoxyde de carbone présent dans l'atmosphère en fixant une partie des 10 milliards de tonnes que les Hommes rejettent chaque année. Les diatomées fixent entre 25 et 50 % du carbone total sur Terre [réf. nécessaire]. En raison du rôle important des diatomées dans le cycle du carbone sur Terre, les génomes de Thalassiosira pseudonana et de Phaeodactylum tricornutum ont été séquencés. D'autre part, étant productrice d'oxygène, l'activité photosynthétique du phytoplancton favorise l'oxygénation des eaux et fait de l'océan une immense source d'oxygène. On estime qu'environ un tiers de l'oxygène produit par les végétaux sur Terre provient du phytoplancton.
L'importance des diatomées est grande également pour les espèces animales benthiques. En ces lieux, le plancton végétal constitue, avec les détritus organiques, les bactéries voir le plancton animal, une source de nourriture importante pour les espèces d'invertébrés filtreurs (moules, huîtres, balanes… ).
En mer comme en eau douce, les diatomées benthiques, associées souvent à d'autres espèces microscopiques, forment sur les roches et les plantes des revêtements organiques souvent minces et peu apparents dont de nombreuses espèces d'invertébrés brouteurs se nourrissent (oursins, bigorneaux, patelles …).
De récents développements dans la recherche sur l'influence des diatomées sur le réseau trophique montrent qu'elles ne se contentent pas d’être mangées par les crustacés et les autres animaux. Les diatomées ont développé de nombreux moyens de défense contre leurs prédateurs. Ces armes sont soit chimiques (production de composés toxiques à court ou long terme pour les crustacés) soit mécaniques. En effet, leur thèques en silice les rendrait moins ingérables pour le microplancton. La possibilité que les diatomées émettent des signaux chimiques en cas d’attaque est aussi à l'étude. Il est très probable qu'une cellule en train d’être mangée par un crustacé produise un signal pour que les autres diatomées puissent renforcer leurs défenses. Il existe au moins cinq espèces de diatomées marines toxiques pour l'Homme, celle-ci peuvent être responsables de troubles digestifs ou neurologiques (souvent ingérés en consommant des coquillages intoxiqués). Au moins deux de ces espèces prolifèrent de plus en plus sur les côtes françaises depuis 1999. La recherche sur les interactions entre les diatomées, et plus généralement tout le phytoplancton, avec leurs prédateurs est à ses débuts, mais cela promet de remettre en question la vision classique et parfois simpliste de l’écologie marine.
Classification
La dénomination « diatomée » a été retenue après que Augustin Pyrame de Candolle (1778-1841) eut, en 1805, baptisé Diatoma un genre de diatomée d'eau douce. En grec, diatomos signifie « coupé en deux ». De Candolle a pu faire allusion au fait que le frustule peut se dissocier en deux (ou plus exactement, l'aptitude de la diatomée mère à se diviser pour donner deux diatomées filles identiques). Les diatomées forment l'une des classes des algues jaunes et brunes (ou chromophytes), caractérisées par la présence de pigments associés à la chlorophylle qui donnent aux chloroplastes une couleur brune ou jaune.
Les premières classifications étaient fondées sur des observations au microscope photonique et furent depuis corrigées avec l'apparition du microscope électronique. En fait, avec la masse croissante d'informations, les diatomistes privilégient tels ou tels caractères et il existe encore beaucoup d'incertitudes sur leur classification : plusieurs systèmes de classification sont proposés actuellement (le plus récent date des années 1980-1990). Ainsi le nombre d'espèces et de genre est très imprécis, oscillant entre 5 000 et 12 000 pour les espèces, et entre 250 et 300 pour les genres.
Les diatomées sont classiquement divisées en deux ordres : les centrales ou centriques (ordre des Biddulphiales) et les pennales ou pennées (ordre des Bacillariales). Les diatomées centrales présentent une symétrie radiale et ont un frustule qui a la forme soit d'un disque, plus ou moins épais, soit d'un tube, cylindrique ou non. Les pennales présentent une symétrie bilatérale et un frustule plus ou moins allongé.
À partir de ces deux symétries de base (radiale ou bilatérale) s'ajoutent de nombreuses variantes parfois si marquées qu'elles masquent plus ou moins la morphologie de base. Ainsi, la forme en disque ou cylindre n'est pas toujours évidente chez les diatomées centrales et les diatomées pennales peuvent avoir une forme en croissant, en baguette, en S, en vrille, être ressérés au milieu ou avoir deux extrémités dissemblables (frustules hétéropolaires). Le raphé peut être plus ou moins excentré, faire le tour de chaque valve ou être très court et limité aux extrémités valvaires. Ces variations morphologiques au sein d'une même espèce peuvent être suffisamment importantes pour que les systématiciens les considèrent comme des variétés. Pour la même raison, de nombreux genres (comme Navicula par exemple) présentent un passage insensible d'une espèce à une autre.
La classification phylogénétique comprend les Bacillariophycidae et les Eunotiophycidae.
 Une diatomée centrale.
Une diatomée centrale.- Ordre des Biddulphiales (diatomées centrales)
- Sous-ordre des Coscinodiscineae, circulaires, aplaties ou peu allongées.
- Famille des Melosiraceae
- Famille des Thalassiosiraceae
- Famille des Coscinodiscaceae
- Famille des Hemidiscaceae
- Famille des Heliopeltaceae
- Sous-ordre des Rhizosoleniineae, forme de cylindre allongé
- Famille des Rhizosoleniaceae
- Sous-ordre des Biddulphiineae, formes variables
- Famille des Hemiaulaceae
- Famille des Chaetocerotaceae
- Famille des Biddulphiaceae
- Famille des Lithodesmiaceae
- Famille des Eupodiscaceae
- Sous-ordre des Coscinodiscineae, circulaires, aplaties ou peu allongées.
- Ordre des Bacillariales (diatomées pennales)
- Sous-ordre des Fragilariineae, absence totale de raphé
- Famille des Fragilariaceae
- Famille des Thalassionemataceae
- Sous-ordre des Bacillariineae, présence de raphé
- Famille des Eunotiaceae
- Famille des Achnanthaceae
- Famille des Naviculaceae
- Famille des Entomoneidaceae
- Famille des Cymbellaceae
- Famille des Auriculaceae
- Famille des Epithemiaceae
- Famille des Surirellaceae
- Famille des Nitzschiaceae
- Sous-ordre des Fragilariineae, absence totale de raphé
Apparition
Les diatomées sont apparues au cours de l'ère secondaire. Les plus anciens fossiles connus datent du début du jurassique (~185 millions d'années ; Kooistra & Medlin, 1996) bien que des récents faits génétiques (Kooistra & Medlin, 1996) et sédimentaires (Schieber, Krinsley & Riciputi, 2000) suggèrent une origine plus ancienne. Medlin et al. (1997) suggèrent que leur origine pourrait se rapporter à la fin du permien (extinction massive), après quoi de nombreuses niches écologiques marines furent ouvertes. Les principaux dépôts fossiles de diatomées remontent au début du Crétacé, un type de roche (appelé Kieselguhr) est composé presque entièrement de ceux-ci.
Les diatomées centrales sont apparues il y a 150 Ma, les pennales il y a 70 Ma et les pennales avec raphé 20 Ma plus tard. Elles sont devenues depuis les principales constituantes du plancton marin et d'eau douce.
Géologie
Les frustules de diatomées étant imputrescibles, leur prolifération intense et leur accumulation pendant des millions d'années ont formé des gisements parfois considérables de tourbe siliceuse ou de roches appelées diatomite (constituées à plus de 80 % par les frustules).
La diatomite (connue aussi sous le nom de tripoli, de farine fossile ou de Kieselguhr) est de couleur claire, tendre, légère et à porosité élevée. La diatomite possède un intérêt économique : elle est utilisée, une fois nettoyée et réduite en poudre, comme support dans certaines techniques de filtration et de clarification (raffinage du sucre, filtration du vin… ) ou les micropores des frustules servent de tamis ultrafin (0,7 à 2 microns). Elle est aussi utilisée comme adjuvant dans de très nombreux produits: peintures, bitumes, détergents, décolorants, désodorisants, engrais... Lors du grattage d'une allumette c'est grâce à l'ultrastructure microperforée des frustules de diatomées que les gaz issus de la combustion du souffre s'échappent sans que le bout de l'allumette n'explose! Nobel utilisa la diatomite pour stabiliser la nitroglycérine afin de fabriquer de la dynamite. De par leur faible densité, ces roches servirent à bâtir le dôme de la Cathédrale Sainte-Sophie à Constantinople en 532 (32 mètres de haut).
Ses poussières sont cancérigènes.
Les gisements les plus importants se situent aux États-Unis : ceux de l'ouest, d'origine marine, s'étendent de San Francisco jusqu'au sud de la Californie, ceux de l'est du Maryland à la Virginie. En France, des gisements d'origine d'eau douce, sont exploités en Auvergne et dans le Gard. Actuellement, des boues à diatomées, futures diatomites, se déposent dans les mers notamment en mer froide mais aussi, en plus faible quantité, dans les grands lacs comme le lac de Lugano, le lac Pavin ou le lac Léman.
Dans des conditions géologiques différentes, les diatomées ont également participé à la formation du pétrole, issu de la substance accumulée durant des millions d'années par des animaux qui vivaient dans la masse d'eau ainsi que celle des végétaux dont les diatomées constituaient une grande partie. Des gisements pétrolifères en Californie et en Roumanie ont ainsi une roche composée quasi-uniquement de frustules.
Diatomites et pétrole permettent de saisir combien les diatomées, malgré leur taille microscopique, ont joué un rôle non négligeable dans la mise en place des roches sédimentaires.
Recherches
Les premières études sur les diatomées remontent au XVIIIe siècle : en 1703, le chercheur hollandais Antoni van Leeuwenhoek (1632-1723), en utilisant l'un des premiers microscopes sur une goutte d'eau, découvre une structure que l'on a identifée a posteriori comme étant une diatomée mais qu'il prit pour un cristal. Pendant tout le XVIIIe siècle, les observations sur les diatomées se multiplièrent, laissant perplexes les observateurs quant à la nature végétale ou animale de ces organismes aquatiques. À la fin du siècle, une à deux dizaines de diatomées étaient connues.
En 1817, Christian Ludwig Nitzsch (1782-1837) publie le résultat de ses recherches approfondies sur les diatomées. Il observe notamment la multiplication des diatomées, mais les considère plutôt comme des animaux du fait de leur capacité à se mouvoir. En 1832, Christian Gottfried Ehrenberg (1795-1876) publie un ouvrage illustré de dessins très précis décrivant 144 espèces de diatomées. Durant la seconde moitié du XIXe siècle, de nombreuses nouvelles espèces sont découvertes et décrites, notamment grâce aux expéditions océnographiques dans les mers froides.
Au début du XXe siècle, 1 000 espèces sont connues rien que pour les eaux marines françaises et la classification des diatomées prend forme (H. et M. Peragallo, 1908). Cette classification s'affine ensuite pendant la seconde moitié du XXe siècle grâce à divers auteurs qui proposeront des classifications plus ou moins différentes les unes des autres. En même temps, l'écologie de ces cellules se développe. À partir de 1960, l'utilisation du microscope électronique à balayage permet d'observer de très près la surface des cellules, relançant l'étude de la structure ultrafine des frustules et fournissant de nouvelles bases pour une classification. Parallèlement, des études sur les molécules qu'elles produisent et sur leurs gènes sont développées soit dans l'optique de manipulations génétiques futures soit pour appuyer la classification sur des critères non plus morphologiques mais génétiques.
La nécessité d'optimiser les moyens d'observation des frustules des diatomées pour les diatomistes du XIXe siècle a été la principale cause des améliorations qu'opticiens et physiciens ont apportées à la construction du microscope, et surtout à la conception des objectifs. De ce point de vue, les diatomées ont joué un rôle primordial dans l'avancement de la microbiologie, permettant de disposer de matériels performants lorsque l'étude des microbes fut abordée.
Observer les diatomées
On dénombre aujourd'hui 100 000 espèces de Bacillariophytes, la majorité sont photosynthétiques. Par ailleurs, elles sont toujours caractéristiques de l'étendue d'eau dans laquelle elles ont été prélevées : deux échantillons de deux lacs différents seront forcément dissemblables l'un de l'autre.
Au microscope, selon sa position, une diatomée est vue soit de dessus (vue valvaire) soit de côté (vue connective).
Exploitations par l'Homme
En mytiliculture et en ostréiculture traditionnelles, les diatomées participent à la croissance des moules et des huîtres qui se nourrissent de celles que la mer contient. Des systèmes d'élevage intégré sont à l'étude pour en améliorer le rendement.
Certaines espèces de diatomées produisent des molécules intéressantes telles que des antibiotiques et des substances antitumorales (Haslea ostrearia par exemple). Elles produisent des acides gras nécessaires aux animaux et aux humains qui ne peuvent pas les produire eux-mêmes. Par ailleurs, la teneur en lipides de certaines espèces en fait une source potentielle de biodiesel. Leurs structures régulières hyperfines permettent d'envisager des exploitation futures dans la nanotechnologie.
Les diatomées répondant à la plupart des paramètres physico-chimiques et selon des paramètres de chaque espèce, elles sont de très bons indicateurs de la qualité des eaux. En France, un Indice biologique diatomées, (IDB), prenant en compte 209 espèces benthiques a été élaboré en 1994 par les Agences de l'eau et le Cemagref.
Dans le domaine de la médecine légale, les diatomées sont utilisées quand une victime est retrouvée dans un cours d'eau ou un lac pour déterminer s'il y a eu noyade ou immersion du corps après le décès.
La calcination de diatomées produit une silice d'environ 92% de pureté, vendue sous le nom de Clarcel, qui est utilisée comme agent de filtration dans de nombreux procédés chimiques ou pharmaceutiques
Les diatomées fossiles ont été utilisées pour stabiliser la nitroglycérine et constituer la dynamite.
Références
- Référence ITIS : Bacillariophyta (fr) ( (en))
- Référence NCBI : Bacillariophyta (en)
- Site de l'Association des Diatomistes des Langue Française -ADLaF-
- Site du Génoscope, laboratoire qui a séquencé le génome de Phaeodactylum cornutum
- Hoek, C. van den, Mann, D. G. and Jahns, H. M. Algae : An introduction to phycology. Cambridge University Press, 1995.
- Mann, D. G. The species concept in diatoms. Phycologia 1999, 38, 437-495.
- Round, F. E. ; Crawford, R. M. ; Mann D.G. The Diatoms. Biology and Morphology of the Genera, Cambridge University Press, 2007
- Yool, A. and Tyrrell, T. Role of diatoms in regulating the ocean's silicon cycle. Global Biogeochemical Cycles. 2003, 17, 1103.
- Loir, Maurice. Guide des diatomées : plus de 200 micro-algues silicieuses photographiées, Paris : Delachaux et Niestlé, 2004.
 Portail de la microbiologie
Portail de la microbiologie Portail de la botanique
Portail de la botanique
Catégories : Portail:Microbiologie/Articles liés | Bacillariophyta







Wikimedia Foundation. 2010.