- Chromatophore
-
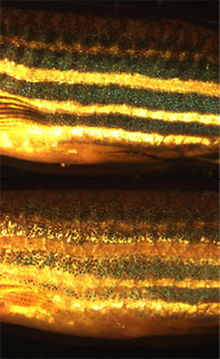 Chromatophores de Poisson zèbre à l'obscurité (au-dessus) et à la lumière (en dessous).
Chromatophores de Poisson zèbre à l'obscurité (au-dessus) et à la lumière (en dessous).
Les chromatophores sont des cellules pigmentaires ou réfléchissant la lumière présentes dans le tégument de certains animaux. Ils sont en grande majorité responsables de la couleur de la peau et des yeux des animaux à sang froid et sont créés par la crête neurale durant le développement embryonnaire. Ils sont situés à la surface du tégument de certains Amphibiens, Poissons, Reptiles, Crustacés et Céphalopodes. Les brusques modifications de couleur des téguments, visibles chez certaines espèces, sont dues aux variations de taille des chromatophores, à la migration des pigments ou à la réorientation de lamelles réfléchissantes, sous contrôle hormonal, nerveux ou mixte. Ces modifications de couleur sont souvent employées comme moyen de camouflage par homochromie, mais peuvent aussi être déclenchées par des variations d'humeur, de température, de nature de l'environnement local, ou par le stress.
Les chromatophores matures sont regroupés en sous-classes selon la couleur (ou plutôt la « teinte ») qu'ils possèdent quand ils sont éclairés par de la lumière blanche : xanthophores (jaune), érythrophores (rouge), iridophores (réfléchissant / iridescent), leucophores (blanc), mélanophores (noir ou brun) et cyanophores (bleu).
À la différence des animaux à sang froid, les Mammifères et les Oiseaux ont leurs propres sortes de cellules, les mélanocytes, qui sont l'équivalent des mélanophores. Ces derniers sont étudiés par les scientifiques pour comprendre les anomalies de pigmentation humaines, et utilisés pour la découverte de médicaments.
Le terme peut aussi se référer aux vésicules colorées associées à la membrane de certaines formes de bactéries phototrophes.
Sommaire
Classification
 Un caméléon casqué, Chamaeleo calyptratus. Les couleurs vertes et bleues sur son corps sont produites par une superposition d'un type de chromatophores qui filtrent et réfléchissent la lumière.
Un caméléon casqué, Chamaeleo calyptratus. Les couleurs vertes et bleues sur son corps sont produites par une superposition d'un type de chromatophores qui filtrent et réfléchissent la lumière.Des cellules pigmentaires d'Invertébrés sont décrites pour la première fois comme chromoforo dans un journal scientifique italien en 1819[1]. Le terme chromatophore est adopté plus tard comme le nom des cellules pigmentaires dérivées de la crête neurale des Vertébrés à sang froid et des Céphalopodes. Le mot en lui-même provient du grec khrōma (χρωμα), couleur, et phoros (φορος), porter. En revanche, le mot chromatocyte (du grec cyte ou κυτε, cellule) a été choisi pour les cellules responsables de la couleur des Oiseaux et des Mammifères. Un seul type de cellule, le mélanocyte, a été identifié chez ces animaux.
Ce n'est qu'à partir des années 1960 que la structure et la coloration des chromatophores furent suffisamment connues pour permettre l'établissement d'une sous-classification fondée sur leur apparence. Cette classification existe encore aujourd'hui même si des études plus récentes ont révélé que certains aspects biochimiques des pigments peuvent être plus utiles pour comprendre le fonctionnement de ces cellules[2].
Biochromes et schemochromes
La production de couleur peut être divisée en deux catégories, selon les chromatophores qui la réalisent : coloration par les biochromes et coloration par les schemochromes.
- Les biochromes possèdent des pigments vrais comme les caroténoïdes et les ptéridines. Ces pigments absorbent une partie du spectre visible de la lumière de façon sélective, permettant aux autres longueurs d'onde d'atteindre l'œil de l'observateur.
- Les schemochromes, aussi connus sous l'appellation « couleur structurelle », produisent une coloration en réfléchissant plusieurs longueurs d'ondes (couleurs) de la lumière "blanche" et en transmettant les autres, en provoquant des interférences des ondes lumineuses au sein de leur structure ou décomposant la lumière reçue.
Remarque:
Si tous les chromatophores contiennent des pigments ou des structures réfléchissantes (à moins de subir une mutation génétique engendrant une anomalie comme l'albinisme), toutes les cellules possédant des pigments ne sont pas des chromatophores. L'hème, par exemple, est un biochrome responsable de l'apparence rouge du sang. Il est principalement présent dans les globules rouges (érythrocytes), qui sont produits dans la moelle osseuse tout au long de la vie, et non pas formés durant le développement embryonnaire. Par conséquent, les érythrocytes ne sont pas classés comme chromatophores.Les lipophores (xantophores et érythrophores)
Les chromatophores qui contiennent d'importantes quantités de pigments jaunes de ptéridine (xanthoptéridine) sont appelés xanthophores et ceux avec un excès de caroténoïdes rouge/orange sont désignés par le terme érythrophores[2]. On a découvert que les vésicules contenant de la ptéridine et des caroténoïdes étaient souvent présentes dans la même cellule, et que la couleur globale dépendait du rapport entre les quantités de pigments jaunes et rouges[3]. De fait, la distinction entre ces deux types de chromatophores est principalement arbitraire.
La capacité à produire de la ptéridine à partir de guanosine triphosphate est une caractéristique retrouvée chez la plupart des chromatophores, mais les xanthophores semblent avoir des voies biochimiques supplémentaires qui mènent à une accumulation excessive de xanthoptéridine.
Les caroténoïdes, eux, sont généralement de source alimentaire et sont transportés dans les érythrophores. Cela a été démontré pour la première fois en élevant des grenouilles vertes communes soumises à un régime alimentaire comprenant des criquets à teneur restreinte en carotène. L'absence de carotène dans l'alimentation de la grenouille impliquait que le "filtre" à couleur rouge/orange des caroténoïdes n'était plus présent dans les érythrophores. Le résultat a été que la grenouille est apparue bleue au lieu de verte[4].Iridophores et leucophores
Les iridophores, ou guanophores, sont des cellules pigmentaires qui réfléchissent la lumière au moyen de lames cristalloïdes de schemochromes produites à partir de guanine cristallisée[5], résidu du catabolisme des acides aminés. À la lumière, elles génèrent des couleurs iridescentes à cause de la diffraction de la lumière sur l'empilement de plaques. L'orientation du schemochrome détermine la nature de la couleur observée[6]. En utilisant les biochromes comme filtres colorés, les iridophores créent un effet d'optique connu sous le nom d'effet Tyndall ou diffusion Rayleigh, produisant des couleurs brillantes bleues et vertes[7].
Un type de chromatophore proche, les leucophores, sont présents chez plusieurs espèces de poissons. Comme les iridophores, ils utilisent de la purine cristallisée pour réfléchir la lumière, produisant la couleur blanche éclatante visible chez certains poissons. Comme pour les xanthophores et les érythrophores, la différence entre iridophores et leucophores chez les poissons n'est pas toujours évidente, mais les iridophores sont généralement considérés comme générant des couleurs iridescentes ou métalliques alors que les leucophores produisent une couleur réfléchissante dans les tons blancs[7].Mélanophores
Article détaillé : Mélanocyte.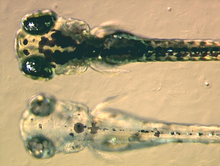 Une larve de Poisson zèbre mutant (en bas) incapable de synthétiser l'eumélanine dans ses mélanophores, en dessous d'une larve non-mutante de type sauvage.
Une larve de Poisson zèbre mutant (en bas) incapable de synthétiser l'eumélanine dans ses mélanophores, en dessous d'une larve non-mutante de type sauvage.Les mélanophores contiennent un type de mélanine, l'eumélanine, qui apparait noire ou marron foncé grâce à ses qualités d'absorption de la lumière. C'est une molécule complexe contenant des groupements dihydroxyindoles et dihydroxyindole-2-acide carboxylique, avec quelques cycles de type pyrrole[8]. Elle est contenue dans des vésicules, les mélanosomes, et distribués dans toute la cellule. L'eumélanine est produite à partir de tyrosine par une série de réactions chimiques catalysées par des enzymes. L'enzyme clé qui intervient dans la synthèse de la mélanine est la tyrosinase. Lorsque cette protéine est déficiente, aucune mélanine ne peut être générée, ce qui provoque certains types d'albinisme.
Chez plusieurs espèces d'Amphibiens, il y a d'autres pigments contenus aux côtés de l'eumélanine. Par exemple, un nouveau pigment rouge foncé a été identifié dans les mélanophores de grenouilles de la famille des Phyllomedusinaes[9]. Il a été ensuite identifié comme étant de la ptérorhodine, un dimère de ptéridine qui s'accumule autour de l'eumélanine. S'il est possible que des espèces moins étudiées aient des pigments complexes dans leurs mélanophores, il est néanmoins vrai que la majorité des mélanophores étudiés jusqu'ici contiennent uniquement de l'eumélanine.
Les humains ont seulement une sorte de cellule pigmentaire, l'équivalent chez les mammifères des mélanophores, qui produit la couleur de la peau, des cheveux et des yeux : le mélanocyte. Pour cette raison, et à cause de leur grand nombre et de leur contraste de couleurs qui font que ces cellules sont souvent aisément repérables, les mélanophores sont de loin les chromatophores les plus étudiés. Cependant, il existe des différences entre la biologie du mélanophore et celle du mélanocyte. En plus de l'eumélanine, les mélanocytes peuvent produire un pigment jaune/rouge appelé la phaéomelanine.
Cyanophores
 Une grenouille Dendrobates pumilio. Plusieurs espèces aux couleurs brillantes possèdent des chromatophores peu communs à la composition pigmentaire inconnue.
Une grenouille Dendrobates pumilio. Plusieurs espèces aux couleurs brillantes possèdent des chromatophores peu communs à la composition pigmentaire inconnue.En 1995, il a été démontré que les couleurs bleu vibrant de plusieurs sortes de poissons mandarins ne sont pas fabriquées par les schemochromes mais par un biochrome cyan de nature chimique inconnue[10]. Ce pigment, trouvé dans les vésicules cellulaires d'au moins deux espèces de poissons de la famille des Callionymidae, est très peu fréquent dans le règne animal, car toutes les autres colorations bleues étudiées jusqu'à présent sont produites par des schemochromes. Par conséquent un nouveau type de chromatophore, le cyanophore, a été proposé. Bien que peu fréquents dans la taxinomie, des cyanophores (ainsi que d'autres chromatophores inhabituels) peuvent être trouvés chez d'autres espèces de poissons et d'amphibiens. Par exemple, des chromatophores brillants aux pigments indéfinis ont été observés dans les familles des Dendrobatidae et des Centrolenidae[11].
Translocation du pigment
Beaucoup d'espèces ont la capacité de faire migrer le pigment à l'intérieur des chromatophores, provoquant un changement de couleur. Ce procédé est surtout étudié au sein des mélanophores, puisque la mélanine est le pigment le plus sombre et le plus visible. Chez la plupart des espèces possédant un derme relativement fin, les mélanophores dermiques ont tendance à être plats et à couvrir une grande surface. Cependant, chez les animaux à derme relativement épais, comme chez les reptiles adultes, les mélanophores dermiques forment souvent des structures tridimensionnelles avec d'autres chromatophores.
Ces structures (ou DCU pour Dermal Chromatophore Units) sont constituées d'une couche supérieure de xanthophores et/ou d'érythrophores, puis d'une couche d'iridophores, et enfin d'une couche de mélanophore en forme de panier avec des prolongements cytoplasmiques recouvrant les iridophores[12].Dans les deux types de disposition, les mélanophores dermiques sont importants dans le changement de couleur physiologique.
Les mélanophores dermiques plats recouvrent souvent les autres chromatophores, ce qui explique que lorsque le pigment se répand dans la cellule, la peau apparait sombre. Lorsque le pigment est aggloméré au centre de la cellule, les pigments des autres chromatophores sont exposés à la lumière et la peau prend leur teinte.
De la même manière, après l'accumulation de la mélanine dans les DCUs, la peau apparaît verte de par le filtrage par les xanthophores (jaunes) de la lumière dispersée par la couche d'iridophores. Lorsque la mélanine se disperse, la lumière n'est plus décomposée; elle est absorbée par la mélanine et la peau apparait sombre.
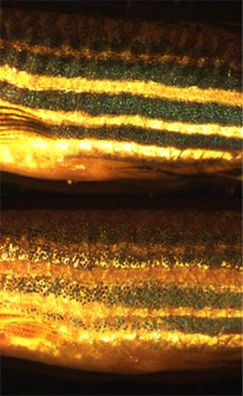
Comme les autres chromatophores biochromatiques sont également capables de réaliser cette migration de pigment, la peau des animaux avec plusieurs sortes de chromatophores peut générer de cette façon un large panel de couleurs[13],[14]. Photographie accélérée d'un mélanophore de Poisson zèbre pendant une agrégation de pigment.
Photographie accélérée d'un mélanophore de Poisson zèbre pendant une agrégation de pigment.Le contrôle et les mécanismes de la translocation de pigment ont été bien étudiés sur un certain nombre d'espèces, notamment parmi les amphibiens et les poissons téléostéens[15],[10]. Il a été démontré que le processus peut être sous contrôle hormonal, neuronal ou les deux. Les substances neurochimiques qui interviennent dans la translocation de pigment comprennent la noradrénaline, dont les récepteurs se trouvent à la surface des mélanophores[16]. Les hormones principales intervenant dans la régulation de la translocation semblent être la mélanocortine, la mélatonine et l'hormone de mélano-concentration (MCH), produites principalement par l'hypophyse, la glande pinéale et l'hypothalamus respectivement. Ces hormones peuvent aussi être fabriquées au moyen d'une régulation paracrine par des cellules de la peau. À la surface du mélanophore, les hormones activent des récepteurs couplés aux protéines G spécifiques qui, alternativement, transmettent un signal à la cellule. La mélanocortine provoque la dispersion des pigments, tandis que la mélatonine et la MCH déclenchent leur agrégation[17].
De nombreux récepteurs de mélacortine, de MCH et de mélatonine ont été identifiés sur les poissons[18] et les grenouilles[19], dont un homologue du M1CR[20], un récepteur de la mélacortine connu pour réguler la couleur de la peau et des cheveux des humains[21]. À l'intérieur de la cellule, l'adénosine monophosphate cyclique (AMPc) est un important messager secondaire de la translocation de pigment. Par un mécanisme encore imparfaitement décrypté, l'AMPc influence d'autres protéines, comme la protéine kinase A, pour activer le moteur protéique qui transporte les vésicules contenant les pigments le long des microtubules et des microfilaments[22],[23],[24].
Adaptation
 Seiche juvénile utilisant l'homochromie pour se fondre sur le fond sableux.
Seiche juvénile utilisant l'homochromie pour se fondre sur le fond sableux.- voir aussi : Mimétisme
La plupart des poissons, des reptiles et des amphibiens changent de couleur en réponse à des modifications dans leur environnement. Ce type de camouflage, ou homochromie, se manifeste généralement par un léger obscurcissement ou éclaircissement de la peau pour imiter approximativement l'environnement immédiat. Il a été démontré que ce procédé dépend de la vision (l'animal doit pouvoir voir son environnement pour s'y adapter[25]), et que la translocation de la mélanine dans les mélanophores est le principal facteur responsable du changement de couleur[26]. Des animaux comme les caméléons, les céphalopodes et les lézards anoles ont une réponse adaptative très développée capable de générer très rapidement de nombreuses couleurs différentes. Ils ont adapté leur capacité à changer de couleur en réponse aux variations de températures, d'humeur, de stress et les interactions sociales, plutôt que pour uniquement imiter leur environnement.
Développement
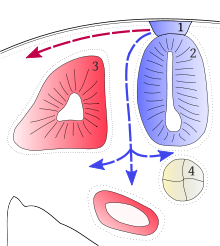 Coupe transversale d'un tronc de vertébré en développement, montrant les voies dorsolatérale (en rouge) et médioventrale (en bleu) empruntées pendant la migration des chromatoblastes.
Coupe transversale d'un tronc de vertébré en développement, montrant les voies dorsolatérale (en rouge) et médioventrale (en bleu) empruntées pendant la migration des chromatoblastes.Pendant le développement embryonnaire des Vertébrés, les chromatophores sont l'un des types de cellules produites dans la crête neurale, qui est une bande de cellules apparaissant sur les bordures du tube neural. Ces cellules ont la capacité de migrer sur de longues distances, permettant aux chromatophores de peupler de nombreux organes du corps, dont la peau, l'œil, l'oreille et le cerveau. Quittant la crête neurale par vagues successives, les chromatophores prennent soit un chemin dorsolatéral à travers le derme, entrant dans l'ectoderme par de petits espaces situés dans la lame basale, soit un chemin médioventral entre les somites et le tube neural. Les mélanophores de l'épithélium pigmenté de la rétine de l'œil constituent une exception. Ils ne sont pas issus de la crête neurale mais d'un diverticule du tube neural, qui génère la cupule optique qui, à son tour, formera la rétine.
Quand et comment les cellules multipotentes précurseurs des chromatophores, appelées chromatoblastes, se différencient pour obtenir les différents types de chromatophores est en cours de recherche. On sait que dans l'embryon de poisson zèbre, par exemple, trois jours après la fécondation, chacun des types de chromatophores retrouvés chez l'adulte (mélanophores, xanthophores et iridophores) est déjà présent. Des études employant un poisson mutant ont démontré que les facteurs de transcription, tel que kit, sox10 et microphthalmia-associated transcription factor (mitf) jouent un rôle important dans la différenciation des chromatophores[27]. Si ces protéines sont déficientes, les chromatophores peuvent être localement ou complètement absents, aboutissant à un problème de leucistisme.
Applications pratiques
En plus de la recherche fondamentale pour mieux comprendre le fonctionnement des chromatophores, les cellules sont également utilisées en recherche appliquée. Par exemple, la larve du poisson zèbre est étudiée pour comprendre comment les chromatophores s'organisent et communiquent pour former les rayures horizontales régulières observables chez l'adulte[28] . Ces larves sont considérées comme un modèle utile afin de mieux comprendre l'évolution du développement embryonnaire.
La biologie des chromatophores est également utilisée pour modéliser certaines affections humaines, comme le mélanome ou l'albinisme. Récemment, on a découvert que le gène responsable de la spécificité des mélanophores dans la variété dorée de Poissons zèbres, Slc24a5, avait un équivalent chez l'humain qui présente une forte corrélation avec la couleur de la peau[29].Les chromatophores sont aussi utilisés comme biomarqueur de la cécité chez les espèces à sang froid, car les animaux présentant certains types de déficience visuelle ne parviennent pas à réaliser l'homochromie[25]. On pense que des homologues chez l'humain des récepteurs intervenant dans la translocation de pigment des mélanophores interviendraient dans la suppression de l'appétit et dans le bronzage, en faisant des cibles attrayantes pour la recherche pharmaceutique[30]. Par conséquence, les compagnies pharmaceutiques ont développé des tests biologiques pour identifier rapidement les composés bioactifs potentiels en utilisant les mélanophores d'un crapaud africain (Xenopus laevis)[31].
D'autres scientifiques ont développé des techniques pour utiliser les mélanophores comme biosenseurs[32], et pour une détection rapide de la coqueluche (basée sur la découverte que la toxine de la coqueluche bloque l'agrégation de pigment dans les mélanophores de poissons)[33].
Des applications militaires potentielles concernant les changements de couleur des chromatophores ont été proposées, principalement comme type de camouflage actif[34].Cas des Céphalopodes
 Une seiche juvénile se fondant dans son environnement.
Une seiche juvénile se fondant dans son environnement.Les céphalopodes de la sous-classe des Coleoidea ont des 'organes' multicellulaires complexes qu'ils utilisent pour changer de couleur rapidement. C'est particulièrement notable chez les calmars, seiches et pieuvres aux couleurs vives. Chaque unité de chromatophores est composée d'une seule cellule pigmentaire et de nombreuses cellules musculaires, nerveuses, et gliales (entre autres des cellules de Schwann)[35]. À l'intérieur de la cellule du chromatophore, des granules de pigments sont enfermés dans un sac élastique, le sacculus cytoélastique. Pour changer de couleur, l'animal déforme le sacculus pour qu'il change de forme ou de taille par contraction musculaire, modifiant ainsi sa transluminescence, sa réflectivité ou son opacité. Ce mécanisme diffère de celui rencontré chez les poissons, amphibiens et reptiles, car il ne fait pas intervenir une translocation des vésicules de pigment à l'intérieur de la cellule mais un changement de forme du sacculus. Cependant l'effet est le même.
Les poulpes actionnent leurs chromatophores par vagues, obtenant une manifestation chromatique complexe et de rapides changements de couleurs. On pense que les nerfs commandant les chromatophores seraient situés dans le cerveau, dans une position similaire aux chromatophores qu'ils contrôlent. Cela signifie que l'ordre dans lequel s'effectue le changement de couleur correspond à l'ordre dans lequel se fait l'activation neuronale. Ceci expliquerait pourquoi le changement de couleur se produit par vagues, parce que les neurones sont activés l'un après l'autre[36]. Comme le caméléon, les céphalopodes utilisent le changement de couleur dans leurs interactions sociales. Ils sont également parmi les plus compétents en matière d'homochromie, ayant la capacité d'adapter leur couleur et leur texture à l'environnement local avec une remarquable précision.
Bactérie
Des chromatophores peuvent aussi être trouvés dans la membrane des bactéries phototrophes. Utilisés principalement pour la photosynthèse, ils contiennent des pigments de chlorophylle bactérienne (chlorophylle a et d) et des caroténoïdes[37]. Chez les bactéries pourpres, par exemple Rhodospirillum rubrum, les antennes collectrices sont intrinsèques aux membranes chromatophores. Cependant, chez les bactéries Chlorobiaceae, elles sont arrangées sous forme d'antennes collectrices spécialisées, les chlorosomes[38].
Voir aussi
Notes et références
- Sangiovanni G. Descrizione di un particolare sistema di organi cromoforo espansivo-dermoideo e dei fenomeni che esso produce, scoperto nei molluschi cefaloso. G. Enciclopedico Napoli. 1819; 9:1–13.
- Bagnara JT. Cytology and cytophysiology of non-melanophore pigment cells. Int Rev Cytol. 1966; 20:173–205. PMID 5337298
- Matsumoto J. Studies on fine structure and cytochemical properties of erythrophores in swordtail, Xiphophorus helleri. J Cell Biol. 1965; 27:493–504. PMID 5885426
- Bagnara JT. Comparative Anatomy and Physiology of Pigment Cells in Nonmammalian Tissues in The Pigmentary System: Physiology and Pathophysiology, Oxford University Press, 1998. ISBN 0-19-509861-7
- Taylor JD. The effects of intermedin on the ultrastructure of amphibian iridophores. Gen Comp Endocrinol. 1969; 12:405-16. PMID 5769930
- Morrison RL. A transmission electron microscopic (TEM) method for determining structural colors reflected by lizard iridophores. Pigment Cell Res. 1995; 8:28–36. PMID 7792252
- Fujii R. The regulation of motile activity in fish chromatophores. Pigment Cell Res. 2000; 13:300-19. PMID 11041206
- Ito S & Wakamatsu K. Quantitative analysis of eumelanin and pheomelanin in humans, mice, and other animals: a comparative review. Pigment Cell Res. 2003; 16:523-31. PMID 12950732
- Bagnara JT et al. Color changes, unusual melanosomes, and a new pigment from leaf frogs. Science. 1973; 182:1034–5. PMID 4748673
- Fujii R. The regulation of motile activity in fish chromatophores. Pigment Cell Res. 2000; 13:300-19. PMID 11041206
- Schwalm PA et al. Infrared reflectance in leaf-sitting neotropical frogs. Science. 1977; 196:1225–7. PMID 860137
- Bagnara JT et al. The dermal chromatophore unit. J Cell Biol. 1968; 38:67–79. PMID 5691979 Full textPDF
- Palazzo RE et al. Rearrangements of pterinosomes and cytoskeleton accompanying pigment dispersion in goldfish xanthophores. Cell Motil Cytoskeleton. 1989; 13:9–20. PMID 2543509
- Porras MG et al. Corazonin promotes tegumentary pigment migration in the crayfish Procambarus clarkii. Peptides. 2003; 24:1581–9. PMID 14706537
- Deacon SW et al. Dynactin is required for bidirectional organelle transport. J Cell Biol 2003; 160:297-301. PMID 12551954 Texte complet
- Aspengren S et al. Noradrenaline- and melatonin-mediated regulation of pigment aggregation in fish melanophores. Pigment Cell Res. 2003; 16:59–64. PMID 12519126
- Logan DW et al. Regulation of pigmentation in zebrafish melanophores. Pigment Cell Res. 2006; 19:206-13. PMID 16704454
- Logan DW et al. Sequence characterization of teleost fish melanocortin receptors. Ann N Y Acad Sci. 2003; 994:319-30. PMID 12851332
- Sugden D et al. Melatonin, melatonin receptors and melanophores: a moving story. Pigment Cell Res. 2004; 17:454-60. PMID 15357831
- Logan DW et al. The structure and evolution of the melanocortin and MCH receptors in fish and mammals. Genomics. 2003; 81:184-91. PMID 12620396
- Valverde P et al. Variants of the melanocyte-stimulating hormone receptor gene are associated with red hair and fair skin in humans. Nat. Genet. 1995; 11:328-30. PMID 7581459
- Snider J et al. Intracellular actin-based transport: how far you go depends on how often you switch. Proc Natl Acad Sci USA. 2004; 101:13204-9. PMID 15331778 Full text
- Rodionov VI et al. Functional coordination of microtubule-based and actin-based motility in melanophores. Curr Biol. 1998; 8:165-8. PMID 9443917 Texte complet
- Rodionov VI et al. Protein kinase A, which regulates intracellular transport, forms complexes with molecular motors on organelles. Curr Biol. 2002; 14:1877–81. PMID 15498498 Texte complet
- Neuhauss SC. Behavioral genetic approaches to visual system development and function in zebrafish. J Neurobiol. 2003; 54:148-60. PMID 12486702. [PDF].
- Logan DW et al. Regulation of pigmentation in zebrafish melanophores. Pigment Cell Res. 2006; 19:206-13. PMID 16704454
- Kelsh RN et al. Genetic analysis of melanophore development in zebrafish embryos.] Dev Biol. 2000; 225:277-93. PMID 10985850
- Kelsh RN. Genetics and evolution of pigment patterns in fish. Pigment Cell Res. 2004; 17:326-36. PMID 15250934
- Lamason RL et al. SLC24A5, a putative cation exchanger, affects pigmentation in zebrafish and humans. Science. 2005; 310:1782–6. PMID 16357253
- Logan DW et al. The structure and evolution of the melanocortin and MCH receptors in fish and mammals. Genomics. 2003; 81:184-91. PMID 12620396.
- Jayawickreme CK et al. Use of a cell-based, lawn format assay to rapidly screen a 442,368 bead-based peptide library. J Pharmacol Toxicol Methods. 1999; 42:189-97. PMID 11033434
- Andersson TP et al. Frog melanophores cultured on fluorescent microbeads: biomimic-based biosensing. Biosens Bioelectron. 2005; 21:111-20. PMID 15967358
- Karlsson JO et al. The melanophore aggregating response of isolated fish scales: a very rapid and sensitive diagnosis of whooping cough. FEMS Microbiol Lett. 1991; 66:169-75. PMID 1936946
- Lee I. Nanotubes for noisy signal processing: Adaptive Camouflage PhD Thesis. 2005; Université de Californie du Sud. [PDF].
- Cloney RA. & Florey E. Ultrastructure of cephalopod chromatophore organs. Z Zellforsch Mikrosk Anat. 1968; 89:250–280. PMID 5700268
- Demski LS. Chromatophore systems in teleosts and cephalopods: a levels oriented analysis of convergent systems. Brain Behav Evol. 1992; 40:141-56. PMID 1422807
- Salton MR. Bacterial membrane proteins. Microbiol Sci. 1987; 4:100-5. PMID 3153178
- Frigaard NU. & Bryant DA. Seeing green bacteria in a new light: genomics-enabled studies of the photosynthetic apparatus in green sulfur bacteria and filamentous anoxygenic phototrophic bacteria. Arch Microbiol. 2004; 182:265-75. PMID 15340781
Catégories :- Anatomie animale
- Cellule de stockage





Wikimedia Foundation. 2010.