- Phyllotaxie
-

 Pin cembro (Pinus cembra)
Pin cembro (Pinus cembra)
On appelle phyllotaxie l’ordre dans lequel sont implantés les feuilles ou les rameaux sur la tige d’une plante, ou, par extension, la disposition des éléments d’un fruit, d’une fleur, d’un bourgeon ou d’un capitule.
On appelle aussi phyllotaxie la science qui étudie ces arrangements. Il existe plusieurs types de phyllotaxies, qui dépendent :
- du nombre de feuilles par nœud
- de l'arrangement de ces feuilles le long de la tige.
Nombre de feuilles par nœud :
- Une feuille par nœud
La disposition est dite alterne. Quand ces feuilles, le long de la tige, sont sur un plan, on appelle cela une phyllotaxie alterne distique. Quand ces feuilles, le long de la tige, sont disposées tout autour de la tige, on appelle cela une phyllotaxie alterne spiralée.
- Deux feuilles par nœud
La disposition est dite opposée. Quand ces feuilles, le long de la tige, sont sur un plan, on appelle cela une phyllotaxie opposée. Quand ces feuilles, le long de la tige, sont disposées de façon perpendiculaire d'un nœud à l'autre, on appelle cela une phyllotaxie opposée décussée.
- Plus de deux feuilles par nœud ;
La disposition est dite verticillée. Elle est forcément disposée sur plusieurs plans.
La phyllotaxie : l'étude de la disposition des feuilles sur la tige
 Une feuille de plante à fleur vue en transparence
Une feuille de plante à fleur vue en transparenceLa feuille : nature et origine évolutive
La feuille est un organe photosynthétique le plus souvent composé d’une surface plane (lame ou limbe) portée par un pétiole. Elle est parcourue de vaisseaux cribro-vasculaires.
La feuille « vraie » n’est présente que chez les végétaux supérieurs dits trachéophytes, sous des formes diverses et plus ou moins dégénérées. On retrouve des organes analogues dans le reste de la lignée verte (microphylle de Physcomitrella patens par exemple) ou même chez les algues rouges par exemple, dont l’exemple le plus frappant est celui de l’algue rouge Desseleria[1].
Dans le cas de la lignée verte, les feuilles (s.l.) sont apparues de manière indépendante six fois au moins au cours de l’évolution (mousses, lycopodes, fougères, sphenopisdés, et angiospermes)[1]. Même si l’on ne sait toujours pas si les rouages génétiques mis en jeu sont similaires, il est établi qu’elles partagent au moins une trajectoire évolutive semblable : dans chaque cas, on a assisté au passage d’un système de ramification latérale à croissance indéterminée, à des structures complètement déterminées. Si les bases moléculaires de cette évolution restent à déterminer, il est par exemple notable qu’un gène, ASYMETRIC LEAVES 1 (ASI chez Arabidopsis, homologue de PHANTASTICA, PHAN, chez le muflier), semble avoir eu un rôle crucial dans l’apparition de la feuille vraie chez les dicotylédones (AS1 a une activité inhibitrice sur l’expression de KNOX, alors que les gènes de la famille KNOX sont impliqués dans le maintien d’une croissance indéterminée au niveau du méristème)[2]. Il reste à comprendre comment sont apparues les feuilles chez les mousses, lycopodes, etc. mais nous pouvons d’ores et déjà avancer que nous sommes en présence d’un exemple d’évolution parallèle particulièrement captivant.
La feuille est attachée à la tige par le biais du pétiole, au niveau de ce que l’on appelle un point d’insertion. Au point d’insertion, il y a jonction entre les faisceaux cribro-vasculaires de la feuille et ceux de la tige, selon des motifs propres à chaque espèce. À l’aisselle de la feuille on trouve généralement un bourgeon secondaire, point d’origine potentiel d’une ramification latérale.
 Chêne (Quercus lobata)
Chêne (Quercus lobata)Les feuilles : arrangements et modes phyllotaxiques
L’observation des feuilles et de la disposition sur la tige (sous forme de séries de points d’insertion) rend compte d’une géométrie qui n’était pas forcément évidente à première vue.
Une représentation usuelle et pratique consiste à identifier la tige à un cylindre ou un cône, et d’indiquer les points d’insertion à sa surface, en les numérotant selon l’ordre d’apparition des feuilles correspondantes. Un mode de coordonnées polaires permet de formaliser cette représentation. Selon les cas, on adoptera une représentation en vue de dessus (pour le cône), ou sous la forme d’un ruban déroulé (pour le cylindre). Cette transformation permet d’obtenir une représentation en deux dimensions, qui rend le dessin et son interprétation plus aisés (voir figure). Introduite par les frères Bravais (cf. historique dans la section suivante), la représentation cylindrique servira de base pour d’importants travaux réalisés en cristallographie et sur la théorie des treillis cylindriques.
On distingue trois grands modes phyllotaxiques[3], en fonction des agencements observés :
- spiralé, défini par la présence d’une seule feuille par nœud,
- opposé, où deux feuilles se font face sur un même nœud,
- verticillé, caractérisé par l’insertion de trois ou plus feuilles sur un même nœud.
Dans le cas spiralé, il a depuis longtemps été reconnu que les feuilles forment un motif spiralé qui remonte sur la tige dans l’ordre des âges décroissants. L’angle de divergence est l’angle qui existe entre deux feuilles mises en place successivement. La spirale ainsi décrite est dite générative, et l’on parle de parastiches pour désigner les motifs spiralés secondaires (plus verticaux) qui apparaissent par le rapprochement spatial de feuilles non directement successives. Le nombre des parastiches (dextres et senestres) est utilisé pour caractériser les différents motifs phyllotaxiques. Une observation remarquable, que l’on doit à Alexander Braun en 1831, établit le lien qui existe entre le nombre de parastiches et la suite de Fibonacci. En effet, dans l’extrême majorité des cas, les nombres de parastiches que l’on retrouve chez une plante, dans un sens et dans l’autre, sont deux termes consécutifs de la suite de Fibonacci (voir figure 2). Nous laisserons à d’autres les spéculations ésotériques sur ce thème, très souvent inspirées par le fait que la limite infinie de la suite de Fibonacci (dans le cas général (1;1)) est j, le nombre d’or… Une remarque à ce sujet sera faite dans la partie Considérations évolutives ci-dessous.
Dans le cas opposé ou verticillé, on assiste entre chaque nœud à une rotation des axes des feuilles, qui vaut la moitié de l’angle entre deux feuilles insérées sur le même axe (par exemple : 90 ° dans le cas d’un système opposé, 60 ° dans le cas de verticilles de trois feuilles).
Le plastochrone est la période qui sépare l’initiation de deux feuilles successives.
Les théories de la phyllotaxie
Approche historique et modèles phénoménologiques
Aperçu historique de l’étude de la phyllotaxie
L’histoire des observations et de la compréhension de la phyllotaxie est indicative sur l’évolution des paradigmes culturels constituant le contexte de pensée des différentes époques considérées (voir pour revue[4]). Si les premières observations des motifs formés par les organes semblent remonter aux Égyptiens de l’Antiquité, trois grandes phases apparaissent dans l’étude « moderne », la caractérisation et la compréhension des types phyllotaxiques.
Les XVe et XVIe siècles ont été marqués par la découverte des motifs phyllotaxiques et leurs liens à la suite de Fibonacci.
Du XVIe au début du XXe siècle, une caractérisation exhaustive des modes phyllotaxiques a été entreprise, et les premiers modèles interprétatifs ont été proposés, s’appuyant en particulier sur des modélisations géométriques et la notion d’espace disponible sur le méristème.
La fin du XXe siècle a été la plus active, en termes d’avancées de la compréhension des mécanismes, tant sur le plan mathématique que biologique avec notamment les découvertes de certains gènes organisateurs du méristème liés à l’établissement des feuilles.
Des premières observations aux premières surprises
Si l’on trouve parmi les pionniers de l’étude de la phyllotaxie des personnages célèbres comme Aristote, Pline l’Ancien, Léonard de Vinci ou Johannes Kepler, qui ont contribué aux premières réflexions sur le sujet, en marge de leurs œuvres majeures respectives, les premières observations systématiques ont débuté avec Charles Bonnet (1754) qui introduit les notions de spirale génératrice et de parastiches, et commence à caractériser les différents types phyllotaxiques en en décrivant quatre principaux types (décrits précédemment par Sauvages) : opposé, verticillé de trois feuilles ou plus, alterne, et sans arrangement constant et en en mentionnant un nouveau, le mode spiralé. Cette classification est utilisée par Carl von Linné en 1751. Ces observations sont reprises moins d’un siècle plus tard par Karl Friedrich Schimper (1830) qui introduit le concept d’angle de divergence et achève de décrire les parastiches.
À partir de ce point, la première moitié du XIXe siècle se montre riche en propositions et avancées : Alexander Braun, en 1831, est le premier a établir le lien entre le nombre de parastiches et la suite de Fibonacci. Dans un article commun publié en 1837, les frères Auguste et Louis Bravais proposent la première représentation géométrique formelle de la disposition des feuilles sur une tige — au moyen d’un réseau de points (treillis) sur un cylindre — et présentent leurs travaux visant à caractériser les propriétés de cette classe d’objets mathématiques, notamment sur le lien entre angle de divergence et nombre de génératrices et parastiches.
 Tournesol (Helianthus annuus)
Tournesol (Helianthus annuus) Tournesol (Helianthus annuus)
Tournesol (Helianthus annuus)Vers une explication du phénomène des spirales phyllotaxiques : construction d’un cadre théorique
S’enclenche un processus de théorisation du problème de la phyllotaxie : l’enjeu est de proposer le mécanisme qui explique la disposition des organes sur la tige. En 1868, Wilhelm Hofmeister (1824-1877), dans son Handbuch der Physiologischen Botanik, propose la première règle simple qui rendrait compte a priori de cette disposition :
« Le primordium apparaît périodiquement dans le plus grand espace disponible. »
En 1875, Julius von Wiesner (1838-1916) reprend les arguments téléologiques développés précédemment par De Vinci et Charles Bonnet (1720-1793), et les transpose sous le regard de la théorie de l’évolution de Darwin[5], en émettant l’hypothèse que la phyllotaxie optimise l’interception de la lumière et que la disposition spiralée est celle qui autorise le minimum d’ombrage d’une feuille vis-à-vis des autres. En 1878, Simon Schwendener propose un modèle où la disposition des feuilles serait orientée par les pressions de contact entre les primordium. Malgré une erreur de raisonnement, il prédit avec beaucoup de précision de nombreux profils phyllotaxiques et relève le premier les questions de transitions entre les différents profils : (1,2) à (3,2) à (3,5)… au cours du développement de la plante. Airy présente en 1873 une variation de la règle de Hofmeister, considérant que les arrangements phyllotaxiques sont une optimisation de la réduction de place occupée et propose un principe d’« économie d’espace ». Le mode spiralé serait ainsi le mode de paquetage qui occuperait le plus petit volume possible.
Ces trois contributions ont un impact important au sein de la communauté et vont amener à un changement radical dans la manière de percevoir la question de la phyllotaxie : c’est à l’apex et au méristème, là où les primordia sont générés et organisés, et non plus à la tige édifiée (reflet figé d’une dynamique qui lui est antérieure), qu’il faut s’intéresser si l’on veut comprendre les origines des dispositions observées.
Après une période de troubles et d’indécision, suite à la controverse lancée par le mathématicien Julius Sachs en 1882 qui arguait que l’idée des spirales phyllotaxiques se résume à un orgueilleux jeu mathématique, purement subjectif et non informatif sur la nature même de l’objet biologique considéré[6], une représentation polaire des primordia sur le méristème est proposée par Church en 1904, persuadé que le mystère de la phyllotaxie trouvera sa solution par l’étude de l’extrémité croissante de la tige. Délaissant le concept de spirale génératrice, il émet l’hypothèse que les parastiches représentent des lignes de forces qui contraignent la surface du méristème par un jeu de tensions. Des ondulations de différentes longueurs d’ondes seraient alors générées qui organiseraient les motifs phyllotaxiques[7].
En 1907, Van Iterson étudie de manière systématique les différents agencements possibles de disques sur un cylindre (en faisant varier l’angle de divergence et le diamètre des disques) et caractérise les différents motifs obtenus par le nombre de parastiches dextres et senestres. (figure 3) Il construit ainsi un diagramme de phase qui décrit l’univers des hélices en fonction des deux paramètres choisis. Ces travaux d’une portée singulière, n’ont pas été reconnus tout de suite et il a fallu attendre près de 70 ans avant de les redécouvrir[8].
De manière contemporaine à Van Iterson, Schoute reprend les travaux de Schwendener de manière critique et constructive : ainsi, s’il pense que c’est bien de cette manière qu’il faut considérer la question de la phyllotaxie (en la replaçant dans un contexte de croissance et en s’intéressant aux transitions entre différents modes), le mécanisme directement impliqué dans l’initiation et l’organisation des primordia est laissé en suspens. Schoute envisage alors l’existence d’un inhibiteur produit par chaque primordium qui empêche le développement d’un nouveau primordium à proximité. Le profil de concentration de l’inhibiteur au sein du méristème va alors déterminer le site d’apparition des nouveaux primordia. Schoute pressent alors ce qui sera développé par Alan Turing en 1952, avec sa théorie des morphogènes, dont l’un des exemples donné pour illustrer son travail est celui d’un anneau de cellules, dans lequel il obtient des profils d’onde stationnaire qui pourrait rappeler la disposition des tentacules de l'hydre aquatique ou de feuilles en verticilles.
L’expérimentation et la phyllotaxie
1932 est également une année intéressante pour la compréhension de la phyllotaxie, avec la parution des travaux des époux Snow. Utilisant des techniques de micro-ablation, ils ont été capables de montrer que le site d’initiation d’une nouvelle feuille est influencé par la position des primordia les plus récemment mis en place. Wardlaw poursuit leur travaux en s’intéressant à la fougère Dryopteris. Mais les conclusions divergent : là où Wardlaw imagine un mécanisme reposant sur la production d’un inhibiteur par les primordia existant qui empêche l’initiation d’un nouveau primordium à proximité, les Snow proposent que les primordia ne se développent que dans le premier espace suffisant et suffisamment éloigné du sommet du méristème[9]. À leur suite, des travaux de manipulation des motifs phyllotaxiques ont été menés que cela soit par ablation ou avec l’utilisation de différents produits chimiques, notamment Meicenheimer (1981 et 1982) avec son travail sur epilobium.
Modèles contemporains
Un certain nombre de travaux ont marqué la deuxième moitié du XXe siècle et ont permis des percées significatives dans la compréhension des mécanismes de la phyllotaxie. Nous nous contenterons ici de les rappeler, pour l’unité de cet aperçu historique, étant entendu qu’une présentation et une analyse de l’état actuel des connaissances sur la phyllotaxie sera proposée dans la section suivante…
On distingue trois grands champs d’investigation :
- Au niveau phénoménologique, les travaux de Adler, Jean et Barabe d’une part, et de Douady et Couder d’autre part, ont permis une compréhension satisfaisante de l’organisation des primordia au niveau du méristème, en s’appuyant sur des considérations physiques. L’idée est grossièrement celle d’une minimisation d’énergie d’interaction entre les différents primordia. Douady et Couder ont notamment reproduit avec une stupéfiante précision l’ensemble des motifs phyllotaxiques, tant par modélisation numérique qu’analogique (grâce à un montage expérimental utilisant des gouttes de ferro-fluides dans un champ magnétique).
- Sous l’impulsion de Green, des progrès notables ont été faits en ce qui concerne la compréhension du comportement physique du méristème et de sa surface (plissements, torsions, etc.). Coen, Fleming et Dumais mènent des recherches sur des thématiques proches, par l’analyse des dynamiques de divisions cellulaires et d’initiation d’organes, au niveau de l’apex.
- Les avancées en biologie moléculaire ont permis la découverte et la caractérisation à un rythme toujours croissant de nombreux gènes impliqués dans le fonctionnement du méristème. Nous pouvons à ce titre mentionner les travaux de Hake sur les gènes à Homéoboîte de la famille KNOX qui ont été précurseurs dans ce domaine. La démocratisation de techniques à haut-débit devrait également permettre d’accéder à la quantité de données nécessaire à la dissection des réseaux de régulation intervenant dans la morphogenèse méristématique. Grâce par exemple aux travaux de Reinhardt, Kuhlemeier, et Traas, un lien avec les hormones végétales a pu être établi. L’auxine occupe notamment le devant de la scène, avec la caractérisation récente de mutants pour des transporteurs d’auxine présentant une absence de feuille, mais on entrevoit d’ores et déjà des rôles pour d’autres effecteurs hormonaux, par exemple les cytokinines, depuis la caractérisation du mutant Abnormal Phyllotaxis par Jackson (AbPhy codant un élément de réponse à la cytokinine). En parallèle, des études sur l’évolution des mécanismes développementaux sont menées, notamment sous l’égide de Cronk, Hasebe, et Paquet (ceci est aujourd’hui rendu possible par un accès de plus en plus étendu et fiable aux données de la génomique et à l’élargissement du spectre des organismes étudiées – génomes séquencés, banques d’EST, études d’organismes satellites…).
Théories actuelles
Vers une confirmation des hypothèses de Hofmeister et de Snow & Snow
Douady et Couder, physiciens à l’ENS, publient en 1996 une série de trois articles sur la phyllotaxie[10]. Ils abordent le problème à partir de deux modèles distincts, et testent de manière empirique le postulat formulé initialement par Hofmeister (1868), élargi ensuite par les travaux des époux Snow (1952) (cf. historique en section précédente).
L’approche est résolument expérimentale, et le paradigme est celui posé par Hofmeister : l’encombrement spatial et la dynamique de croissance du méristème et de mise en place des primordia sont une condition suffisante pour l’organisation des primordia selon des spirales régulières. Le dispositif expérimental du premier modèle est novateur[11],[10] et séduit par la simplicité de son principe : des gouttelettes de ferro-fluide (de même dimension) sont disposées à intervalles réguliers sur un plateau circulaire horizontal, au sein d’un champ magnétique orienté verticalement. Ces gouttelettes se comportent alors comme des petits aimants, se repoussant les uns les autres, avec une intensité qui varie uniquement en fonction de la distance qui les sépare les unes des autres. Selon la période qui sépare chaque goutte, il est possible de décrire l’ensemble des motifs phyllotaxiques spiralés de la branche « de Fibonacci » au cours d’un régime stationnaire. En faisant varier cette période au cours d’une même expérience, il est possible d’observer des régimes de transition entre deux modes successifs de spirales. C’est l’illustration, par exemple, des transitions d’ordres observées au sein des inflorescences des Astéracées, comme le tournesol (l’ordre des spirales proches du centres (21; 34) et (34; 55) est inférieur à celui des spirales externes (55; 89)).
Le deuxième modèle expérimental est une simulation numérique qui s’appuie sur un jeu d’hypothèses restreint en reprenant le même principe directeur. Les résultats obtenus par simulations numériques confirment les données expérimentales.
La période entre chaque goutte est directement liée au plastochrone[12], de même que l’est la vitesse de migration des gouttes dans le champ magnétique (cf. définition du plastochrone ci-dessus). Les données accumulées par Douady et Couder viennent donc renforcer les travaux de leur prédécesseurs en établissant une relation directe entre l’ordre des spirales et le plastochrone de Richardson. En jouant avec les conditions initiales, la variation continue du plastochrone permet de parcourir l’ensemble de l’arbre de Van Iterson.
Les simulations, en se fondant sur un jeu d’hypothèses correspondant au postulat de Snow and Snow, permettent de retrouver les motifs verticillés, et d’observer les transitions verticilles→spirales / spirales→verticilles, comme l’avait décrit Schoute dans les années 1930, à partir d’observation.
 Pin sylvestre (Pinus sylvestris)
Pin sylvestre (Pinus sylvestris) Pin sylvestre (Pinus sylvestris)
Pin sylvestre (Pinus sylvestris)Biophysique du méristème
Green propose dès 1992[13] une théorie qui envisage l’organisation des primordia comme un problème de biophysique du méristème. Il considère en effet que c’est la réaction de différentes assises cellulaires (en particulier la couche L1) à des contraintes de tensions et de turgescences, qui entraîne la formation de motifs spiralés ou verticillés. En distinguant deux niveaux[14],[15], topographique (i.e. à l’échelle du méristème) et cellulaire, il propose une théorie dans laquelle les repliements des tissus selon une longueur d’onde particulière, qui est fonction des caractéristiques propres du matériel contraint, détermine la régularité du processus. Les équations différentielles de la physique des matériaux (flambage) permettent d’appréhender ces mécanismes. Ces mouvements globaux sont eux-mêmes dépendants de dynamiques locales : les divisions cellulaires orientées et les renforcements des parois en cellulose induits par les microtubules contrôlent les arrangement cellulaires et l’orientation de la croissance et déterminent ainsi les propriétés biophysiques des tissus.
Ces propriétés latentes sont influencées par un jeu de conditions limites qui spécifie la manière dont s’exprime le plissement : spiralé ou verticillé… Une importance particulière est donnée à la géométrie du méristème, en particulier aux variations des rayons de courbure locaux, puisque il existe une boucle de rétroaction dans laquelle la courbure influence l’orientation des microtubules et donc des plans de divisions et la direction de croissance, direction de croissance qui détermine directement l’évolution du rayon de courbure. Green, par une approche originale, confirme le consensus qui a émergé au cours du XXe siècle : il existe un rôle prépondérant de la géométrie du méristème et des initia foliaires sur la géométrie du positionnement des feuilles le long de la tige. De fait, certaines formes de méristème seraient donc incompatibles avec certains profils phyllotaxiques. Ces considérations théoriques indiquent que l’établissement rapide de configuration énergétiquement stable se révélerait être un moyen robuste de spécification des motifs phyllotaxiques au niveau du méristème[16],[17].
Ses propos sont argumentés sur la base d’observation de réponses organogénétiques à de simples interventions chirurgicales au niveau de différents méristèmes. Par exemple, le pincement temporaire d’un méristème grâce à une pince en verre (modification artificielle des contraintes et des courbures au sein du méristème) induit l’apparition d’une feuille ectopique orientée perpendiculairement à l’axe des contraintes. Lorsque la pince est retirée, de nouvelles feuilles se forment, organisée normalement (verticille de 5 primordia)[18]. Les travaux de Fleming sur la protéine expansine (voir ci-après), qui n’est censée affecter que les propriétés physiques de la paroi cellulaire (relâchement), ont montré que l’application d’expansine sur le côté d’un méristème suffisait à induire la formation d’un primordium ectopique. Ces résultats suggèrent clairement un rôle des forces biophysique dans la régulation de l’organogenèse de la plante.
Dans ce sens, les travaux initiés par Coen, Fleming ou Dumais, sont complémentaires de ceux de Green et s’inscrivent dans ce que Coen appelle « la génétique de la géométrie ».
« La génétique de la géométrie »
Coen et son équipe du John Innes Centre à Norwich est célèbre pour ses travaux sur le développement des fleurs, et sur les mécanismes de régulation de l’architecture florale (rupture de symétrie, etc.), notamment sur la plante modèle Antirrhinum majus, le muflier. S’intéressant dans un premier temps à l’identification des déterminants génétiques de ces mécanismes, Coen a par la suite su apporter un cadre conceptuel au sujet en tentant de formaliser les liens possibles et/ou nécessaires entre « morphogènes » (produits de gènes), dynamiques cellulaires et établissement de l’identité de cellules/tissus[19]. Pour suivre la croissance d’un tissu, quatre paramètres doivent être connus : le taux de croissance ainsi que l’anisotropie, la direction et la rotation qui caractérise cette croissance. Il passe ainsi en revue différentes approches expérimentales que l’on peut mettre en place tant pour comprendre les dynamiques de croissance que pour tester un certain nombre d’hypothèses (par simulations notamment). En s’appuyant sur les concepts de morphogènes annoncés par Turing (équations de réaction-diffusion), et qui ont depuis trouvé illustration dans le vivant par de nombreux exemples, il poursuit la théorie en l’appliquant à des modèles en croissance qui intègrent l’effet de croissances anisotropes en un champ de contraintes et de déformations biophysiques.
Ces travaux font réellement le lien entre l’approche purement génétique du problème et les travaux de Green. Les premières confirmations expérimentales sont fournies par Fleming et Dumais.
Des outils pour suivre les dynamiques cellulaires
Dumais, en collaboration avec Kwiatkowska[20],[21], a développé une méthode d’observation non destructive du méristème qui permet de suivre la croissance et les dynamiques cellulaires dont il est le siège. Pourvu des algorithmes nécessaires, le procédé permet l’acquisition des paramètres clés de croissance de chaque cellule, et permet d’établir le lien avec des niveaux d’expression de gènes (préalablement marqués par une sonde fluorescente). Cette avancée technique est un outil précieux qui autorisera une connaissance fine des dynamiques cellulaires, et facilitera sans aucun doute la caractérisation de phénotypes mutants pour des gènes du développement et la compréhension de certains mécanismes.
Dynamiques cellulaires et morphogenèse au niveau du méristème
Fleming s’intéresse particulièrement aux dynamiques cellulaires et à leur lien avec les processus développementaux : il montre avec Wyrzykowska que les divisions cellulaires peuvent être altérées, au moyen de diverses substances, mais avec des effets différents en termes de morphogenèse. Dans un premier article[22], il montre que la protéine expansine déclenche la formation d’un renflement, lorsqu’elle est appliquée sur le méristème (par micro-injection), renflement qui peut dans certains cas se développer en des organes qui ressemblent à des feuilles (trichomes + marqueurs moléculaires feuille spécifique). L’expansine est une protéine connue pour accroître l’extensibilité des parois des cellules végétales.
Une expérience intéressante est également présentée dans laquelle une micro-injection d’expansine judicieusement placée permet d’inverser le sens de la spirale phyllotaxique. Dans un autre cadre, des données d’expression indiquent également que l’expression du gène de l’expansine co-localise avec les zones d’initiation des primordia. Fleming et Wyrzykowska suggèrent que l’expansine induit des modifications dans les parois des cellules qui conduisent à une modification des contraintes physiques dans le méristème. Cela provoque un renflement, qui acquiert alors une identité de primordium. Ces données biomoléculaires sont en conformité avec les théories biophysiques évoquées précédemment. Deux autres articles de Fleming poursuivent dans cette direction, en s’intéressant cette fois ci aux relations entre les dynamiques cellulaires et la morphogenèse elle-même[23],[24]. En modifiant l’expression de certains gènes du cycle cellulaire, il provoque une altération des motifs de division (prolifération cellulaire notamment). Ces altérations n’ont aucun effet morphogénétique particulier au niveau du méristème alors qu’au niveau du primordium foliaire, l’induction de ces gènes conduit à une profonde modification de la croissance du limbe et à terme, à une modification importante de la forme de la feuille. Ces résultats l’amènent à penser que l’effet des dynamiques cellulaires sur la morphogenèse est dépendant du contexte. Cependant, une étude avec des micro-injections de phragmoplastine utilisée pour perturber les cycles cellulaires et les divisions montre que, dans ce cas également, la modification de l’orientation des plans de divisions cellulaires dans le méristème ne suffit pas à déclencher l’organogenèse (d’un primordium par exemple), mais que cette altération est accompagnée d’un changement dans les profils d’expression de gènes impliqués dans l’apparition d’une nouvelle feuille (PHANTASTICA-like, STM-like). Ces travaux permettent d’écarter l’hypothèse qui établissait un lien causal entre division cellulaire et morphogenèse[25]. Au contraire, il semble se dégager des mécanismes alternatifs cohérents avec ces résultats, avec en premier plan le rôle des propriétés physiques des parois des cellules du méristème. Il est également important de noter que la modification des profils d’expression de certains gènes semble indiquer l’existence d’un mécanisme qui permettrait à un groupe de cellules de percevoir des modifications dans les divisions cellulaires de ce groupe, de les interpréter comme marqueurs d’une initiation d’organogenèse, et de s’engager dans cette voie de différenciation (profils d’expression altérés). Ceci sans qu’apparemment ce mécanisme se suffise à lui-même (aucune organogenèse observée).
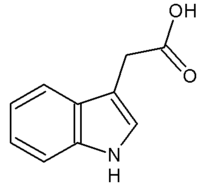 Une auxine : l'acide indole 3-acétique
Une auxine : l'acide indole 3-acétiqueL’auxine et régionalisation du méristème
Des avancées notables ont été effectuées au cours de ces dix dernières années, sur le rôle de l’auxine dans l’organogénèse foliaire et en particulier dans l’organisation du méristème. C’est en particulier à l’équipe de Kuhlemeier que l’on doit les travaux les plus exceptionnels[24],[26].
Le rôle de l’auxine comme facteur de croissance (Darwin père et fils, 1880) et son mode d’action potentiellement anisotrope expliqué par répartition asymétrique (Boysen-Jensen, 1913) sont connus depuis le début du XXe siècle, même si la molécule n’a été véritablement isolée et purifiée qu’un peu plus tardivement (Went, 1926). Cinquante ans plus tard, les travaux de Rubery et Sheldrake (1974) mettent en évidence le lien entre la distribution anisotrope d’auxine dans les tissus et la répartition asymétrique de transporteurs d’auxine. Les dernières avancées en la matière concernent la caractérisation d’une famille de protéines PINFORMED (PIN) dont l’expression est intimement liée au transport polarisé de l’auxine.
Deux séries de données sur les protéines PIN concernent particulièrement le champ de la phyllotaxie. Ainsi, il a été montré que le transporteur d’efflux d’auxine, PIN1, est exprimé selon une disposition particulière, avec, au sein des assises cellulaire supérieures (tunica), accumulation sur les parois orientées vers le centre des primordia émergents. Cette localisation suggère une orientation du flux d’auxine et l’accumulation d’auxine au centre des primordia[16]. D’autre part, l’application localisée d’auxine à la surface du méristème est suffisante pour déclencher la formation d’un nouveau primordium. Ces observations concordent avec les modèles où l’on considère les primordia comme des puits qui drainent l’auxine. Il s’établit alors un profil de concentration d’auxine autour du méristème. En tout point du méristème, la concentration en auxine dépendra de plusieurs paramètres, en particulier du motif phyllotaxique déjà établi (primordia déjà initiés), et de la distance relative aux puits les plus puissants. Une zone, la plus éloignée des derniers primordia mis en place aura, de facto, un maximum local d’auxine, qui déclenchera, si le niveau atteint est suffisant, la formation d’un nouveau primordium.
Un certain nombre de résultats expérimentaux semble aller dans le sens de ce cadre de pensée. Par exemple, la formation d’organes latéraux est inhibée si l’on disrupte l’expression des gènes PIN, ou si l’on retire la couche L1 (par ablation chirurgicale). Mais elle est rétablie dans tous les cas par la micro-application d’auxine au niveau de l’apex.
Il convient de faire deux remarques à propos de ces résultats :
- l’auxine est connue pour son action relaxante au niveau des parois végétales. Elle a ainsi un rôle semblable, sur ce point, à l’expansine utilisée par Fleming (cf. ci-dessus) ;
- avec ce modèle explicatif, on perçoit assez aisément un parallèle entre les profils d’auxine autour du méristème et les considérations introduites par les (bio)physiciens, de profils d’énergie ou d’encombrement spatial. L’auxine pourrait très bien matérialiser ou au moins marquer les champs de tensions dans le méristème.
Évolution des modèles de compréhension
Cependant, la coïncidence du profil de répartition d’auxine, des profils d’expression de gènes de différenciation, et des contraintes biophysiques s’appliquant à la surface du méristème nécessite une réflexion sur les liens de causalité qui unissent ces trois facettes d’un même phénomène. L’existence d’une boucle régulation positive entre concentration et direction du flux d’auxine et disposition des transporteurs qui fait que ce mécanisme s’auto-entretient, et l’influence de l’auxine et de l’expansine dans la modification des propriétés physiques des parois sont des premiers pas prometteurs vers la compréhension de ces interactions mais la question des transducteurs et de l’intégration de ces deux fronts (biophysique/physiologique) au sein du méristème reste encore ouverte à la discussion. Peut-être pouvons-nous supposer que cette intégration est réalisée par l’activité cellulaire des zones concernées qui, si elle n’est pas l’élément initiateur de l’organogenèse, en est un maillon clé puisque c’est elle qui va déterminer l’organisation des cellules qui serviront de canaux à l’auxine, et qui constitueront le tissu même soumis à l’action des contraintes biophysique.
Bilan et questions ouvertes
Les découvertes récentes présentées ci-dessus rendent très actuelles les théories élaborées aux XIXe et XXe siècles ; et les concepts qui avait été proposés alors méritent d’être reconsidérés pleinement : ils prennent corps, aujourd’hui, dans l’auxine organisatrice de Reinhardt et Kuhlemeier ou dans les équations différentielles de Green. Les intuitions formulées alors, les considérations stériques des frères Bravais et d’Hofmeister, les pressions de contact et les ondes de contraintes de Church, ou les inhibiteurs de Wardlaw ou Schoute, ont en fait véritablement représenté les projets de recherches conduits aujourd’hui.
Il semble, qu’enfin, la levée de la controverse est proche et qu’un consensus s’établit autour d’une synthèse « mécano-physiologique » qui concilie les théories biophysiques et les avancées dans la compréhension de la physiologie des plantes et de leur développement. Les prochaines avancées sont à attendre dans la compréhension qualitative et quantitative des réseaux de gènes impliqués dans le contrôle et la régulation du méristème. C’est également au niveau des transducteurs et des zones intégratrices qu’il faut poursuivre : d’importantes découvertes devrait en surgir. Les questions évolutives, abordées par l’évo-dévo, qui touchent à la mise en place de toute cette machinerie sont d’ailleurs tout autant passionnantes et ne manqueront pas de se révéler riches en surprises.
Considérations fonctionnelles et évolutives
La découverte d’Alexander Braun (lien entre phyllotaxie et suite de Fibonacci) a, au fil des siècles, suscité moult discussions. Et la transition entre explication téléologique et évolutive n’a pas forcément limité le nombre d’incongruités énoncées à ce sujet.
Ainsi l’idée selon laquelle la disposition des feuilles en spirale est optimale[27], et « qu’on doit y voir » la trace du lent modelage par l’évolution, bien que séduisante, perd un peu de sa force face à quelques considérations simples : ainsi, l’étude de la répartition des modes phyllotaxiques dans le règne végétal montre que la moitié au moins des genres (chez les angiospermes) a une phyllotaxie verticillée ou opposée. Le groupe qui a la plus grande répartition géographique dans le monde (les Poacées) est doté d’une phyllotaxie alterne distique extrêmement conservée, les seules exceptions connues étant les quelques mutants pour la phyllotaxie comme Abphyl chez le maïs ou Sho chez le riz par exemple (respectivement opposée et spiralée altérée). L’avantage relatif qui découlerait d’une phyllotaxie spiralée semble donc minime ou négligeable à la vue de ces statistiques.
L’observation d’un arbre en plein soleil (sans considération pour sa phyllotaxie propre) montre que seule une fraction (non négligeable cependant) des feuilles reçoit la lumière directe du soleil, somme faite sur l’ensemble de la journée. Les simulations montrent que c’est plutôt l’orientation des branches (conduite par le phototropisme) qui semble être le facteur prépondérant. Dans le même esprit, les modélisations réalisées par Niklas (1988), dans lesquelles un grand nombre de paramètres du développement des plantes étaient pris en compte dans leur relation avec l’interception de la lumière, il ressortait en particulier que le besoin en lumière n’amenait pas nécessairement la plante à adopter l’angle de Fibonacci, car l’illumination optimale était obtenue par la production de longue et fines feuilles. De surcroît, l’existence des pétioles autorise une certaine plasticité dans l’orientation de la feuille pour s’adapter aux conditions de son microenvironnement, comme l’indique ces deux clichés de rhododendron pris sur les bords d’un loch ensoleillé (voir figure 4).
Il est cependant certain que la phyllotaxie est sujette à sélection, tout du moins indirectement. Ainsi, par toutes les contraintes que nous avons pu énumérer en ce qui concerne le méristème, qu’elles soient biophysiques ou, plus pragmatiquement, génétiques et physiologiques, le méristème est un tissu en équilibre dynamique. Certains mutants (comme terminal ear1) montrent une telle dérégulation au niveau du fonctionnement du méristème et un phénotype tellement anarchique qu’ils sont extrêmement peu viables. La grande rareté de mutants clairs pour la phyllotaxie (i.e. avec une altération de la phyllotaxie qui ne soit pas simplement un effet secondaire d’une altération de processus développementaux plus larges, par exemple, abphyl semble être un bon candidat)) est elle aussi révélatrice. Il semblerait donc qu’une altération au niveau des gènes impliqués dans la régulation du méristème soit dans la plupart des cas létale, et c’est en particulier la découverte de mutants comme ceux de la série pinhead, ou encore wuschel ou clavata3, qui ont permis de franchir les premières étapes dans la compréhension de cette machinerie finement régulée. Des différences infimes dans le fonctionnement du méristème (notamment en ce qui concerne sa géométrie propre ou sa taille) permettraient d’expliquer des différences de phyllotaxie, les pressions évolutives ne s’appliquant pas directement sur la phyllotaxie elle-même mais plutôt sur des paramètres propres du méristème qui assurent sa viabilité[10].
Les processus évolutifs qui ont assuré l’apparition d’une entité embryogénique aussi structurée que le méristème restent aujourd’hui encore obscurs, même en ce qui concerne par exemple l’éventuelle filiation entre cellule apicale de bryophyte et méristème des ptéridophytes. C’est sans doute l’identification de processus, possiblement distincts et à différents stades évolutifs de la lignée verte, mais ayant recruté des gènes homologues que l’on pourra appréhender les trajectoires évolutives suivies : filiation directe ou convergence évolutive.
Notes
- Cronk QCB (2001) Plant evolution and development in a post-genomic context. Nat. Rev. Gev. 2 (8): 607-619.
- Byrne ME, Barley R, Curtis M, Arroyo JM, Dunham M, Hudson A, Martienssen RA (2000) Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis, Nature, 408 (6815): 967-971 DEC 21 2000.
- Steeves TA and Sussex IM Patterns in plant development. Cambridge University Press, Cambridge, 1989.
- Adler I, Barabé D et Jean RV (1997) A history of the study of phyllotaxis. Ann. Bot. 80: 231-244.
- La première édition de On the Origin of Species est publiée en 1860.
- En 1950, Plantefol reprendra dans une certaine mesure cette approche, en se restreignant à l’étude d’une seule famille de spirales (les spirales foliaires) et en s’affranchissant de toute considération mathématique.
- Cette hypothèse est aujourd’hui soutenue sous une forme remise au goût du jour par le laboratoire de Paul Green (décédé en 1998) et Paul Dumais et bénéficie des dernières avancées en termes de modélisation et de mécaniques des milieux hétérogènes que sont les tissus biologiques.
- Par Erickson en 1977, qui s’en sert pour la caractérisation expérimentale de motif phyllotaxiques et la détermination des paramètres associés (divergence et plastochrone).
- Sans foncièrement s’opposer, ces deux théories ne s’intéressent pas au même niveau de compréhension. Les Snow y voie un argument en faveur de la théorie de Wilhelm Hofmeister (1824-1877) et l’élargissent, en y rajoutant une notion de temporalité (signifié par le déplacement relatif du primordium sur le méristème), tandis que Wardlaw se place délibérément dans le cadre de la théorie des champs inhibiteurs.
- Douady S and Couder Y (1996) Phyllotaxis as a Dynamical Self Organizing Process (Part I, II, III), J. Theor. Biol. 139, 178-312.
- « Our aim was to see whether these principles were able, by themselves, to produce the spiral organizations hitherto observed in plants. We thus tried to find a non biological system reproducing the dynamical properties given by Hofmeisters’ hypotheses. » Douady et Couder
- Dans la démonstration de Douady et Couder, le paramètre G est utilisé, avec G=ln(a).
- Selker JML, Stichic GL and Green PB (1992) Biophysical mechanisms for morphogenetic progressions at the shoot apex. Dev. Biol. 153, 29-43.
- Green PB (1999) Expression of pattern in plants: combining molecular and calculus-based biophysical paradigms. Am. J. Bot. 86: 1059-1076
- Green PB, Steele CS and Rennich SC (1996) Phyllotactic patterns: a biophysical mechanism for their origin. Annals of Botany 77: 515–527.
- Fleming AJ (2005) Formation of primordia and phyllotaxy, Curr. Op.in Plant Biol., 8, 53-58
- Shipman PD and Newell AC, Phyllotactic patterns on plants, Phys Rev Lett 92 (2004), p. 168102.
- Green PB (1999) Expression of pattern in plants: combining molecular and calculus-based biophysical paradigms. Am. J. Bot. 86: 1059-1076, figure 3
- Coen E, Rolland-Lagan AG, Matthews M, Bangham A and Prusinkiewicz P (2004) The genetics of geometry. Proc. Natl. Acad. Sci. USA 101,4728 –4735.
- Dumais J, Kwiatkowska D (2002) Analysis of surface growth in shoot apices. Plant Journal. 31, 229-241.
- Kwiatkowska D, Dumais J (2003) Growth and morphogenesis at the vegetative shoot apex of Anagallis arvensis L. Journal of Experimental Botany. 54, 1585-1595.
- Fleming AJ, McQueen-Mason S, Mandel T, Kuhlemeier C (1997) Induction of leaf primordia by the cell wall protein expansin. Science. 276: 1415–1418.
- Wyrzykowska J, Pien S, Shen WH and Fleming AJ, (2002) Manipulation of leaf shape by modulation of cell division, Development. 129, 957–964.
- Reinhardt D, Mandel T, Kuhlemeier C. (2000) Auxin Regulates the Initiation and Radial Position of Plant Lateral Organs. Plant Cell. 12: 507-518
- Cette hypothèse fut soutenue pendant une longue période et on en retrouve la trace dans nombre de revues sur ce sujet. Bien qu’intuitive, la prolifération cellulaire serait un moteur pour la croissance, les données de Fleming et Wyrzykowska indiquent clairement qu’une activité cellulaire accrue n’est pas une condition suffisante au déclenchement de l’organogénèse.
- Reinhardt D, Pesce E, Stieger P, Mandel T, Baltensperger K, Bennett M, Traas J, Friml J and Kuhlemeier C (2003) Regulation of phyllotaxis by polar auxin transport. Nature 426, 256-260.
- Puisque la limite de l’angle de divergence est un irrationnel et donc que les feuilles ne se recouvrent jamais entièrement, limitant ainsi les phénomènes d’ombrage.
Voir aussi
Bibliographie
- Steeves TA and Sussex IM Patterns in plant development. Cambridge University Press, Cambridge, 1989.
- Cronk QCB (2001) Plant evolution and development in a post-genomic context. Nat. Rev. Gev. 2 (8): 607-619.
- Byrne ME, Barley R, Curtis M, Arroyo JM, Dunham M, Hudson A, Martienssen RA (2000) Asymmetric leaves1 mediates leaf patterning and stem cell function in Arabidopsis, Nature, 408 (6815): 967-971 DEC 21 2000.
- Adler I, Barabé D et Jean RV (1997) A history of the study of phyllotaxis. Ann. Bot. 80: 231-244.
- Douady S and Couder Y (1996) Phyllotaxis as a Dynamical Self Organizing Process (Part I, II, III), J. Theor. Biol. 139, 178-312.
- Selker JML, Stichic GL and Green PB (1992) Biophysical mechanisms for morphogenetic progressions at the shoot apex. Dev. Biol. 153, 29-43.
- Green PB (1999) Expression of pattern in plants: combining molecular and calculus-based biophysical paradigms. Am. J. Bot. 86: 1059-1076
- Green PB, Steele CS and Rennich SC (1996) Phyllotactic patterns: a biophysical mechanism for their origin. Annals of Botany 77: 515–527.
- Fleming AJ (2005) Formation of primordia and phyllotaxy, Curr. Op.in Plant Biol., 8, 53-58
- Shipman PD and Newell AC, Phyllotactic patterns on plants, Phys Rev Lett 92 (2004), p. 168102.
- Coen E, Rolland-Lagan AG, Matthews M, Bangham A and Prusinkiewicz P (2004) The genetics of geometry. Proc. Natl. Acad. Sci. USA 101,4728 –4735.
- Dumais J, Kwiatkowska D (2002) Analysis of surface growth in shoot apices. Plant Journal. 31, 229-241.
- Kwiatkowska D, Dumais J (2003) Growth and morphogenesis at the vegetative shoot apex of Anagallis arvensis L. Journal of Experimental Botany. 54, 1585-1595.
- Fleming AJ, McQueen-Mason S, Mandel T, Kuhlemeier C (1997) Induction of leaf primordia by the cell wall protein expansin. Science. 276: 1415–1418.
- Wyrzykowska J, Pien S, Shen WH and Fleming AJ, (2002) Manipulation of leaf shape by modulation of cell division, Development. 129, 957–964.
- Wyrzykowska J, Fleming A (2003) Cell division pattern influences gene expression in the shoot apical meristem. Proc. Natl. Acad. Sci. USA, 100:5561-5566.
- Reinhardt D, Mandel T, Kuhlemeier C. (2000) Auxin Regulates the Initiation and Radial Position of Plant Lateral Organs. Plant Cell. 12: 507-518
- Reinhardt D, Pesce E, Stieger P, Mandel T, Baltensperger K, Bennett M, Traas J, Friml J and Kuhlemeier C (2003) Regulation of phyllotaxis by polar auxin transport. Nature 426, 256-260.
- B Veit, SP Briggs, RJ Schmidt, MF Yanofsky, S Hake (1998) Regulation of leaf initiation by the terminal ear 1 gene of maize. Nature
- Paquet N, Bernadet M, Morin H, Traas J, Dron M, Charon C. (2005) Expression patterns of TEL genes in Poaceae suggest a conserved association with cell differentiation. J Exp Bot. 56(416): 1605-14.
Liens externes
- [en] site du Smith college dédié à la phyllotaxie avec notamment une section qui reprend panneau par panneau l'ensemble de l'exposition originale








Wikimedia Foundation. 2010.