- Cténaire
-
Ctenophora
Cténophores

Ctenophorae
dans Kunstformen der Natur, 1904
de Ernst HaeckelClassification classique Règne Animalia Sous-règne Eumetazoa Embranchement Ctenophora
Eschscholtz, 1829Classes de rang inférieur Voir texte Position phylogénétique  Retrouvez ce taxon sur Wikispecies
Retrouvez ce taxon sur Wikispecies
D'autres documents multimédia
sont disponibles sur CommonsParcourez la biologie sur Wikipédia : 





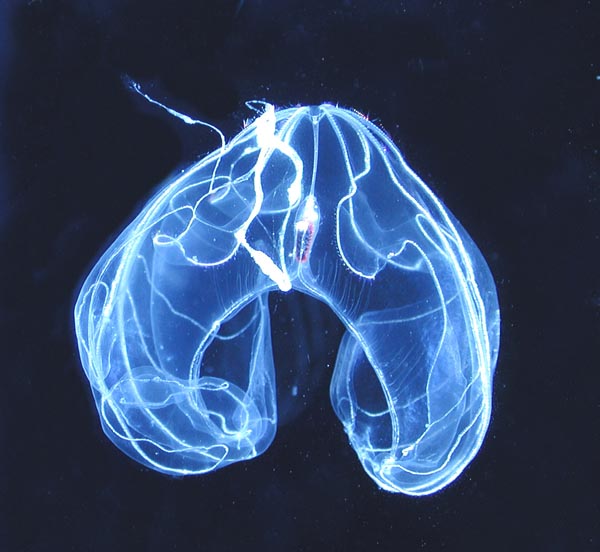
Les cténophores (Ctenophora, du grec ktenos, « peigne » et phorein, « porter ») ou « Cténaires » sont des organismes marins représentés par près de 150 espèces, répandus dans tous les océans du monde.
Leur nom provient de la structure en peigne de rangées de cils locomoteurs. Leur ressemblance superficielle avec les méduses les a fait autrefois classer auprès de ces dernières dans le groupe des cœlentérés, classification aujourd'hui tombée en désuétude. Contrairement aux méduses, les Ctenophora ne possèdent pas de cnidocytes (cellules urticantes), mais des colloblastes (cellules collantes); ils sont de plus hermaphrodites.
Ce sont des organismes prédateurs le plus souvent planctoniques ; ils constituent une bonne part de la biomasse planctonique mondiale.
Les cténaires les plus connus sont la groseille de mer (Pleurobrachia pileus) et la ceinture de Vénus (Cestum veneris). Certaines espèces, comme la groseille de mer, peuvent atteindre des populations si nombreuses qu'elles colmatent les filets des pêcheurs ; d'autres ne sont connues que par quelques spécimens.Sommaire
Morphologie
Aspect général
Les cténophores ont une forme ovoïde, rubanée, en poire ou en cloche. Translucides et gélatineux, ils ont le plus souvent 8 rangées de cils vibratiles soudés à la base, formant des peignes locomoteurs, et la plupart portent 2 grands tentacules.
Les cténophores sont extrêmement fragiles, donc très difficiles à prélever et à étudier.
Les espèces les plus communes ne font souvent que quelques centimètres de long. Exceptionnellement, les espèces du genre Cestum peuvent atteindre 1,5 m.
Coloration
 La lumière se diffracte sur les cténidies de ce Mertensia ovum, produisant une iridescence
La lumière se diffracte sur les cténidies de ce Mertensia ovum, produisant une iridescence
Les Cténophores sont généralement translucides et incolores, mais certaines espèces peuvent présenter de la couleur : rouge, orange voire noir. Les rangées de peignes locomoteurs (cténidies) sont de plus souvent iridescentes, par diffraction de la lumière.
Les espèces d'eau profonde, comme la "Red tortuga" (qui ne possède pas actuellement de nom scientifique), peuvent apparaître vivement colorées grâce à des pigments qui absorbent certaines longueurs d'ondes de la lumière visible (les radiations bleues). Elle apparaissent donc rouge sombre dans la mer.
De nombreuses espèces de Ctenophora sont par ailleurs capables d'émettre de la lumière par bioluminescence. Une espèce en particulier, Eurhamphaea vexilligera, peut produire une sorte d'encre rouge qui émet une lueur bleue dans l'obscurité, peut-être pour dissuader les prédateurs.
Anatomie
Symétrie et organisation interne
Les Ctenophora ont une symétrie intéressante, nommée symétrie biradiaire, mais ils présentent quelques structures asymétriques, comme les pores anaux, le statocyste ou parfois les rangées de peignes natatoires. Cette symétrie est compliquée d'une symétrie bilatérale chez les Tentaculata.
Leur organisation est diploblastique (ils ne possèdent que deux feuillets embryonnaires transparents).
Le feuillet externe, ou ectoderme, donne naissance à l'épiderme externe, constitué de deux couches de cellules, en grande partie recouvert par une couche protectrice de substance visqueuse excrétée par des cellules glandulaires. L'endoderme (épiderme interne) entoure une cavité qui sert d'estomac, reliée à l'extérieur et à l'ouverture de la bouche par un œsophage long et étroit. Les proies capturées sont prédigérées dans l'œsophage par de puissantes enzymes, puis leur hydrolyse s'achève dans l'estomac. Il n'y a pas de véritable « sortie » de l'estomac, à part deux pores anaux qui, malgré leur nom, ne sont que modérément utilisés pour l'excrétion des déchets indigestibles (principalement rejetés par la bouche).
L'espace situé entre les épidermes externe et interne est occupé par la mésoglée, une couche gélatineuse visqueuse et transparente formée de collagène et de tissu conjonctif. L'important réseau de protéine structurale est maintenu par des cellules particulières ressemblant à des amibes (amibocytes). La mésoglée est parcourue par de petits canaux utilisés pour le transport et le stockage des nutriments. La position de ces canaux varie en fonction des espèces, mais ils se trouvent principalement en dessous des tissus qu'ils desservent.
La mésoglée pourrait aussi jouer un rôle dans la flottabilité de ces animaux.
Les cils vibratiles situés dans les canaux du système digestif pourrait servir à pomper l'eau vers l'intérieur ou l'extérieur de la mésoglée, pour ajuster la pression osmotique interne, par exemple lorsque le cténophore passe d'un milieu strictement marin à de l'eau plus saumâtre côtière.Les Cténophores ne possèdent pas de système circulatoire différencié, ni d'organe respiratoire, ni de système excréteur. Les échanges gazeux et l'excrétion des déchets du métabolisme comme l'ammoniaque se réalise au niveau de la surface entière de l'organisme par simple diffusion.
La mésoglée est aussi parcourue par un réseau simple de cellules nerveuses (neurones), sans centre nerveux différencié. Ces neurones sont concentrés autour de la bouche, des tentacules (quand ils existent), des cténidies (rangées de peignes locomoteurs) et du statocyste. Ils sont reliés aux cellules musculaires de la mésoglée et avec la couche de cellules la plus interne de l'épiderme externe. Cette organisation complexe de la mésoglée lui confère, selon certains auteurs, le statut de mésenchyme.
Statocyste et peignes locomoteurs
Il s'agit d'un système spécialisé servant aux cténophores d'organe d'équilibration et qui contrôle aussi leurs mouvements. Il se trouve à l'extrémité opposée de celle de l'ouverture de la bouche. Il est formé d'un ensemble formé d'une centaine de cellules calcifiées appelées statolithes reposant en équilibre sur leur flagelles sinueux, répartis en quatre groupes horizontaux. Lorsque les conditions extérieures poussent le cténophore à changer de position, le statolithe exerce davantage de pression sur un des quatre groupes de flagelles et moins sur les trois autres. Cette information sensorielle est transmise à l'ectoderme via le réseau de neurones. Le cténophore ajustera donc l'activité de ses rangées de peignes locomoteurs en réponse à cette stimulation.
Ces peignes sont généralement disposés en 8 rangées allongées dans le sens longitudinal, appelées cténidies. Chaque peigne est formé d'une rangée de plusieurs centaines de cils vibratiles, fusionnés à la base, formant ainsi une sorte de palette natatoire de 2 à 5 mm de long. En soulevant ces peignes successivement, le cténophore les utilise comme des rames qui, quand les huit cténidies sont convenablement synchronisées, lui permettent de se propulser dans l'eau. Chacun des quatre groupes de cellules flagellées du statocyte contrôle un quart de l'animal, soit 2 rangées de peignes. Le rythme de battement et la synchronisation des peignes se propage automatiquement, de façon mécanique, sans intervention des cellules nerveuses. La position dans l'espace du cténophore (géotaxis) provoque l'action de la gravité sur le statocyste, augmentant ou diminuant la fréquence des battements. Le cténophore peut aussi modifier le rythme de battements des rangées de peignes pour nager vers le haut ou le bas de la colonne d'eau. La position dans l'espace est déterminée par le cténophore grâce au statocyste et au réseau nerveux, mais aussi grâce à la détermination de la quantité de lumière ambiante.
Tentacules
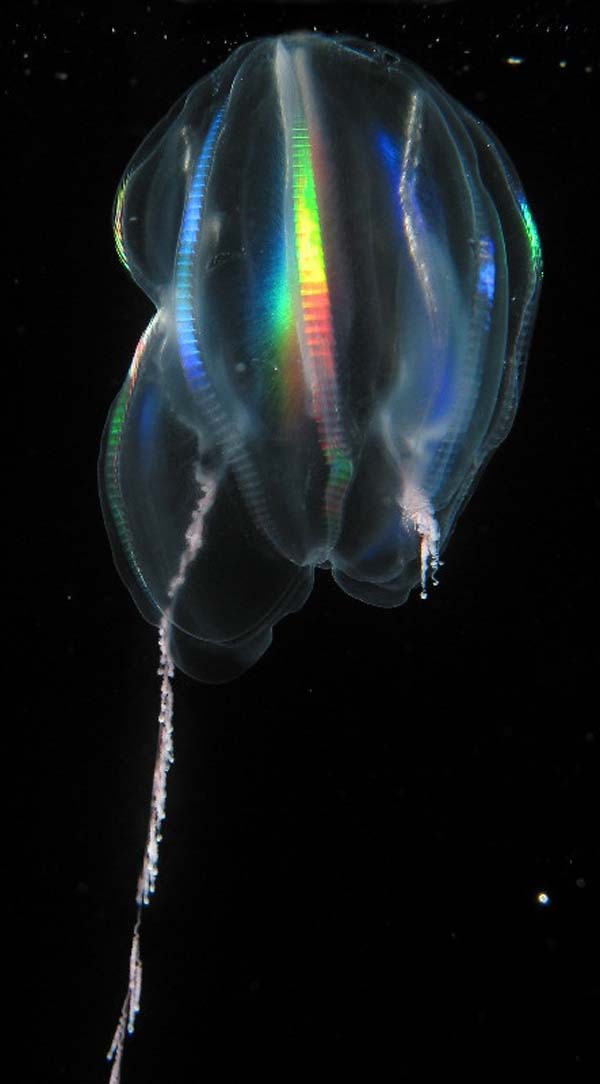
 Espèce non déterminée de cténophore présentant deux tentacules ramifiés
Espèce non déterminée de cténophore présentant deux tentacules ramifiésDe nombreuses espèces de cténophores possèdent deux tentacules implantés vers le milieu du corps, opposés par rapport à l'axe de symétrie. Ils sont utilisés pour capturer les proies ; chaque tentacule peut se rétracter entièrement par enroulement dans une poche située à sa base, la gaine tentaculaire. Ils ont une croissance continue par la base, qui compense l'usure de l'extrémité. Ces tentacules portent des ramifications appelées tentilles qui, contrairement aux tentacules des Cnidaires, ne contiennent pas de cellules urticantes (cnidocytes), mais des cellules collantes (colloblastes). Ces cellules se déploient brutalement quand une proie entre en contact avec un tentacule : le filament cytoplasmique en spirale qui constitue leur base se détend alors, projetant la partie apicale collante de la cellule vers la proie, qui sera alors capturée. Contrairement aux cnidocytes, les colloblastes ne sont pas détruits après usage : grâce à l'élasticité du filament cytoplasmique, ils peuvent, une fois débarrassés de la proie, reprendre leur position initiale[1].
Biologie et comportement
Locomotion
De nombreux cténophores se laissent simplement porter par les courants. Ils peuvent cependant nager, parfois assez rapidement, grâce aux battements de leurs peignes locomoteurs. Ce sont les plus gros animaux se déplaçant à l'aide de cils vibratiles. Ils peuvent atteindre une vitesse de 5 cm/s. Les battements coordonnés des peignes confèrent à cet animal une nage sans à-coups, ce qui pourrait être un avantage au niveau de l'évolution, car cette nage ne génère pas de vibrations susceptibles d'alerter les proies ou les prédateurs.
Certaines espèces nagent grâce aux battements de leurs lobes oraux lorsqu'ils fuient, d'autres se déplacent grâce à des ondulations de leur corps, voire rampent comme des vers plats.
Alimentation
Mode d'alimentation
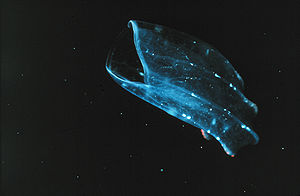 Un cténophore (genre Beroe) en quête de nourriture. La bouche se situe à gauche.
Un cténophore (genre Beroe) en quête de nourriture. La bouche se situe à gauche.Les Ctenophora sont des animaux prédateurs qui utilisent leurs tentacules pour capturer leurs proies (plancton, larves d'animaux marins, vers marins, crustacés (comme les copépodes), Cnidaires, autres Ctenophora, et parfois petits poissons. Quand leurs tentacules ont capturé une proie, ils sont rétractés vers la bouche et débarrassés de la proie. La nourriture est alors ingérée. La progression vers l'estomac se fait soit à l'aide de mucus, soit par l'action de cils vibratiles internes, situés dans le système digestif.
Les cténophores ne possèdent pas tous des tentacules. Certains, comme le Beroe, ingèrent directement des proies gélatineuses. D'autres utilisent les lobes musculeux qui entourent leur bouche pour capturer les proies.
Cas particuliers
Les espèces du genre Haeckelia se nourrissent presque exclusivement de cnidaires, mais ne digèrent pas leurs cnidocystes. Ces derniers seront réutilisés par incorporation au niveau de l'épiderme de leurs propres tentacules (kleptocnides). Ce "vol" a pendant longtemps désarçonné les zoologistes, qui présumèrent à tort que certains cténophores pouvaient eux aussi produire des cnidocytes.
Le parasitisme n'a été observé que dans un seul genre, Lampea, qui parasite les salpes quand il est trop petit pour les ingérer en entier.
Reproduction
 Larve appartenant à un individu du genre Bolinopsis
Larve appartenant à un individu du genre BolinopsisÀ l'exception de quelques espèces de l'ordre des Platyctenida, la reproduction est sexuée.
Presque tous les cténophores sont hermaphrodites, possédant à la fois les organes reproducteurs mâle et femelle, situés directement en dessous des cténidies, à proximité des canaux de la mésoglée. Cependant, le genre Ocyropsis, de l'ordre des Lobata, présente des sexes séparés.
A maturité, les produits sexuels sont émis dans la mer par de petites ouvertures dans l'ectoderme nommées « gonopores ». Le déclenchement de l'émission des gamètes est stimulé par certaines conditions lumineuses. La fécondation est donc externe ; l'autofécondation est rare. L'espèce Tjalfiella tristoma, de l'ordre des Platyctenida, est cependant vivipare : les larves se développent dans une cavité de l'organisme parental.
Certaines espèces de cténophores, comme Beroe ovata, ont une méthode particulière pour éviter la polyspermie. Lors de la fécondation, plusieurs spermatozoïdes mâles ont pénétré l'ovule. Le pronucleus femelle se met alors à migrer dans le cytoplasme de l'ovule, et choisit le pronucleus mâle avec lequel il fusionnera, les autres seront rejetés. Le principal critère de sélection semble être l'absence de consanguinité.
Après fécondation, la cellule-œuf commence à se diviser. La segmentation de la cellule-œuf est totale, inégale et à détermination précoce. Dès la deuxième division de la cellule-œuf (stade 4 cellules), on voit apparaître la symétrie radiaire. L'embryon se développe alors en une larve cydippide nageuse. Les cydippides sont très semblables entre eux, même quand ils appartiennent à d'autres espèces de cténophores. Certains auteurs critiquent l'appellation de « larve », car parfois, le cydippide ressemble fortement à l'adulte. Dans certains ordres par exemple, comme les Lobata and Platyctenida, la larve cydippide et l'adulte ne sont pas différents morphologiquement, de sorte que l'appellation "larve" est dans ce cas guère appropriée.
La larve subira une métamorphose plus ou moins importante selon les espèces, parfois inexistante, pour passer à l'état adulte. Les cténophores sont monomorphes : ils ne présentent jamais de stade fixé.
L'extrême fragilité de leur constitution rend la recherche sur leur mode de vie très difficile ; c'est pourquoi il n'y a pas de données sur leur durée de vie. Mais il est connu que les cténophores commencent à se reproduire à un âge précoce, et on peut ainsi supposer qu'ils ont un cycle de génération court.
Régénération
Les cténophores ont d'extraordinaires capacités de régénération ; même si la moitié de l'organisme est détruit, la partie restante peut reformer un individu complet. Certains organes, comme le statocyste, peuvent se régénérer même s'ils sont entièrement détruits.
Répartition et habitat
Habitat
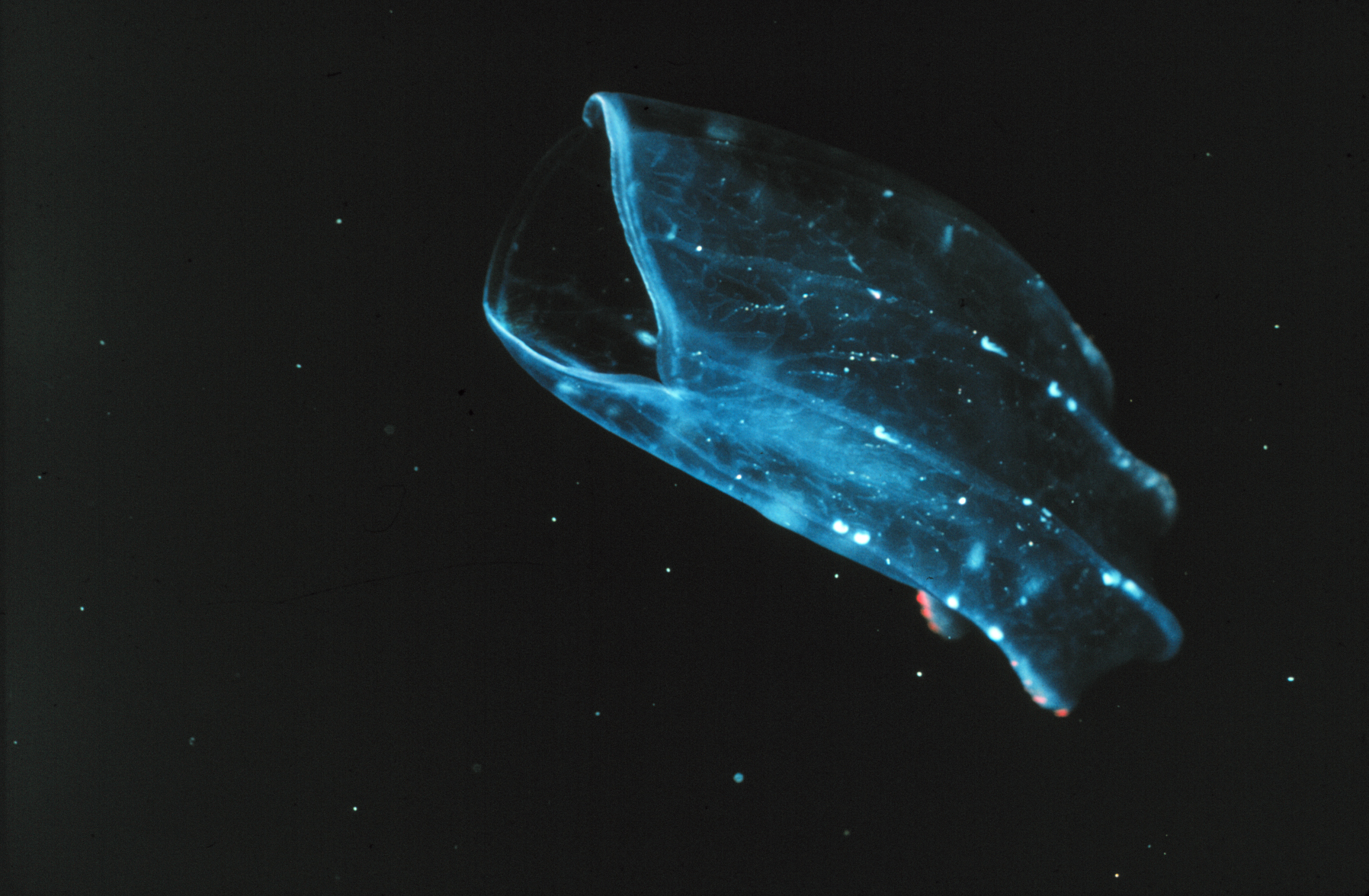
 Deux cténophores du genre Mnemiopsis
Deux cténophores du genre MnemiopsisTous les cténophores sont marins, il n'y a pas d'espèces d'eau douce. Ils vivent à des profondeurs variant de la surface de la mer jusqu'à -4 000 m.
Certaines espèces peuvent passer sans dommage des eaux océaniques aux eaux saumâtres des estuaires.
Les espèces les mieux connues vivent au milieu du plancton dans les couches océaniques situées près de la surface. Cependant, comme ils sont majoritairement transparents, extrêmement fragiles et ne dépassent que rarement quelques centimètres en longueur, ils sont généralement inconnus du grand public. Les promeneurs peuvent éventuellement rencontrer des espèces du genre Pleurobrachia (groseille de mer) échouées sur la plage. Les genres Bolinopsis, Mnemiopsis et Beroe peuvent aussi être rencontrés assez fréquemment.
Environ 35 espèces de cténophores sont benthiques et vivent sur le fond marin. Ces espèces font partie de l'ordre des Platyctenida, appelé ainsi car ses membres ont une forme aplatie qui les fait davantage ressembler à des limaces de mer ou à des vers plats.
Répartition
En tant que membre du plancton, les cténophores sont sujets à de grands déplacements au gré des courants marins, même si certaines espèces sont inféodées à un habitat particulier. On peut les trouver dans toutes les mers et océans du globe. Ils sont particulièrement abondants sous les tropiques et aux deux pôles.
Le cténophore Mertensia ovum est un des membres prédominants du plancton des eaux arctiques.
Espèces invasives
Bien que les cténophores soient généralement discrets et que leur influence sur un écosystème soit de toute évidence assez faible, ils peuvent tout de même provoquer des dommages quand ils apparaissent dans des eaux dont ils ne sont pas natifs.
L'espèce Mnemiopsis leidyi est originaire de l'ouest de l'Atlantique, entre Cape Cod, USA (41° N, 70° W) au nord et la Peninsule de Valdez, Argentine (43° S, 64° W) au sud. Elle fut accidentellement importée en Mer Noire dans les années 1980, transportée dans l'eau contenue dans des ballasts de navires[2]. Sur place, l'espèce s'est rapidement répandue. En dix ans, l'industrie de pêche à l'anchois en mer Noire a subi une forte récession, car le cténophore nouvellement introduit se nourrit du même plancton que la larve d'anchois, qui s'est ainsi retrouvée affamée par cette concurrence. La biomasse de cténophores en mer Noire a atteint jusqu'à un million de tonnes.
Depuis la similaire soudaine apparition, en 1997, d'un autre cténophore étranger en mer Noire, Beroe ovata (qui se nourrit de Mnemiopsis leidyi), l'équilibre fut en quelque sorte rétabli ; mais la mer Noire est dorénavant occupée par deux espèces allochtones. Le même scénario, avec les mêmes espèces, est en train de se dérouler en Mer Caspienne depuis les années 1998-1999, et Mnemiopsis leidyi a été signalée en Mer du Nord en 2006. On estime donc que le même scénario est à redouter en mer Baltique, où les scientifiques finlandais ont découvert que Mnemiopsis leidyi avait survécu à l'hiver et se multipliait rapidement : une expédition récente (2007) a mesuré 600 cténophores par mètre cube dans la mer Baltique[3]
Prédateurs et parasites
Plusieurs animaux sont des prédateurs des cténophores. On trouve parmi eux des cnidaires, des tortues marines, divers poissons (comme par exemple le saumon, le maquereau et le lump), des oiseaux de mer, et même les autres espèces de cténophores.
Les cténophores sont souvent infestés de crustacés parasites du groupe des Amphipodes.
Histoire de la taxinomie et de la phylogénie des cténophores
Fossiles de cténophores
Le corps mou des cténophores rend très improbable la fossilisation, de sorte que la phylogénie des cténophores fossiles est très peu documentée.
Parmi les fossiles appartenant de façon indéniable au groupe des cténophores, on peut citer Archaeocydippida hunsrueckiana et Paleoctenophora brasseli, datant du Dévonien ; ces fossiles, conservés dans le schiste à grain très fin de Hunsrück, présentent de nombreux détails permettant une identification.
Il y a par contre des discussions en ce qui concerne l'espèce Maotianoascus octonarius, pour savoir si ce fossile de la faune de Chengjiang (datant du Cambrien inférieur) est un cténophore ou non. Dans la faune de Burgess, datant du Cambrien, on trouve trois espèces qui appartiennent sans doute au groupe des cténophores : Ctenorhabdotus capulus (qui possédait 24 tentacules), Fasciculus vesanus et Xanioascus canadensis.
Première classification
Les marins avaient observé des cténophores depuis fort longtemps, mais la première observation consignée par écrit, réalisée par un médecin de bord, ne date que de 1671. Le taxonomiste suédois Carl von Linné classa les cténophores avec les autres invertébrés « primitifs » (tels que les spongiaires ou les cnidaires) sous le nom de zoophytes (« animal-plante »), faisant ainsi allusion à leur mode de vie passif, en apparence « végétatif ». Le zoologiste français Georges Cuvier soutint cette classification. Ce n'est qu'au XIXe siècle que les cténophores furent reconnus comme un taxon à part entière.
Place du taxon dans la systématique
La classification classique des cténophores plaçait ce groupe dans les Cœlentérés (Coelenterata), au côté des cnidaires :
Classification classique
- Eumetazoa
- Coelenterata
- Cnidaria
- Ctenophora
- Bilateria
- Coelenterata
Cette classification a été détrônée par les apports de la cladistique, méthode actuellement utilisée pour reformer la classification des espèces, apports qui montrent que les cténophores sont plus proches de Bilateria (animaux à symétrie bilatérale) que des cnidaires (Cnidaria). Les critères retenus pour étayer ce rapprochement est l'existence de tentacules symétriques par rapport à un plan, ce qui complique leur symétrie biradiée d'une symétrie bilatérale. Autre argument, la forme des spermatozoïdes de cténophores, qui ne possèdent qu'un seul grand acrosome, est plus proche de celle des spermatozoïdes des Bilateria. De plus, les cténophores possèdent un vrai tissu musculaire.
Les cténophores diffèrent donc des Cnidaires sur ces trois points (les cnidaires ne possèdent ni véritable tissu musculaire, ni ébauche de symétrie bilatérale, et leurs spermatozoïdes possèdent plusieurs vésicules acrosomales).Pour ces raisons, une nouvelle classification a été proposée, plaçant le groupe des cténophores dans les Acrosomata[4], au côté des Bilateria :
Nouvelle classification
- Eumetazoa
- Cnidaria
- Acrosomata
- Ctenophora
- Bilateria
Cette approche présupposait que les cténophores étaient le groupe frère des bilatériens, or d'autres études[5] [6] considèrent les cnidaires plus proches des bilatériens que les cténophores.
Il a aussi été aussi suggéré que les cténophores pourraient avoir une parenté avec les vers plats, à cause entre autres des similarités existant entre ces derniers et les cténophores benthiques de forme aplatie de l'ordre des Platyctenida. Mais d'autres zoologistes considèrent cette ressemblance trop superficielle pour justifier une parenté.
Classification des différents ordres de Ctenophora
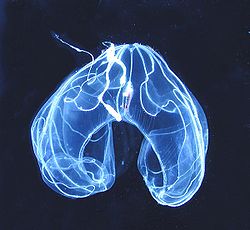 Bathocyroe fosteri, de l'ordre des Lobata, est un cténophore commun dans les eaux profondes. La bouche est orientée vers le bas.
Bathocyroe fosteri, de l'ordre des Lobata, est un cténophore commun dans les eaux profondes. La bouche est orientée vers le bas.C'est un groupe encore assez mal connu qui pose un problème dans la classification phylogénétique car nous ne possédons que très peu de données et certains gènes ont un taux d'évolution lent comparé aux bilatériens.
De nos jours, une centaine d'espèces sont connues et décrites ; elles sont traditionnellement divisées en deux classes : les Tentaculata (aussi nommés Tentaculifera) et les Nuda (aussi nommés Atentaculata).
- Les Tentaculata sont constitués et de loin par le plus grand nombre d'espèces. Ils possèdent des tentacules, parfois vestigiaux. Cette classe est divisée en 6 ordres :
- Les Nuda ne contiennent qu'un seul ordre, Beroida, auquel appartient le genre Beroe. Ils ne possèdent pas de tentacules, pas même à l'état vestigial.
Du fait de l'incertitude passée et actuelle sur la classification des cténophores, il n'est pas certain que la classification ci-dessus reflète la véritable phylogénie des cténophores. Des études moléculaires indiquent que la classe Cydipidda est un groupe paraphylétique, ce qui signifie qui n'inclut pas tous les descendants de leur ancêtre commun. La classification ci-dessus serait donc à réviser.
Le diagramme suivant montre une phylogénie des cténophores basée sur des critères morphologiques et sur des données moléculaires concernant l'ARN :
Ctenophora |--Cydippida (Mertensiidae) |-- |--Platyctenida |-- |--Cydippida (Pleurobrachidae) |-- | |--Nuda Beroida | |--Cydippida (Haeckeliidae) | |-- |--Lobata |--Cestida |--ThalassocalycidaCette classification n'est pas confirmée. Pour le moment, la phylogénie des cténophores doit être considérée comme non établie.
Classifications selon trois organismes en 2008 Selon ADW : Selon ITIS : Selon NCBI : - classe Nuda
- ordre Beroida
- ordre Thalassocalycida
- classe Tentaculata
- classe Nuda Chun, 1879
- ordre Beroida
- classe Tentaculata Eschscholtz, 1825
Liens et références
Notes et références
- ↑ Beaumont A et Cassier P, Biologie animale, tome 1, Dunod université, Paris, 1981 (réimpr. 1991), 147-153 p. (ISBN 2-04-011432-7)
- ↑ Invasion of the Caspian Sea by the comb jelly Mnemiopsis, First international meeting, Baku (Azerbaijan), Avril 2001
- ↑ Internytt, 2007-08-27
- ↑ Peter Ax, 1995, Das System der Metazoa I-III. Ein Lehrbuch der phylogenetischen Systematik, Gustav Fischer Verlag, Stuttgart/Spektrum Akademischer Verlag, Heidelberg, (ISBN 3-8274-0972-1); 1996, Multicellular animals: a new approach to the phylogenetic order in nature, Springer, (ISBN 3-540-60803-6)
- ↑ Allen G. Collins, 1998, Evaluating multiple alternative hypotheses for the origin of Bilateria: an analysis of 18S rRNA molecular evidence. Proc. Natl. Acad. Sci. USA 95:1545863
- ↑ Mónica Medina, Allen G. Collins, Jeffrey D. Silberman & Mitchell L. Sogin, Evaluating hypotheses of basal animal phylogeny using complete sequences of large and small subunit rRNA.
Bibliographie
- Beaumont A et Cassier P, Biologie animale Tome 1, Dunod université, Paris, 1981 (réimpr. 1991), 147-153 p. (ISBN 2-04-011432-7)
- D. T. Anderson, Invertebrate Zoology, 2nd ed, Oxford Univ. Press, 2001, Ch. 3, p. 54, ISBN 0-19-551368-1
- R. S. K. Barnes, P. Calow, P. J. W. Olive, D. W. Golding, J. I. Spicer, The invertebrates – a synthesis, 3rd ed, Blackwell, 2001, ch. 3.4.3, p. 63, ISBN 0-632-04761-5
- R. C. Brusca, G. J. Brusca, Invertebrates, 2nd Ed, Sinauer Associates, 2003, ch. 9, p. 269, ISBN 0-87893-097-3
- J. Moore, An Introduction to the Invertebrates, Cambridge Univ. Press, 2001, ch. 5.4, p. 65, ISBN 0-521-77914-6
- E. E. Ruppert, R. S. Fox, R. P. Barnes, Invertebrate Zoology – A functional evolutionary approach, Brooks/Cole 2004, ch. 8, p. 181, ISBN 0-03-025982-7
- W. Schäfer, Ctenophora, Rippenquallen, in W. Westheide and R. Rieger: Spezielle Zoologie Band 1, Gustav Fischer Verlag, Stuttgart 1996
- Bruno Wenzel, Glastiere des Meeres. Rippenquallen (Acnidaria), 1958, ISBN 3-7403-0189-9
- Harbison, G. R. 1985. On the classification and evolution of the Ctenophora. pp 78-100 in The Origins and Relationships of Lower Invertebrates. (S. C. Morris, J. D. George, R. Gibson and H. M. Platt, eds.). Oxford University Press, Oxford.
- M. Q. Martindale, J. Q. Henry, Ctenophora, in S. F. Gilbert, A. M. Raunio, Embryology: Constructing the Organism, Sinauer, 1997, p. 87
- C.E. Mills. Internet 1998-present. Phylum Ctenophora: list of all valid species names. Web page established March 1998, last updated (see date at end of page).
- M. Podar, S. H. D. Haddock, M. L. Sogin, G. R. Harbison, A molecular phylogenetic framework for the phylum Ctenophora using 18S rRNA genes, Molecular Phylogenetics and Evolution, 21, 2001, p. 218
- T. A. Shiganova, Invasion of the Black Sea by the ctenophore Mnemiopsis leidyi and recent changes in pelagic community structure, Fisheries Oceanography, 1998, p. 305
- G. D. Stanley, W. Stürmer, The first fossil ctenophore from the lower devonian of West Germany, Nature 303, 1983, p. 518
Références taxonomiques
- Référence Tree of Life Web Project : Ctenophora (en)
- Référence Animal Diversity Web : Ctenophora (en)
- Référence ITIS : Ctenophora Eschscholtz, 1829 (fr) ( (en))
- Référence NCBI : Ctenophora (en)
Liens externes
- University of Washington - Ctenophores
- (en)Introduction to Ctenophora, University of California Museum of Paleontology
- Ctenophores from the São Sebastião Channel, Brazil
- Video of ctenophores at the National Zoo in Washington DC
- Mills C.E. (1998). Phylum Ctenophora: list of all valid species names[1]
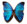 Portail de la zoologie
Portail de la zoologie
Catégories : Plancton | Ctenophora - Eumetazoa



Wikimedia Foundation. 2010.