- Transmission synaptique
-
Synapse et transmission synaptique
Neurosciences 
Niveaux d'analyse Moléculaire • Synaptique • Neuronal • Réseau neuronal • Organique • Systémique
Méthodes Imagerie cérébrale • Électrophysiologie • Lésion cérébrale • Intelligence artificielle
Branches d'études Neuroanatomie • Neurophysiologie • Neuroendocrinologie • Psychophysiologie • Neurosciences cognitives •
Concepts majeurs Neurone • Potentiel d'action • Synapse et transmission synaptique • Réseau neuronal • Neuromédiateur • Plasticité neuronale • Plasticité synaptique • Précablage • Réflexe • Récompense • Cognition • Modularité de l'esprit
Chercheurs Ramón y Cajal • C.S. Sherrington • P. Broca • J. Olds • J. LeDoux • D.H. Hubel • T. Wiesel • E. Candel • J.P. Changeux
Champs d'application Neurologie • Neurochirurgie • Neuropsychologie • Psychiatrie • Neuropharmacologie • Chronobiologie •
Voir aussi Catégorie Neurosciences
- L'adjectif synaptique redirige ici. Pour le logiciel (gestionnaire de paquet), voir Synaptic. Pour l'entreprise (fabriquant de pavé tactile), voir Synaptics.
- L'adjectif synaptique possède également un paronyme : synoptique.

La synapse (du grec. syn = ensemble; haptein = toucher, saisir; c'est-à-dire connexion) désigne une zone de contact fonctionnelle qui s'établit entre deux neurones, ou entre un neurone et une autre cellule (cellules musculaires, récepteurs sensoriels...). Elle assure la conversion d'un potentiel d'action déclenché dans le neurone présynaptique en un signal dans la cellule postsynaptique. On estime, pour certains types cellulaires (par exemple cellule pyramidale, cellule de Purkinje...), qu'environ 40 % de la surface membranaire est couverte de synapses.
On distingue habituellement deux types de synapses :
- la synapse chimique, très majoritaire, qui utilise des neurotransmetteurs pour transmettre l'information ;
- la synapse électrique où le signal est transmis électriquement par l'intermédiaire d'une jonction communicante (en anglais gap-junction).
On les distingue au microscopie électronique par la taille de la fente synaptique ; de l'ordre de 2 nanomètres pour les synapses électriques, entre 10 et 40 nm pour les synapses chimiques. On peut également, dans le cas des synapses électriques, observer les jonctions communicantes. Au niveau d'une synapse, il s'agit toujours d'un contact entre deux membranes plasmiques, il n'y a jamais fusion en un syncitium.
Histoire des sciences : synapses et réfutation de la théorie réticulariste
La découverte du mode de fonctionnement du système nerveux a eu lieu au XIXe siècle. Camillo Golgi inventa un colorant permettant une teinture optimale du neurone et de ses prolongements cellulaires.
Par la suite, Santiago Ramón y Cajal reprit les méthodes de coloration de Golgi et proposa le concept de neurone. Le terme de « synapse », quant à lui, fut proposé en 1897 par le physiologiste et Prix Nobel britannique Sir Charles Scott Sherrington pour désigner le point de contact entre deux neurones. Toutefois, Golgi était lui-même opposé à l'hypothèse selon laquelle le système nerveux pouvait être composé d'unités discontinues. En 1906 Golgi et Cajal reçurent conjointement le Prix Nobel de médecine et physiologie, pour deux théories de l'organisation du tissu neuronal (neuronisme et réticularisme). Il a été demontré par la suite que les synapses électriques sont très rares et l'on admet aujourd'hui que le système nerveux est constitué majoritairement d'unités contiguës (thèse neuroniste).
La synapse chimique
La synapse chimique est la plus fréquente des synapses du système nerveux. Ce type de synapses transmet le signal nerveux d'un neurone à un autre en utilisant un neurotransmetteur qui est émis par le neurone afférent, diffuse dans la fente synaptique et se lie aux récepteurs postsynaptiques.
Un dysfonctionnement de certains type de synapses est à l'origine de beaucoup de troubles nerveux, tels que l'épilepsie ou le trouble déficitaire de l'attention traité avec le méthylphénidate mieux connu sous l'appellation de Ritalin®, inhibiteur de recapture présynaptique de la dopamine.
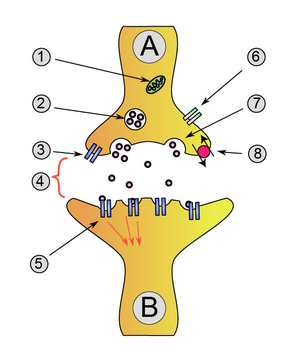

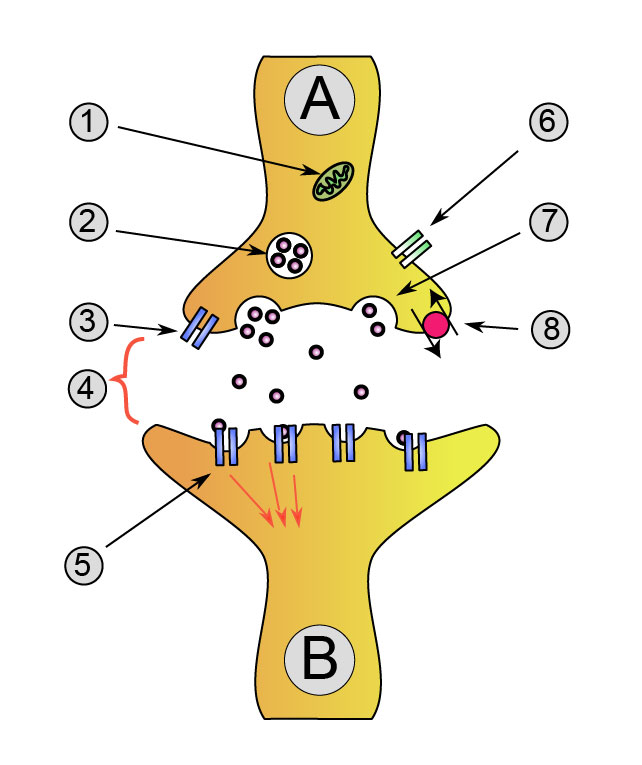
Transmission chimique du neurone A (émetteur) au neurone B (récepteur)
- Mitochondrie
- Vésicule synaptique avec des neurotransmetteurs
- Autorécepteur
- Fente synaptique avec neurotransmetteur libéré (ex : sérotonine ou dopamine)
- Récepteurs postsynaptiques activés par neurotransmetteur (induction d'un potentiel postsynaptique)
- Canal calcium
- Exocytose d'une vésicule
- Neurotransmetteur recapturé
Nombreux contacts synaptiques
Dans certaines synapses les cellules gliales jouent un rôle particulièrement actif, notamment en effectuant la recapture du neurotransmetteur ( ex : glutamate). Dans les synapses motrices, une enzyme, la cholinestérase, dégrade le neurotransmetteur dans la fente synaptique.
Morphologie
Il existe deux morphologies de synapses chimiques : la synapse en bouton et la synapse « en passant ». Elles fonctionnent toutes les deux de la même façon et l'on y retrouve les mêmes composants. La synapse en bouton se situe à l'extrémité de la fibre nerveuse alors que les synapses en passant sont réparties régulièrement le long de l'axone.
Il existe un grand « bestiaire » de synapses chimiques selon le type de neurone, la localisation, etc. Le calice de Held dans le tronc cérébral auditif, par exemple, est un cas de synapse géante qui entoure quasiment complètement la cellule postsynaptique.
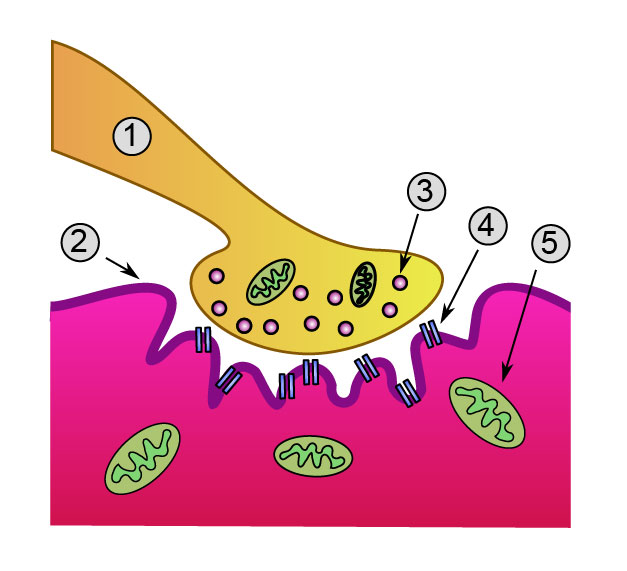
La synapse est constituée de trois parties : l'élément présynaptique, l'élément postsynaptique et entre les deux l'espace intersynaptique.
- L'élément présynaptique se présente sous la forme d'un renflement de l'axone, rempli de petites vésicules de formes variées (les vésicules synaptiques) contenant le neurotransmetteur. On y trouve aussi un appareil de Golgi très développé et de nombreuses mitochondries, signe d'une activité de synthèse intense. Les neurotransmetteurs sont en effet en partie synthétisés sur le lieu d'utilisation.
- L'élément postsynaptique en revanche est totalement dépourvu de vésicule. Mais il contient quelques mitochondries, nécessaires pour assurer le fonctionnement de la synapse. Dans certains cas la membrane y est plus épaisse, ce qui permet de caractériser des synapses asymétriques.
- L'espace intersynaptique (ou fente synaptique) est la zone qui sépare les membranes des deux neurones. Elle est de petite dimension (quelques dizaines de nanomètres) et dépourvue de lame basale (contrairement à la plaque motrice).
Transmission de l'influx nerveux
L'influx nerveux est transmis le long d'un neurone sous la forme d'une séquence de potentiel d'action. Au niveau d'une synapse chimique, l'information change de nature : elle est transmise par une libération de neurotransmetteurs dans l'espace synaptique. Les trains d'onde de dépolarisation supportés par des courants électrochimiques (les potentiels d'action), sont convertis en codage par concentration de neurotransmetteur dans la fente synaptique.
Pendant longtemps, le credo a fait force de loi : un neurone, un neurotransmetteur. Aujourd'hui, on sait qu'un neurone peut libérer plusieurs neurotransmetteurs au niveau de la synapse, en général un transmetteur principal associé à un ou plusieurs neuropeptides. Le transmetteur principal peut même évoluer, comme par exemple certains neurones orthosympathiques (noradrénergiques), qui peuvent libérer de la sérotonine suite à une lésion.
Evénements présynaptiques : la libération des neurotransmetteurs
Il faut d'emblée différencier les neurotransmetteurs peptidiques et non peptidiques. Les neurotransmetteurs peptidiques sont produits par le neurone à partir d'acides aminés précurseurs présents dans le sang. Une grande partie de leur synthèse a lieu dans le péricaryon, en suivant le schéma classique de toute production protéique (Transcription de l'ADN en ARNm, lecture et traduction de l'ARNm par un ribosome sur le réticulum endoplasmique) puis transport antérograde rapide le long du cytosquelette de l'axone dans des vésicules provenant du bourgeonnement de l'appareil de Golgi. Une étape de maturation a lieu dans les vésicules golgiennes (clivages des extrémités N-ter et C-ter par des exopeptidases, clivage dans le peptide par des endopeptidases, amidation sur des acides aminés Glycine, acétylation...). Les vésicules sont ensuite accumulées près de l'extrémité présynaptique, dans l'attente d'une dépolarisation.
Les neurotransmetteurs non peptidiques sont produits à partir d'acides aminés (Catécholamines comme l'adrénaline ou la noradrénaline à partir de la tyrosine, le GABA (Gamma AminoButyric Acid),...), de lipides (THC pour TetraHydroCannabinol)... Ils sont produits dans le cytoplasme du neurone ou de la cellule excitable et sont activement pompés (par des enzymes utilisant l'ATP ou ATPases) dans des vésicules issues des endosomes ou d'une endocytose.
Le changement de polarité de membrane provoqué par l'arrivée d'un potentiel d'action (PA) au niveau d'une synapse déclenche l'ouverture de canaux calcium membranaires dépendants du voltage (VOC = Voltage Operated Channels). L'augmentation de la concentration en calcium intracellulaire qui en résulte provoque la fusion de la membrane vésiculaire avec la membrane plasmique et la libération des neuromédiateurs. Ce phénomène s'appelle l'exocytose. La biologie cellulaire a montré que cette exocytose était assurée par un complexe appelé SNARE composé principalement de 3 protéines :
- VAMP (aussi appelée synaptobrévine), insérée dans la membrane plasmique de la vésicule
- la syntaxine arrimée à la membrane plasmique dde la cellule
- SNAP 25 arrimée dans la membrane plasmique
Lors d'une dépolarisation ouvrant des VOC au calcium (VOC Ca++), une brusque entrée de calcium précipite la fusion de VAMP avec SNAP 25 et la syntaxine, ce qui arrime la vésicule à la membrane plasmique. La modification tridimensionelle de ce complexe ternaire conduit à la fusion de la vésicule avec la membrane et à la libération du neurotransmetteur dans la fente synaptique.
Trois mécanismes peuvent arrêter l'exocytose et donc faire cesser la libération de neurotransmetteur dans la fente synaptique :
- l'ouverture de canaux potassium, qui ramènent le potentiel de membrane à sa valeur d'origine et inhibent ainsi les canaux dépendants du voltage;
- des pompes calciques, situées sur le réticulum et la mitochondrie, qui captent les ions calcium entrés dans la cellule, ce qui fait cesser le signal calcique ;
- disparition des vésicules synaptiques chargées en neurotransmetteur capable de fusionner avec la membrane (épuisement).
Ces trois mécanismes expliquent l'existence de plasticités synaptiques à plus ou moins long terme.
La diffusion des neurotransmetteurs dans la fente synaptique
Les neurotransmetteurs libérés dans la fente synaptique atteignent la membrane postsynaptique par simple diffusion. Avec le délai nécessaire pour provoquer l'exocytose, c'est l'étape qui nécessite le plus de temps dans la transmission synaptique. Dans le cas de la plaque motrice, la concentration en acétylcholine dans la fente atteint une concentration de 100 mmol/l 10 µs après sa libération. Elle mettra environ 100 µs pour revenir à une concentration proche de zéro. Cette disparition du neurotransmetteur de la fente synaptique peut impliquer un recaptage ou une hydrolyse par une enzyme spécialisée. Le codage de l'information étant fréquentiel, il est important de faire cesser l'excitation le plus vite possible.
Les événements postsynaptiques : l'activation des récepteurs membranaires
Les neurotransmetteurs se fixent sur des récepteurs de la membrane postsynaptique. Il en existe deux sortes :
- les récepteurs ionotropes qui sont des protéines-canal s'ouvrant pour générer un courant ionique ;
- les récepteurs métabotropes qui sont des transducteurs de signal produisant des seconds messagers dans le cytoplasme. Les seconds messagers peuvent s'associer à une protéine-canal ou bien provoquer une cascade de réactions. Parmi les voies métaboliques activées par ces seconds messagers, des facteurs de traduction de l'ADN sont impliqués, ce qui influence le pool de gènes exprimé par la cellule, et donc pourrait être impliqué dans le phénomène de mémorisation. Cette voie est beaucoup plus lente que la première.
On assiste alors à une réponse physiologique locale appelée potentiel générateur, potentiel gradué (PG) ou Potentiel postsynaptique. On caractérise deux types de potentiel postsynaptique :
- Le Potentiel Postsynaptique Excitateur (ou PPSE) diminue la différence de potentiel entre les deux côtés de la membrane plasmique. Autrement dit le PPSE dépolarise localement la membrane ;
- Le Potentiel Postsynaptique Inhibiteur (ou PPSI) augmente la différence de potentiel. Elle hyperpolarise la membrane.
Si la membrane dépasse le seuil critique de dépolarisation, un potentiel d'action est initié. Les PPSI empêchent le déclenchement d'un potentiel d'action alors que les PPSE le favorisent.
En général, un neurone est couvert de synapses excitatrices et de synapses inhibitrices. Il se produit alors une sommation à la fois temporelle et spatiale des entrées synaptiques pour "décider" du déclenchement ou non d'un potentiel d'action. En fait les dendrites ont peu de canaux sodiques dépendants du voltage, responsables du déclenchement du potentiel d'action. Il est donc rare qu'un potentiel d'action y soit déclenché. Les potentiels postsynaptiques se propagent le long des dendrites jusqu'au péricaryon. À la jonction du péricaryon et de l'axone se trouve une région particulièrement riche en canaux sodiques dépendants du voltage, il s'agit du cône d'initiation. C'est au niveau du cône d'initiation que sont générés le plus souvent les potentiels d'actions qui se propageront ensuite le long de l'axone vers d'autres synapses...
Le potentiel d'action, une fois initié, a toujours la même amplitude et le même décourt temporel. Sa valeur informative ne dépend pas de l'importance de la dépolarisation qui l'a initié. C'est cela qu'on appelle la loi du tout ou rien. Si la dépolarisation continue suffisamment longtemps après le déclenchement du potentiel d'action, un autre potentiel d'action peut être initié. Les potentiels d'action codent l'information en fréquence.
Plusieurs molécules étant libérées lors de la transmission synaptique et plusieurs types de récepteurs pour le même neurotransmetteur pouvant être présents sur la même membrane postsynaptique, plusieurs effets peuvent avoir lieu simultanément. C'est par exemple le cas de nombreuses synapses GABAergiques qui présentent un PPSI rapide dû aux récepteurs ionotropes GABAA et un PPSI lent dû aux récepteurs métabotropes GABAB.
L'arrêt de la stimulation nerveuse
Pour éviter que la stimulation du neurone postsynaptique ne se prolonge, deux systèmes éliminent la molécule de l'espace intersynaptique :
- La dégradation, qui met en jeu des enzymes spécifiques qui vont métaboliser le neurotransmetteur, mettant fin à son effet sur le neurone postsynaptique exemple la MAO issue des synthèses mitochondriales ;
- La recapture, pendant laquelle le neurotransmetteur ou ses précurseurs issus de la dégradation enzymatique est récupéré par le neurone présynaptique, ou par la cellule gliale avoisinante, pour être réutilisé ou détruit.
En général les deux sont associés. Dans le cas de l'acétylcholine, une dégradation limitée est suivie d'une recapture de la choline qui sera utilisée pour resynthétiser l'acétylcholine.
Influence des substances psychoactives
Les substances psychoactives sont des drogues, des médicaments,... qui modifient les perceptions sensorielles, les sensations, l'humeur ... Elles ont comme principal mode d'action de modifier le passage des neurotransmetteurs.
Comme pour les neurotransmetteurs, il existe plusieurs modes d'action possibles à ces drogues, dont :
- Se lier aux récepteurs sans entrainer d'effet (effet antagoniste). Les récepteurs ne sont alors plus disponibles pour lier l'agoniste (neurotransmetteur) ;
- Empêcher ou limiter la sortie ou la destruction de neurotransmetteur, qui active davantage et plus longtemps le récepteur (exemple du Prozac®).
Les conséquences à long terme sont de modifier la réceptivité de la synapse, par exemple en modifiant le nombre de récepteur, en réaction de défense, ce qui entraine l'accoutumance et la dépendance.
La jonction neuromusculaire
Lors du réflexe myotatique CF réflexe de flexion, l'élément présynaptique rencontre la plaque motrice de la fibre musculaire qui est composée d'une membrane plasmique appelée sarcolemme faisant office d'élément postsynaptique et contenant plusieurs centaines de myofibrilles. La jonction neuromusculaire est historiquement très importante puisque ce sont les observations sur le muscle extenseur du muscle de patte de grenouille qui ont donné naissance à l'électrobiologie qui eut un retentissement rapide auprès du grand public, comme en témoigne l'engouement de l'époque pour les phénomènes électriques. On se rappellera que le monstre du Dr Frankenstein est animé par l'électricité.
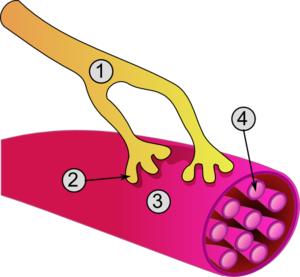
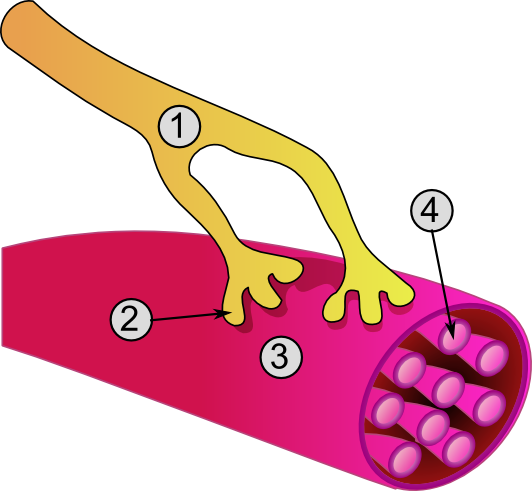
- Axone
- Jonction
- Fibre musculaire
- Myofibrille
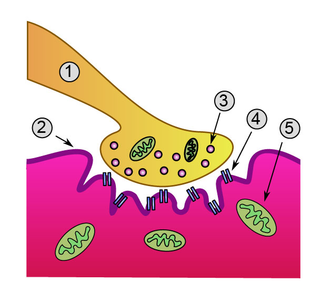
Vue détaillée d'une jonction. - Élément pre-synaptique
- Sarcoplasme
- Vésicules synaptiques
- Récepteur cholinergique nicotinique
- Mitochondrie
L'acétylcholine intervient dans la contraction musculaire lors des réflexes de flexion ou d'extension au niveau de la jonction neuromusculaire. Les neurones la produisant s'appellent neurones cholinergiques. Ses précurseurs sont la choline d'origine alimentaire qui est captée par la terminaison présynaptique dans le sang et l'acétylcoenzyme A d'origine mitochondriale. Ils sont synthétisés par l'enzyme choline-acétyltransférase (CAT) qui les transforment en acétylcholine. Ces neuromédiateurs sont alors enveloppés par des vésicules provenant du bourgeonnement de l'appareil de Golgi et sont transportés jusqu'au renflement (ou bouton) synaptique. Au niveau présynaptique il y a non pas un seul renflement mais des centaines afin d'assurer une surface de contact plus large, on parle d'arborisation terminale.
Sous l'effet du calcium, les vésicules chargés de neuromédiateurs fusionnent avec la membrane plasmique, déversant leur contenu dans la fente synaptique (exocytose). Les neuromédiateurs se fixent alors sur des récepteurs spécifiques de la plaque motrice du muscle strié ce qui a pour conséquence de provoquer sa contraction. L'excès de neuromédiateur est ensuite dégradé par une enzyme : acétylcholinestérase (ACHE) qui libère de l'acide acétique et de la choline qui pourra être ensuite recapturée par les récepteurs de l'axone présynaptique et recyclé.
La synapse électrique
Dans la synapse électrique, les membranes des deux neurones sont reliées par des jonctions communicantes, parfois appelées également nexus (GAP junctions). Les ions se transmettent donc d'une cellule à une autre, ainsi que la dépolarisation membranaire associée. L'influx nerveux se transmet sans intervention de neurotransmetteur. Ce type de synapse, qui joue un rôle important dans le système nerveux immature, est ensuite relativement rare au stade adulte et est majoritairement retrouvé chez les invertébrés. Ce type de communication est très fréquent dans les épithéliums.
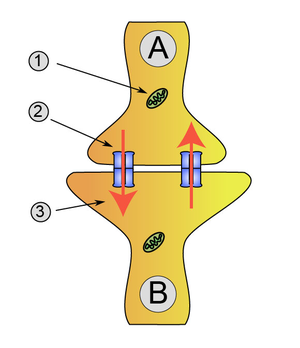
Transmission électrique du neurone A (émetteur) au neurone B (récepteur)
- Mitochondrie
- Connexine
- courant ionique
Les caractéristiques principales de ce type de synapse sont :
- un délai de transmission quasi-inexistant (pas de temps de latence dû au franchissement d'une synapse)
- une conduction dans les 3 directions de l'espace
- l'absence de période réfractaire (la synapse est restimulable immédiatement après la fin de la transmission)
Intégration du signal : zoom sur l'élément postsynaptique
Les synapses sont regroupées selon deux catégories selon les effets qu'elles engendrent : excitatrices ou inhibitrices. Le principal neuromédiateur inhibiteur du cerveau est le GABA qui se fixe sur les canaux récepteurs GABAA dont l'ouverture provoque un influx d'ions chlorure et donc une hyperpolarisation de la membrane. Il existe une plus grande diversité de récepteurs ionotropes excitateurs, par exemple les récepteurs au glutamate ou à l’acétylcholine. L'élément postsynaptique possède en général ces deux catégories de récepteurs ainsi que des canaux sodium ou calcium activés par dépolarisation. Il réalise une sommation temporelle des signaux excitateurs (PPSE, potentiel postsynaptique excitateur) et inhibiteurs (PPSI, potentiel postsynaptique inhibiteur). Il propagera le potentiel d'action à la condition que la somme des excitations soit supérieure à la somme des inhibitions et si un seuil de dépolarisation est atteint. Ce seuil correspond au voltage auquel un nombre suffisant de canaux sodiums sont activés.
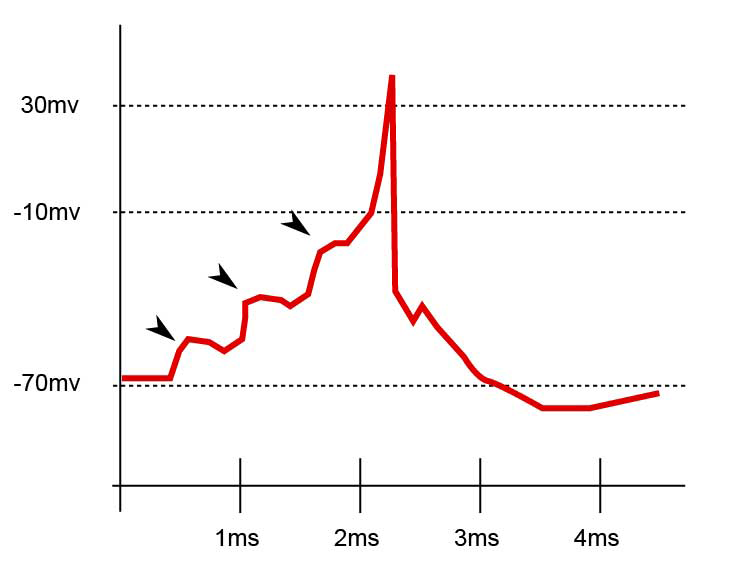
 Enregistrement postsynaptique du potentiel membranaire. Les flèches marquent les PPSE de trois évènements afférents. Une sommation de trois PPSE donne naissance dans ce cas au déclenchement du potentiel d'action.
Enregistrement postsynaptique du potentiel membranaire. Les flèches marquent les PPSE de trois évènements afférents. Une sommation de trois PPSE donne naissance dans ce cas au déclenchement du potentiel d'action.La sommation spatiale se réfère aux différentes synapses afférentes à l'élément postsynaptique. Un neurone peut en effet recevoir plus d'un millier d'afférences différentes mais il ne peut réagir que d'une seule manière : conduction ou absence de conduction. Si le résultat de la somme algébrique de tous les éléments afférents est supérieure à une valeur seuil, aux environs de -15 mV dans le schéma ci-contre, le neurone intégrateur sera le siège d’un potentiel d’action.
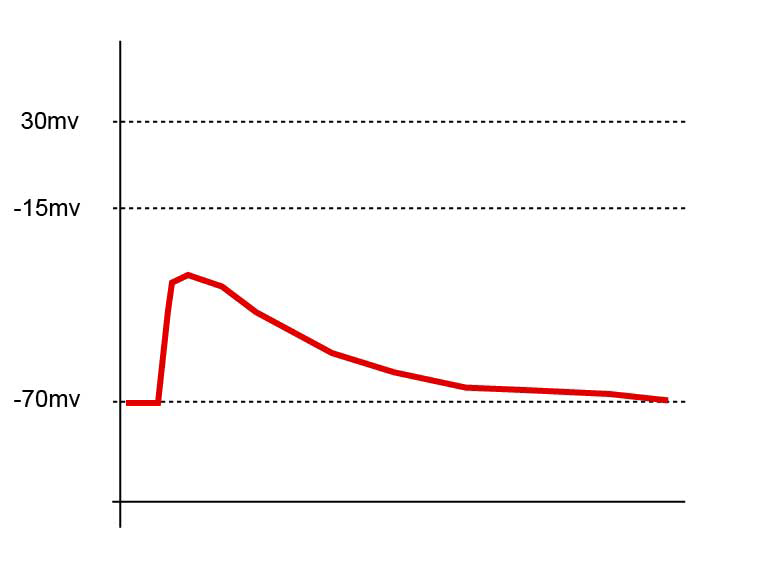
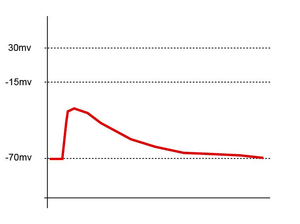 Dépolarisation subliminale un seul PPSE ne dépolarise pas suffisamment la membrane pour générer un potentiel d’action.
Dépolarisation subliminale un seul PPSE ne dépolarise pas suffisamment la membrane pour générer un potentiel d’action.Une sommation dite temporelle a aussi lieu au niveau de l’élément postsynaptique. Elle est due à la vitesse d'entrée des ions à l'intérieur de la cellule. Si beaucoup de PPSE sont rapprochés dans le temps, ils s'ajoutent et peuvent également atteindre le seuil de dépolarisation et donner lieu à un potentiel d’action.
Un dernier élément d’intégration est dû à l’existence de la période réfractaire du neurone. Si deux signaux afférents excitateurs sont espacés de moins d'une milliseconde, le second ne donnera naissance à aucun PPSE et sera donc silencieux.
Pathologie
L'anomalie de fonctionnement de la synapse neuro-musculaire est responsable d'une maladie neuromusculaire nommée myasthénie.
Articles connexes
- Neurone
- Cellule gliale
- synapse et transmission synaptique
- neurotransmetteur
- plasticité synaptique
- potentiel postsynaptique
- réflexe d'extension
- réflexe de flexion
- cerveau
- moelle épinière
- Classement thématique des neurosciences
- Histoire de l'électrophysiologie
- Électrophysiologie
- Potentiel de repos
- Biophysique des canaux ioniques
- Cellules nerveuses
Catégories : Neuroscience | Neurophysiologie


Wikimedia Foundation. 2010.