- Potentiel de repos
-
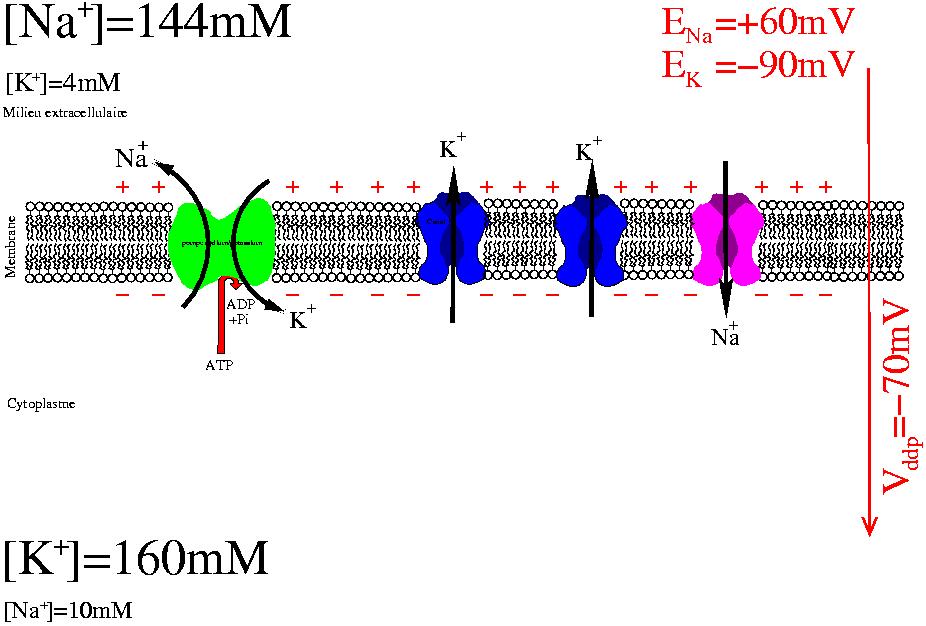
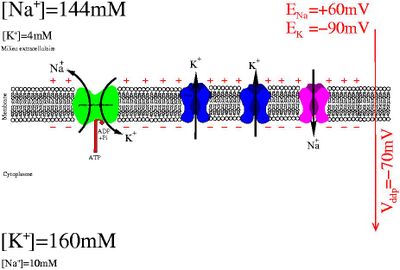 Le potentiel de membrane d'une cellule est dû à la séparation de charges consécutive au flux ionique à travers les canaux potassium, lui même dû au déséquilibre ionique entretenu activement par les pompes sodium/potassium
Le potentiel de membrane d'une cellule est dû à la séparation de charges consécutive au flux ionique à travers les canaux potassium, lui même dû au déséquilibre ionique entretenu activement par les pompes sodium/potassium
Le potentiel de repos (soit un des états possible du potentiel de la membrane) est la polarisation électrique en situation physiologique de repos d'une membrane plasmique. En introduisant une électrode de mesure à l'intérieur de la cellule (voir la méthode de patch-clamp), on constate une différence de potentiel (ddp): la face interne de la membrane est négatif par rapport à une électrode de référence placée sur la face externe de la membrane.
Cette différence de potentiel est due à la séparation de charge de part et d'autre de la membrane provoquée par un courant permanent majoritairement d'ion potassium à travers des canaux ioniques. Ce courant dissipe la force électroosmotique causée par des différences de concentration entre différentes espèces ioniques. Cette différence de concentration est maintenue en permanence par l'activité consommatrice en énergie des pompes sodium-potassium.
L'existence d'un potentiel de membrane est universelle aux cellules vivantes.
Sommaire
Mécanismes
Facteurs biologiques
Les propriétés actives de la membrane plasmique sont à l'origine de cette différence de potentiel (ddp):
- Propriétés propres à la bicouche phospholipidique:
- imperméabilité qui permet de maintenir les différences de concentration de part et d'autre de la membrane, et donc un gradient chimique (voir le tableau ci-dessous)
- haute résistance électrique qui permet de maintenir une différence de potentiel, donc un gradient électrique
- Propriétés propres aux protéines membranaires
- les canaux potassium de fuite sont responsable d'une perméabilité sélective de la membrane ne laissant passer que les ions potassium
- les pompes potassium/sodium transportent activement les ions sodium vers l'extérieur et les ions potassium vers l'intérieur.
Les concentrations physiologiques des principaux ions chez l'homme Ion concentration intracellulaire (mmol/l) concentration extracellulaire (mmol/l) Rapport Potentiel d'équilibre d'après l'équation de Nernst Na+ 7-12 144 1:12 env. + 60 mV K+ 160 4 40:1 - 91 mV Ca2+ 10-5-10-4 2 + 125 mV à + 310 mV Mg2+ 0.1-3 1 Cl- 4-7 120 1:22 env. - 82 mV HCO3- 8-10 26-28 1:3 - 27 mV Protéine anionique (chargée négativement) 155 5 Mécanismes physiques
Le modèle pompe/fuite (pump/leak) repose sur les effets antagonistes de la pompe sodium/potassium (Na/K ATPase) d'une part et des canaux potassium d'autre part.
La Na/K ATPase utilise l'énergie contenue dans l'ATP pour maintenir une différence de composition ionique entre l'intérieur de la cellule et l'extérieur. L'activité de la pompe a pour effet direct que les ions potassium sont majoritaires dans le cytoplasme de la cellule, tandis que les ions sodium sont majoritaires à l'extérieur de la cellule. L'ouverture de canaux potassiums, les seuls canaux qui soient ouverts à l'état basal dans la majorité des cellules, permet au gradient chimique du potassium de se dissiper. La séparation de charge résultante crée la différence de potentiel électrique mesurée. L'électroneutralité des deux compartiments est violée à proximité de la membrane. Toutefois, compte tenu de la géométrie du système, il ne faut qu'un surplus d'environ 2 ions sur 100 000 pour rendre compte du potentiel de membrane. L'électroneutralité est bien respectée macroscopiquement. Le champ électrique créé empêche les ions potassiums de sortir.
Pour résumer, le potentiel chimique des ions potassiums est en faveur d'une sortie de ces ions. Cette sortie crée une force électrique qui s'oppose à la sortie d'un nombre plus important d'ions potassium.
Dans les neurones et autres cellules excitables, un signal provoque l'ouverture transitoire des canaux sodium responsables d'une dépolarisation transitoire appelée potentiel d'action. Le potentiel de Nernst des ions sodiums est de l'ordre de + 60 mV. Également, la spécificité des potentiels d'actions est conférée par la grande diversité des canaux ioniques impliqués selon la cellule considérée.
Calcul de la séparation de charge
L'application numérique qui suit a pour but de démontrer que l'électroneutralité de la solution est respectée, malgré l'existence d'une différence de potentiel membranaire.
Soit une cellule de 20 µm de diamètre ayant une concentration de 160 mM en ions K+.
La membrane est un isolant séparant deux milieux conducteurs. Elle est donc un condensateur. La charge d'une condensateur plan est donné par :
Q = C.V
où- C est la capacité en farad (F)
- V la tension électrique (en anglais, voltage) en volt (V)
- Q la charge en coulomb (C).
Note: Rappelons qu'un coulomb est l'unité SI utilisée pour représenter une quantité de charges électriques. Un coulomb équivaut à 1.6x1019 charges électriques élémentaires.
Calculons la capacité Cm de la membrane sachant qu'il vaut 1 µF.cm-2 soit 0.01 pF.µm-2.
Il faut calculer la surface de la cellule qui est donné en première approximation par celle d'une sphère :
 µm2 (où d est le diamètre)
µm2 (où d est le diamètre)Ce qui correspond à une capacité de Cm=12 pF. La charge Q accumulée sur les deux faces de la membrane donnant naissance à un champ électrique de - 70 mV est de :
Q = 12.10-12 * 0.07 = 8.4.10-13 C,
soit :
 ions.
ions.
Il faut à présent comparer ce nombre au nombre total d'ion K+ contenu dans la cellule.Le volume de la cellule est approximativement celui d'une sphère :
 soit 4.10 − 12 litre. Il y a donc 0.16 * 4.10 − 12 * 6.02.1023 = 3.8.1011 ions K+ dans la cellule.
soit 4.10 − 12 litre. Il y a donc 0.16 * 4.10 − 12 * 6.02.1023 = 3.8.1011 ions K+ dans la cellule.Ainsi, 1.4 ion sur 100 000 est responsable de la différence de potentiel (ddp) mesurée. Cet excès de charge est localisé à la membrane (voir figure). L'électroneutralité de la solution est donc violée à la proximité de la membrane mais est respectée partout ailleurs.
Notons que l'intensité du champ électrique se dissipant à travers la membrane d'épaisseur 75 nm est de 1.5 million de volts par mètre, ce qui est considérable et est à la limite de la fracture de la membrane.
Rôles physiologiques
- Toute cellule vivante a un potentiel de membrane. Le potentiel électrochimique qui lui correspond constitue une réserve d'énergie potentielle qui permet d'assurer les transports de substances nécessaires à la survie de la cellule.
- Le potentiel de repos joue un rôle important pour les cellules excitables, comme les neurones et les myocytes. En effet le franchissement par le potentiel de repos d'un certain seuil de dépolarisation déclenche chez ces cellules un potentiel d'action par l'activation de canaux dépendent du potentiel (canaux voltage-dépendant). L'importance de la polarisation du potentiel de repos détermine donc l'excitabilité de la cellule. Quand il est très hyperpolarisé (par l'activation tonique de canaux potassiques ou chlorures par exemple), la cellule est difficilement excitable, c'est-à-dire qu'il faut beaucoup dépolariser la cellule avec des potentiels postsynaptiques excitateurs pour qu'elle décharge un potentiel d'action. Quand le potentiel de repos est très dépolarisé (par la fermeture de canaux potassium ou par l'ouverture permanente de canaux sodium), la cellule est plus proche du seuil de déclenchement d'un potentiel d'action, et donc plus excitable.
Articles connexes
- Propriétés propres à la bicouche phospholipidique:
Wikimedia Foundation. 2010.