- Membrane plasmique
-
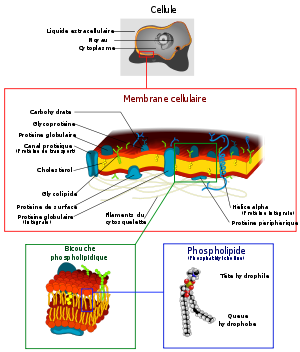 La membrane plasmique d'une cellule eucaryote.
La membrane plasmique d'une cellule eucaryote.
La membrane plasmique, ou plasmalemme, est la membrane qui délimite une cellule, elle sépare le cytoplasme du milieu extérieur.
Sommaire
Description
 Structure de la bicouche lipidique.
Structure de la bicouche lipidique.La membrane plasmique forme une pellicule continue de 6 à 9 nm d’épaisseur (en moyenne 7) délimitant le cytoplasme du milieu extracellulaire. Ce micro-écotone est composée de phospholipides, de protéines (intracellulaires, extracellulaires ou insérées dans la double couche phospholipidique) et de molécules de cholestérol.
Grâce à une perméabilité très sélective, elle joue un double rôle de protection et de contrôle des échanges entre les milieux intracellulaire et extracellulaire.
Elle est à distinguer des endomembranes qui délimitent les organites.Composition
Deux feuillets sont accolés ;
- l'hémimembrane E (« E » pour exoplasmique) ; C'est la couche externe de la membrane.
Ce feuillet est parfois lui-même recouvert d'une structure finement fibrillaire dite glycocalix ou cell-coat.
Sur cette face on trouve aussi fréquemment des glucides ou dérivés de glucides (sous forme d'acide sialique par exemple), dont toutes les fonctions ne sont pas encore comprises ; - l'hémi-membrane P, (« P » pour protoplasmique) côté interne, éventuellement plus ou moins liée à d'autres membranes intra-cellulaires.
Ces feuillets sont formés de phospholipides et de glycolipides (uniquement du côté extracellulaire) qui se font face par leur pôle hydrophobe.
Entre les molécules phospholipides se trouvent des molécules dites intrinsèques et du cholestérol (uniquement chez les eucaryotes et dans le monde animal, des phytostérols jouant un peu le même rôle chez les végétaux).
Sur les faces externe et interne, constituées par les pôles hydrophiles des phospholipides membranaires, se fixent des protéines extrinsèques sur lesquelles s’accroche le cytosquelette ou les molécules de la substance (matrice) extracellulaire.- Cet assemblage non-covalent de lipides et de protéines, forme une structure souple permettant à presque tous ses éléments - dans une certaine mesure - de se déplacer latéralement. Cette « fluidité membranaire » sera d'autant plus importante que les acides gras constituant les lipides membranaires sont courts et insaturés.
- Des protéines spéciales assurent la perméabilité sélective de la membrane (pores), la reconnaissance de signaux chimiques ou portent des marqueurs (antigènes du CMH…).
Les pores sont les portes de la membranes. Ce sont des édifices macromoléculaires de nature protéique ou glycoprotéique, parfois très complexe. Ils sont insérés dans la bicouche et contrôlent les échanges entre milieu intra et extra cellulaire (avec des récepteurs, transporteurs).La membrane est asymétrique et parfois irrégulière, en raison de
- différences de présence de lipides entre ses deux feuillets,
- différence de nature et composition en sucres,
- différence dans le mode d’insertion des protéines,
- différence de composition biochimique (d'un type cellulaire à l’autre, d'un endroit de la membrane à un autre ; aux niveaux apical ou basal par exemple). Ces différences évoluent parfois rapidement dans le temps.
Lipides
Ce sont des constituants essentiels du « bi-couches » membranaire (environ 50 % du poids sec de la membrane) dont la cohérence est assurée par les oppositions de pôle hydrophile et hydrophobe (les lipides sont amphipathiques).
Deux types lipidiques sont plus abondants :
- le cholestérol ; bien que presque absent des membranes des compartiments intracellulaires, il est essentiel dans la membrane plasmique ; Avec 15 à 20 % du poids sec de la membrane (et jusqu'à 30 % dans la membrane des érythrocytes), il y est en moyenne numériquement autant présent que les lipides (en nombre de molécules) ; Ce cholestérol forme un film étanche supplémentaire disposé entre les lipides membranaires, qu'il consolide sans la rigidifier. Il bloque toutes les molécules hydrophiles qui pourraient accidentellement s'insinuer dans le film lipidique moins cohérent ;
- les phospholipides, répartis en deux familles :
- phosphoglycérides. Au nombre de trois (chez les mammifères), ces dérivés du glycérol sont
-
- - Phospatidyléthanolamine,
- - P.choline
- - P.sérine (P.sérine = P. lipide chargé négativement)
-
- la sphingomyéline (2;l*m0%), molécule polaire, dérivée de la sphingosine, avec une chaîne d'acides gras liée par un phosphate. Cette molécule contient de la choline, isolant électrique, composant principal de la gaine de myéline des axones.
- d’autres phospholipides, moins abondants, mais essentiels pour les transmission de signaux, dont
- - des dérivés de l'inositol, qui sont :
- - des Glycosyl-Phosphatidyl-Inositol (GPI ou glycosil phosphatidyle inositol), qu'on ne trouve que dans le feuillet externe. Ce GPI permet l' ancrage de certaines glycoprotéines au feuillet externe de la membrane plasmique ;
- - divers autres dérivés présents sur le feuillet interne de la membrane (rôle majeur dans la transmission du signal)
La partie polaire : Elle forme un film souple composée d'un plan de phosphate estérifié par une molécule polaire (éthanolamine, choline, sérine) et relié à une molécule hydrophobe (sphingosine : alcool aminé) ou à un phosphoglycéride (en l'occurrence du glycérol, qui est également une molécule polaire).
La partie hydrophobe : C'est un film d'acides gras (AG), disposés de telle sorte qu'ils ne forment pas de motif de type cristallin, ce qui garantit à la membrane un bon compromis entre cohérence, résistance (forces de Van der Waals) et souplesse, avec des propriétés électriques intéressantes, la membrane isolant en quelque sorte la cellule et ses organites.
Protéines
- Elles représentent l'autre moitié du poids sec de la membrane, mais occupent un volume plus important que les lipides qui sont plus petits que les protéines (50 molécules de lipides = 1 molécule de protéine en masse).
- Le type de protéine varie selon le type de cellule et ses fonctions. De même pour leur quantité : très abondantes dans la membrane interne de la mitochondrie ou dans la membrane du réticulum endoplasmique (70 % de la masse membranaire), abondante dans la membrane des hématies, hépatocytes ou des érythrocytes (50 % de la masse membranaire), elles sont plus éparses dans la membrane des cellules nerveuses (qui doivent être électriquement isolées, d'où l'abondance (20 % du poids sec de la membrane) de myéline).
- Elles sont divisées en deux classes :
- - protéines dites « extrinsèques » (ou périphériques) ; Elles sont insérées dans la bicouche lipidique, mais par des liaisons de faible énergie (Force de van der Waals) ; une simple variation de pH ou des conditions ioniques suffit à les en détacher ;
- - protéines dites « intrinsèques » ; transmembranaires ou solidement greffées par une liaison covalente à l'un ou l'autre des feuillets de la bicouche (à l’aide d’un acide gras ou par un GPI) ; Elles ne se détachent pas de la membrane ; le seul moyen de les extraire est de détruire la bicouche lipidique membranaire en utilisant un détergent puissant.
Glucides
Les sucres fixés sont des polysaccharides ou des oligosaccharides.
Ils sont toujours liés soit à des protéines (avec lesquels ils forment des glycoprotéines), soit à des lipides en formant alors des glycolipides. Les sucres les plus abondants sont des glycolipides fixés aux sphingolipides (et donc insérés dans les radeaux lipidiques).
Ils sont tous localisés à la face extracellulaire de la membrane, alors qu'à l'intérieur de la cellule où ils sont synthétisés, quand ils sont fixés, ils le sont sur les parois internes des cavités endomembranaires (organites à membranes).Architecture fonctionnelle de la membrane plasmique
Bicouche lipidique
La bicouche enveloppe les protéines intrinsèques. Par exemple : on solubilise les lipides des globules rouges, on récupère les lipides de la membrane, on les met en suspension, on peut ainsi calculer la surface de la membrane plasmique de cet ensemble de cellules. Cela démontre l’organisation de ces lipides en deux couches. Organisation asymétrique : les glycolysations ont toujours lieu sur la face extracellulaire. Le cytosquelette : vers le cytoplasme. Les ponts disulfures sur les protéines, à part quelques exceptions, sont toujours à l’extérieur. Cette membrane est en perpétuel mouvement : les constituants de la membrane bougent. Les constituants se déplacent plus ou moins librement : propriétés de fluidité de la bicouche. Fluidité conditionnée par trois facteurs : la température (accélère les mouvements), la quantité de cholestérol (diminue la fluidité) et la nature des phospholipides. Les acides gras insaturés facilitent la fluidité, les acides gras saturés assurent la rigidité de la membrane. Trois types de mouvements : la diffusion latérale (un lipide peut changer de place avec son voisin), la rotation (sur place), le flip flop (changement de feuillet avec basculement, favorisé par des flipases avec consommation d'énergie sous forme d'ATP). Les protéines ne peuvent pas faire ce flip flop mais elles sont aussi capables de bouger dans cette membrane par rotation, diffusion latérale. Le capping (formation d’une petite coiffe), on marque la surface membranaire avec des anticorps fluorescents, les protéines sont marquées de façon diffuse, on observe rapidement des mouvements de ces protéines et des agrégats à certains endroits formant des coiffes à un pôle de la cellule. Au bout d’un certain temps la fluorescence disparaît et se concentre à l’intérieur du cytoplasme. Ce phénomène nécessite et consomme de l’énergie sous forme d'ATP. Ce phénomène peut être bloqué cinétiquement par le froid, ou artificiellement par des poisons métaboliques au niveau des mitochondries.
- Diffusion latérale des protéines limitant les mouvements du cytosquelette ;
- Interaction de la membrane plasmique avec la matrice extra-cellulaire ;
- Interaction avec des protéines membranaires ; c’est-à-dire qu'elles s’accrochent entre elles, ce qui peut limiter les déplacement membranaires.
Le fait que les protéines au niveau de la membrane soient accrochées aux protéines d’autres cellules va limiter les déplacements. Tous ces phénomènes régulent et limitent la diffusion des protéines de la membrane.
Composants chimiques
Les protéines trans-membranaires qui sont liées par un GPI peuvent être clivées par des enzymes appelées protéases. Ces protéases effectuent des coupures qui peuvent avoir lieu dans le milieu extracellulaire ou dicytosolique (grâce aux protéines cospases). Cela a été découvert lors de l’apoptose (mort programmée de la cellule. C’est ce qui fait que nous n’avons pas des mains en forme de palmes). Pour les cultures des cellules eucaryotes : Respecter les températures.
Échanges avec l’extérieur
Sur les membranes plasmiques des cellules, on a des lieux d’échange entre les deux milieux. On dit souvent que les cellules épithéliales sont polarisées : deux domaines particuliers : le pôle apical et le pôle basolatéral : distinction au niveau de la composition en protéines, etc.. Le pôle apical constitue une zone d’interactions entre les protéines membranaires et le cytosquelette, il y a notamment des structures appelées des microvillosités : extensions cytoplasmiques (de diamètre : 0,1 µm). Au sein de ces microvillosités, on a des microfilaments d’actine (un des trois composants du cytosquelette) associés à des protéines : échanges avec le milieu extracellulaire. Ex : les cellules épithéliales du système intestinal ont à leur surface des microvillosités permettant l'absorption des aliments. Zone apicale : lumière de l’intestin. La face basolatérale : face sur laquelle les cellules sont tournées vers le tissu conjonctif. Cette face sert au transfert vers les vaisseaux sanguins des aliments absorbés. On a des replis membranaires servant dans les échanges hydro-minéraux (cellules rénales, glandes salivaires, ..). On a aussi dans certaines cellules des cils (système respiratoire) : extensions cytoplasmiques : microtubules et protéines associées. Au niveau des bronches, c’est les battements de ces cils qui permettent l’évacuation des microbes sous forme de mucus.
Rôles principaux de la membrane plasmique
Il existe trois types de rôle :
Ces membranes servent à la communication intercellulaire via des signaux chimiques. Deux types de signaux chimiques sont produits. Première catégorie de signaux : ils sont captés et décodés par des récepteurs spécialisés dans les membranes. Ce sont de signaux hydrosolubles (peptidiques ou neurotransmetteurs). Le deuxième catégorie : signaux qui vont pénétrer dans la membrane plasmique : soluble dans la membrane plasmique lipide : liposolubles. Donc les messages sont de nature chimique soit par l’intermédiaire de récepteurs ou traversant la membrane.
Cette membrane plasmique sert à l’adhérence des cellules entre elles et dans le milieu (matrice extracellulaire) : jonctions intercellulaires, forme des domaines spécialisés d’adhérence. Ces domaines sont étroitement liés au cytosquelette.
Troisième grand rôle : transport de molécules au niveau de la membrane. Plusieurs types de transports existent : ceux sans mouvements particuliers : ces transports se déroulent à l’échelle moléculaire : trois caractéristiques majeures, le matériel transporté passe directement du milieu extra cellulaire au milieu intracellulaire ou inversement. Ce matériel transporté n’est jamais enfermé pendant son transport. Enfin, il n'y a jamais d’interventions du cytosquelette. Néanmoins, certains de ces transports peuvent nécessiter de l’énergie.
Transports avec mouvements : phénomènes d’endo ou d’exocytose. Ils impliquent une membrane en mouvement et impliquent le système endomembranaire : les molécules qui vont être transportées vont être pendant une partie de leur transport véhiculées par des vésicules ou des vacuoles. Cette membrane d’enveloppe peut provenir de la membrane plasmique (phagocytose) ou du compartiment du système membranaire (exocytose). Dans le cas de l’exocytose, le matériel qui va être exporté provient du cytosol : les molécules rentrent dans le système endomembranaire. Pour l’endocytose le matériel vient du milieu extracellulaire, traverse la membrane au niveau de structures appelées endosomes ou liposomes puis gagne le cytoplasme. La traversée du système endomembranaire fait intervenir des perméases. Ces mécanismes de transports nécessitent de l’énergie et l’intervention du cytosquelette : cortical.
En permanence la cellule contrôle ces échanges à travers la membrane. Ce contrôle va conduire à la différence de concentration de plusieurs petites molécules Ces transports nécessitent ou non de l’énergie et présence ou non d’une perméase (protéine ou complexe de plusieurs protéines membranaires directement responsables du transport). S'il y consommation d’énergie, on parle de transports actif, sinon passif. Certaines perméases possèdent une activité enzymatique qui va être utilisée pour fournir l’énergie nécessaire au transport. Certaines mitochondries, peroxysomes etc. possèdent aussi des perméases.
Transports
Transport passif sans perméases
Correspond à ce qu’on appelle la diffusion simple : sans consommation d’énergie. Aucune protéine membranaire n’intervient. Les molécules (ex : hormones lipophiles et liposolubles) sont prises dans la bicouche lipidique puis vont diffuser et repasser de l’autre côté de la membrane. Elle s’effectue selon le gradient de concentration (du milieu le plus concentré vers le moins concentré). Les particules qui diffusent à travers la membrane plasmique sous forme de diffusion simple sont les molécules liposolubles (ou hydrophobes) non polaires et les petites molécules polaires non chargées (comme l'eau, l'urée, etc.).
Transport passif avec perméases
On l'appelle la diffusion facilitée : transport passif, pas d'énergie avec perméases : nécessite une glycoprotéine transmembranaire : le transporteur passif. Canal ionique : complexe de protéines. Ce transport se déroule dans les deux sens selon le potentiel électrique établi à travers la membrane par les molécules chargées. Dans la diffusion facilitée, les molécules ne se dissolvent pas dans la bicouche, elles sont prises en charge par des protéines : permet de protéger les petites molécules de tout contact avec le cœur hydrophobe. La diffusion facilité permet aux molécules polaires (sucres, acides aminés, ions..) chargées de traverser cette membrane plasmique. Exemple d’un système facilité : pour le glucose, la perméase va alterner entre deux conformations possibles : le site de fixation du glucose est tourné vers l’extérieur faisant basculer le transporteur poussant le site de fixation vers l’intérieur de la cellule libérant tout le glucose. Cette diffusion facilitée va donc créer un flux continu de glucose comme le milieu extracellulaire est plus concentré que le milieu intracellulaire. Ce glucose disparaît vite car il est métabolisé rapidement. Ainsi la concentration intra n’augmente pas et on a une arrivée permanente de glucose dans la cellule.
Autre exemple : Les canaux ioniques dépendent de la concentration et de la charge des ions : reposent sur différentes propriétés.
- - Le passage est très rapide (1 million d’ions par seconde traversent un canal ouvert).
- - Ces canaux sont très sélectifs car leur pores sont très étroits.
- - Et ils sont intermittents. Il faut un signal (stimulus) pour l’ouverture. Il existe deux types de canaux selon le type d’ouverture, les canaux ioniques ligands-dépendants, fixation d’un ligand sur le canal (un neurotransmetteur par exemple agissant sur le milieu extracellulaire, ou de l’ATP, etc, agissant sur le milieu intracellulaire). Le fonctionnement des canaux est contrôlé par le potentiel de membrane et ses variations. On distingue différents types de canaux qui sont potentiels dépendants : Na+, K+, etc. Ce potentiel membranaire est défini par une équation, comprenant la charge des ions. Le potentiel membranaire est fixé par tous les ions de la membrane.
Chez le calamar, au repos cet axone est plus perméable aux K+ et Na+ qu’aux autres, quand on a un influx nerveux, la membrane se dépolarise. Ceci est dû à l’ouverture et la fermeture très rapide des canaux ioniques Na+ et K+. Cette dépolarisation successive des régions des membranes plasmiques permet la transmission rapide tout le long de l’axone.
Les hormones non lipidiques sont transportées à travers la paroi par un relais membranaire (récepteur spécifique).
Transports actifs avec perméase
Présentent deux caractéristiques : le transport est couplé avec un mécanisme qui va produire de l’énergie. Le transport fonctionne contre le gradient de concentration. Il peut y avoir consommation d’ATP, dans ce cas la perméase se nomme : Pompe Na, K/ATPase, véritable pompe. Leur rôle est de maintenir le gradient ionique à travers la membrane plasmique. 3 Na+ pour deux ions K+ : les ions Na+ commencent à se fixer à des sites de haute affinité : phosphorylation de l’ATP qui modifie la pompe (changement de concentration), simultanément les ions K+ se fixent à des sites accessibles à la surface cellulaire engendrant une hydrolyse des groupements phosphates liés à la membrane entraînant un changement de conformation, les zones d’affinités sont moins importantes : libération de K+ dans le cytoplasme. Cette pompe a plusieurs fonctions essentielles pour la cellule : ajuste la pression osmotique et le volume cellulaire.
Deuxième type de transport actif lié aussi à une ATPase : transporteur ABC. Formé de deux protomères. Chacun d’eux comportent six domaines transmembranaires. Ces transporteurs, présents dans les cellules normales ou cancéreuses, sont aussi appelés PGT . Dans les cellules du foie, elles permettent d’éliminer les substances toxiques, dans les cellules cancéreuses, ces transporteurs permettent de rejeter certain types de médicaments anti cancéreux (permet à la cellule cancéreuse de résister au traitement : chimiorésistance).
Couplage d’un transport actif et passif
Si les deux transports se déplacent dans le même sens, on parle de symport.
Ex : glucose et Na+ où l’absorption du glucose s’effectue sur un transporteur transportant en même temps deux ions Na+ et une molécule de glucose. Le flux d’ions Na+, fournit l’énergie nécessaire pour que la cellule importe le glucose alimentaire. Les ions K+ sont importés par Na+. Cet échangeur Na+ (passif) et H+ (actif) entraine le PH cytosolique.Avec mouvement :
- l'endocytose : transfert du milieu extra- vers le milieu intracellulaire. C’est l’inverse pour l’exocytose. Ces phénomènes ont lieu continuellement.
- la pinocytose : entrée d’un faible volume dans la membrane plasmique et ingéré dans la cellule, sur le membrane on a des revêtements glycoprotéiques piégeant les petites particules.
Sous-types de membranes plasmiques
- sarcolemme (membrane de la cellule musculaire)
- névrilemme (membrane de la cellule nerveuse)
Article connexe
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire
Catégorie :- Biologie des membranes
- l'hémimembrane E (« E » pour exoplasmique) ; C'est la couche externe de la membrane.


Wikimedia Foundation. 2010.