- Filament D'actine
-
Filament d'actine
Un filament d'actine, ou microfilament, est un homopolymère d'actine, protéine de 42 kDa (Unité de masse atomique). C'est un constituant essentiel du cytosquelette des cellules eucaryotes, ainsi que des fibres musculaires. L'actine sous forme de filaments est parfois appelée actine F (Fibrillaire), tandis que la forme monomérique est appelée actine G (Globulaire).
Sommaire
Structure d'un filament
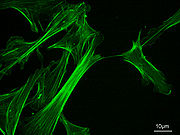
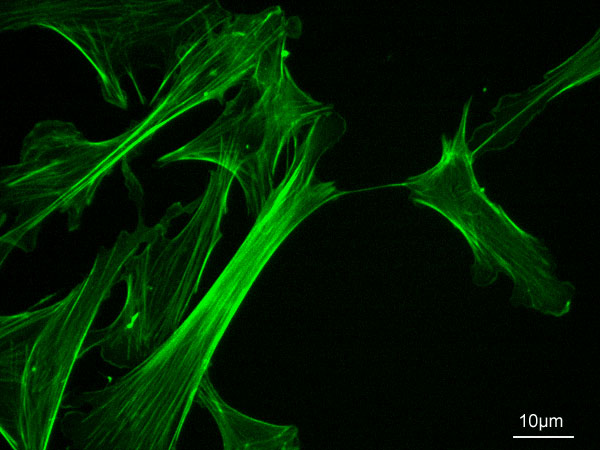
 Actine F; chaîne de 13 monomères basée sur le modèle de Ken Holmes
Actine F; chaîne de 13 monomères basée sur le modèle de Ken Holmes
Un filament d'actine a une structure en double hélice avec un diamètre d'environ 7 nm. Sa longueur de persistance est d'environ 17 µm, soit l'ordre de grandeur du diamètre des cellules.
Dynamique de polymérisation
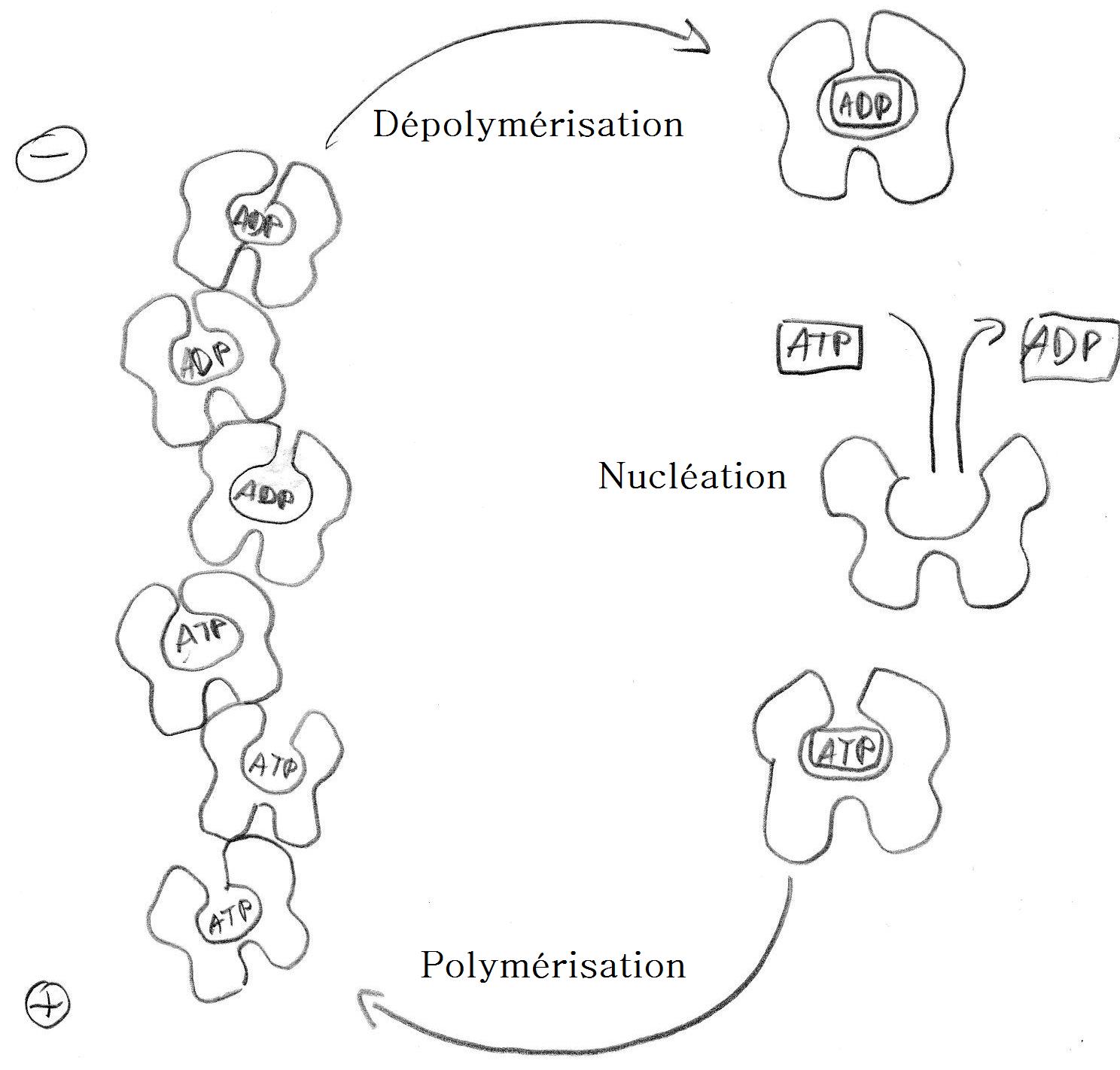
L'actine G globulaire se polymérise en actine F (filament d'actine).
La polymérisation s'amorce par une phase de nucléation, où sont formés majoritairement des trimères. Les monomères s'assemblent ensuite suivant une double hélice, qui n'a donc pas de centre de symétrie. À l'une des extrémités, notée (+) ou extrémité barbue ou encore en brosse, les constantes cinétiques sont en ordre de grandeur 10 fois supérieures à celles de l'autre extrémité, notée (-) ou extrémité pointue. En outre, les monomères associés à l'ATP (ATP-actine), présents en majorité dans les cellules vivantes, ont plus tendance à polymériser que ceux associés à l'ADP (ADP-actine).L'actine associée à un filament a tendance à hydrolyser son ATP. Cette propriété est, avec la polarité du filament, à l'origine du phénomène dit de "tapis roulant" (anglais : treadmilling). En effet, l'extrémité (+) va avoir tendance à capter en très grande majorité de l'ATP-actine, favorisant par conséquent la polymérisation à cette extrémité. En revanche, l'extrémité (-) étant moins active, l'actine du filament qui en est proche a passé plus de temps sous forme filamentaire, et est majoritairement sous forme d'ADP-actine. Par conséquent, à l'extrémité (-) l'équilibre est déplacé vers la dépolymérisation.
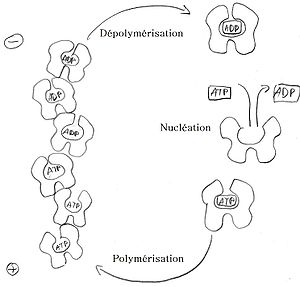 Schéma explicatif du phénomène de polymérisation du filament
Schéma explicatif du phénomène de polymérisation du filamentCes deux déplacements d'équilibre concomitants font que la chaîne croît en permanence du côté (+) et décroît du côté (-). Si on maintient un monomère central fixe, l'ensemble de la chaîne semble donc se déplacer. L'apport d'énergie nécessaire pour maintenir cet état hors équilibre se fait dans le milieu liquide environnant, où l'ADP-actine est régénérée en ATP-actine. Le terme "tapis roulant" suggère que les monomères quittant l'extrémité (-) reviennent se fixer à l'extrémité (+) après un passage en solution. Cependant, aucune masse n'est transportée macroscopiquement. Ce processus permet la réalisation d'un moteur moléculaire qui permet à certaines cellules de se déplacer au moyen d'un lamellipode. Il est aussi à l'origine du mouvement de la bactérie Listeria. Cependant, plusieurs autres protéines sont nécessaires et l'actine seule ne peut pas convertir l'énergie chimique d'hydrolyse de l'ATP en travail.
Protéines associées
Les protéines associées à l'actine (en anglais, Actin associated proteins, AAP) sont la clé du contrôle par la cellule de son stock d'actine. Elles permettent de réguler la polymérisation et d'organiser spatialement les filaments. Elles sont à leur tour contrôlées par des protéines régulatrices qui s'insèrent dans le réseau complexe interagissant avec toute la cellule.
Voici quelques exemples :
- Gelsoline : permet de passer de l'actine GEL à l'actine SOL.
- ARP2/3 : protéine proche de l'actine engendrant sa polymérisation, signal de départ.
- Fimbrine : maintient les filaments serrés d'actine fasciculée.
- Cofiline : impose une torsion supplémentaire à l'actine entraînant sa dépolymérisation.
- Villine : maintient les filaments serrés d'actine fasciculée, spécifique des microvillosités.
- Filamine : bloque l'actine réticulée pour l'empêcher de passer en actine fasciculée.
- Profiline : se fixe à l'actine monomérique aidant à la réintégration de l'actine dans le polymère à l'extrémité +
- Thymosine : se fixe à l'extrémité - de l'actine globulaire et empêche les passages ATP-ADP.
- Myosine de type 1 : maintient l'actine large fasciculée avec l'aide de l'alpha-actinine.
- Myosine de type 2: contraction musculaire.
- Myosine de type 5: protéine moteur capable de se déplacer vers l'extrémité +.
- Tropomoduline (stabilisation).
- Gelosine (désagrégation).
- Formine : nucléateur.
- Protéine de coiffe (capping protein) : se lie à l'extrémité + et empêche sa polymérisation.(Ex: protéine CAP Z)
- spectrine : permet l'accrochage du microfilament d'actine a la membrane plasmique
- alpha actinine : protéine dimérique liant 2 microfilaments entre eux, les maintenant parallèles entre eux. (point focal d'ancrage...). Elle se lie au pôle + pour permettre l'intéraction actine-myosine notamment dans le mouvement amiboïde.
On en dénombre plusieurs dizaines.
Certaines toxines agissent sur les filaments d'actine :
- Cytochalasine: bloque la polymérisation
- Phalloïdine: s'oppose à la dépolymérisation
- Latrunculline: se fixe sur les sous-unités et empêche la polymérisation
Superstructures
Les filaments d'actine s'organisent de façons très diverses suivant leurs fonctions biologiques. Voici quelques exemples de structures observées et leurs principales propriétés : faisceaux, câbles de tension, réseaux, comètes.
Rôles
- Maintien des édifices structuraux par exemple des villosités.
- Mouvements intracellulaires: /contraction des cellules musculaires striées: ces cellules musculaires sont organisées en myofibrilles (=disposition particulière des filaments d'actines et de myosines et d'autres protéines). L'unité fonctionnelle de la myofibrille est le sarcomère qui va se raccourcir lors de la contraction. /contraction des cellules musculaires lisses: pas d'organisation sarcomérique, arrangement particulier des filaments qui parcourent la cellule transversalement, longitudinalement et en diagonale, pour s'insérer sur les corps denses.
- Les mouvements cellulaires: déplacement de l'amibe, des leucocytes (pseudopodes).
- La cytodiérèse (en fin de division cellulaire).
- La formation des jonctions cellulaires.
- Le mouvement des organites: chez les végétaux supérieur le déplacement des chloroplastes dans la cellule en fonction des conditions lumineuse fait intervenir les filaments d'actine.
Effets des toxines
Certaines toxines naturelles ont des effets sur les mircofilaments d'actine.
- Phalloïdine : se lie aux filaments d'actine et s'oppose à leur dépolymérisation
- Cytochalasine D : se lie aux extrémités (+)
- Latrunculine A : se lie au monomère d'actine-G et inhibe la polymérisation en filaments
- Jasplakinolide : induit une polymérisation amorphe avec une nucléation tous les 3 actine-G : le filament s'en trouve alors désorganisé.
Lienx externes
- (en) The inner life of the cell: Mini film très bien fait, par Biovision d'Harvard, de l'intérieur de la cellule. On voit l'actine globulaire en rose se polymériser et se dépolymériser.
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire
Catégorie : Biologie cellulaire
Wikimedia Foundation. 2010.