- Cyanotoxine
-
Les cyanotoxines sont des métabolites synthétisés par les cyanobactéries. Elles peuvent être regroupées en fonction de leur structure chimique en trois classes, soit les peptides cycliques, les alcaloïdes et les lipopolysaccharides. Il est aussi possible de les différencier selon leurs mécanismes de toxicité. On y retrouve les hépatotoxines, les neurotoxines et les dermatotoxines[1].
Sommaire
Peptides cycliques et Hépatotoxines
-

Structure moléculaire générale des microcystines
-

Structure moléculaire générale des nodularines
Alcaloïdes et Neurotoxines
-
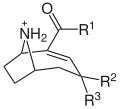
Structure moléculaire générale des anatoxines
-

Structure moléculaire générale des saxitoxines
-

Structure moléculaire générale des cylindrospermopsines (Ce n’est pas une neurotoxine, mais plutôt une cytotoxine)
Dermatotoxines et Lipopolysaccharides (LPS)
-

Structure moléculaire générale d’une endotoxine de E. coli O111:B4[2] (Hep) L-glycérol-D-manno-heptose; (Gal) galactose; (Glc) glucose; (KDO) acide 2-céto-3-déoxyoctonique; (NGa) N-acétyl-galactosamine; (NGc) N-acétyl-glucosamine.
-

Structure moléculaire de l’aplysiatoxine
-

Structure moléculaire de la debromoaplysiatoxine
-

Structure moléculaire de la lyngbiatoxine
-

Structure moléculaire de l’acide domoïque
-

Structure moléculaire de l’acide okadaïque
Les dermatotoxines se retrouvent exclusivement en milieu marin salé[3],[4], alors que les LPS sont des composantes de la paroi cellulaire de toutes les cyanobactéries.
Toxicologie
Plusieurs voies d’exposition sont possibles pour entrer en contact avec les cyanotoxines. Principalement, on retrouve l’ingestion d’eau contaminée et le contact primaire avec cette même eau. On suspecte que l’inhalation possible des toxines sous formes d’aérosols et la consommation d’organisme, qui aurait bio-accumulé une toxine, sont des sources d’exposition plausibles[5].
Une panoplie d’étude toxicologique ont été effectuées pour déterminer les effets aigus, chroniques, cancérogènes et tératogènes des différentes cyanotoxines sur plusieurs organismes (majoritairement les souris) et selon plusieurs routes d’exposition[1],[3],[5].
Le tableau suivant présente des valeurs de DL50 chez la souris, obtenues par injection intra-péritonéale[6].
Valeurs de toxicité aiguë de cyanotoxines Cyanotoxine DL50 (ug/kg) Microcystine-LR 50 Microcystine-LA 50 Microcystine-YR 70 Microcystine-RR 600 (D-Asp3) microcystine-LR 50 à 300 (D-Asp3) microcystine-RR 250 (Dha7) microcystine-LR 250 (6Z-Adda) microcystine-LR > 1200 Nodularine 50 Cylindrospermopsine (pure) 2000 Anatoxine-a et homoanatoxine-a 200 à 250 anatoxine-a(S) 20 Saxitoxine 10 À titre de comparaison, la toxicité aigüe par gavage chez la souris est située entre et 5 et 10 mg/kg de masse corporel pour la microcystine-LR, les nodularines, les cylindrospermopsines et pour l’anatoxine-a. La saxitoxine (STX) présente des valeurs typiquement plus basses, soit près de 260 ug/kg chez la souris[3].
Le tableau suivant présente des valeurs de toxicité subchronique chez la souris où la DSENO est la dose sans effet nocif observé[3].
Valeurs de toxicité aiguë de cyanotoxines Cyanotoxine DSENO (ug/kg jour) Microcystine-LR 40 (souris, gavage, 13 semaines) Cylindrospermopsine 30 (souris, gavage, 11 semaines) anatoxine-a 98 (souris, gavage, 28 jours) Les tests de toxicité pour une même cyanotoxine sont difficiles à mettre en perspective car ils ne sont pas tous effectués dans les mêmes conditions (méthodes de mise en contact, organismes testés, extraits purs vs extraits algales). Dans le dernier cas, il pourrait y avoir des effets de synergie et/ou d’antagonisme entre l’organisme étudié et tous les métabolites d’origine algale, contrairement à la seule présence de cyanotoxines. Il faut être prudent dans l’interprétation des résultats[7]..
Normes
Les normes qui découlent des études toxicologiques sont appliquées à l’eau potable et aux activités qui comportent un contact avec l’eau. Selon le cas, elles distinguent les concentrations seuil par type de cyanotoxines ou de cyanobactéries au-delà desquelles on doit prendre des précautions et/ou des actions pour remédier à la situation. Les valeurs appliquées aux cyanotoxines prennent en compte les cyanotoxines totales, c’est-à-dire celles qui sont sous la forme libre (dissoutes dans l’eau) et celles qui sont situées à l’intérieur des cellules. Elles sont exprimées en µg/L. Le tableau suivant présente les normes actuellement en vigueur et celles qui sont proposées dans divers pays[6],[8].
Concentrations maximales tolérables de différentes cyanotoxines dans l’eau potable Cyanotoxine Pays/organisation Valeurs (ug/L) Microcystines France 1,0 Microcystines Canada 1,5 Microcystines OMS 1,0 Microcystines Brésil 1,0 Microcystines Australie 1,3* Microcystines Nouvelle-Zélande 1,0 et 0,1** Nodularines# Nouvelle-Zélande 1,0 Anatoxine-a# Nouvelle-Zélande 3,0 Anatoxine-a(S)# Nouvelle-Zélande 1,0 Cylindrospermopsine# Australie 1,0 Cylindrospermopsine# Nouvelle-Zélande 3,0 Saxitoxines# Brésil 3,0 Saxitoxines# Australie 3,0**% Saxitoxines# Nouvelle-Zélande 1,0 # Valeurs proposées ou recommandées
- Valeur qui prend en compte la toxicité de l’ensemble des microcystines
- Valeur qui prend en compte l’effet de promotion tumorale
- Valeur en ug STX-équivalent/L[3]
- Valeur qui prend en compte l’effet de promotion tumorale
Les valeurs utilisées au niveau de la concentration maximale tolérable de cyanobactéries sont exprimées en # cellules/mL. Les normes en vigueur à propos de la concentration maximale tolérable dans l’eau de consommation et dans l’eau contaminée utilisée pour des fins récréatives, et avec laquelle il se produit des contacts primaires, sont de 2000 et 20 000 cellules/mL respectivement[9],[1]. Comme en moyenne une cellule contient 0,2 pigogramme de toxines[6], une valeur de 2 000 cellules/mL correspond à 0,4 µg/L de toxine totale et la valeur de 20 000 cellules/mL correspond à 4 µg/L de toxine totale. Ces valeurs (cellules/mL) sont des moyennes et sont valables seulement si toutes les cyanobactéries contenues dans le volume d’eau sont productrices de cyanotoxines.
Analyses des cyanotoxines
Les méthodes d’analyses peuvent être départagées en méthodes de criblage qui sont généralement très sensibles mais peu sélectives et en méthodes de confirmation qui sont très sélectives et présentent une sensibilité appréciable[3]. Parmi les méthodes de criblage, on retrouve les bio-essais sur différents organismes, la méthode immunologique ELISA (Enzyme linked immuno sorbent assay) et différentes méthodes enzymatiques utilisant les propriétés biochimiques des cyanotoxines. Les méthodes de confirmation sont principalement des variantes de la chromatographie liquide haute performance couplée avec différentes méthodes de détection dont la plus performante est la spectrométrie de masse, qui elle-même présente une pluralité de types d’analyseurs.
Préparation des échantillons
Un obstacle important aux méthodes d’analyse est la préparation de l’échantillon. En effet, mis à part pour les bio-essais où la préparation ne requiert que la lyse des cellules de façon à extraire les toxines intracellulaire, il est souvent nécessaire de se débarrasser de plusieurs substances pouvant créer des interférences et/ou de pré-concentrer les molécules d’intérêt.
Lyse et Extraction
Les cyanotoxines sont dissoutes dans la matrice liquide ou bien sont piégées à l’intérieur des cellules. Il est utile, voire nécessaire, de départager les échantillons en fonction de la matrice qui contient les toxines. Par filtration et centrifugation[10], on peut séparer les fractions solides, c'est-à-dire les cellules de cyanobactéries, de la matrice aqueuse constituant la fraction liquide.
Différentes techniques de lyses de la fraction solide sont utilisées comme le gel-dégel, la lyophilisation et la sonication (aussi nommée l’ultrasonication)[11]. D’autres protocoles utilisent une procédure de lyse/extraction simultanée, comme par exemple l’extraction dans un solvant en ébullition et l’utilisation d’un micro-onde, qui permet d’effectuer une extraction de type Soxhlet[12]. L’utilisation d’une extraction liquide pressurisée à température élevée est aussi utilisée[13].
Présentement, aucun solvant n’est universel pour l’extraction de l’ensemble des cyanotoxines, dû aux différentes structures moléculaires qui présentent un caractère hydrophobe ou polaire variable. De plus, le pH de la solution d’extraction est un paramètre qui est pour l’instant peu compris mais qui a fort probablement un impact sur la solubilité des composés vu les groupements ionisables présents dans la majorité des cas[11].
Le tableau suivant présente les solvants qui sont majoritairement utilisés pour l’extraction des grandes classes de cyanotoxines. Mis-à-part pour les microcystines, les autres solvants ne sont pas validés et font objet de proposition. Les conditions d’extraction des microcystines s’appliquent vraisemblablement aux nodularines qui sont des analogues structuraux des microcystines[3],[1].
Solvant d’extraction et grande classes de cyanotoxines Cyanotoxine Solvant Microcystines Eau : Méthanol* Anatoxines Eau légèrement acidifiée ou Eau : Méthanol Cylindrospermopsines Méthanol ou solution acide acétique 5 % Saxitoxines Eau : Méthanol** - % de méthanol varie entre 50 et 75 %
- % méthanol = 25 % et pH =3 (acide acétique)
Purification et pré-concentration
Un problème inhérent à l’extraction, est la co-extraction d’interférant générant un échantillon qui devient rapidement complexe. De plus, les fractions liquides, généralement aqueuse, ont une concentration très faible en cyanotoxines dissoutes. Par conséquent, il faut purifier les molécules d’intérêt avant de procéder à leur quantification ou bien concentrer celles-ci de façon à dépasser les limites de quantification de l’appareil d’analyse[1].
Ces deux objectifs peuvent être atteints par le biais de l’extraction sur phase solide qui est la méthode la plus utilisée. Dans cette technique, on fait passer la solution au travers d’une cartouche rempli de particules et on élue avec un solvant adéquat nos analytes d’intérêts qui ont été adsorbés sur la phase solide. La phase solide la plus courante est une phase greffée C18 qui permet de retenir efficacement les microcystines et les nodularines. Cependant, cette phase solide est peu sélective pour les autres cyanotoxines qui sont plus polaires. Récemment, une étude a démontrée les capacités de rétention des cartouches qui utilisent du charbon noir sous forme de graphite. Cette phase solide agit principalement comme une phase greffé C18, mais lorsqu’utilisée sous certaines conditions elle peut agir comme échangeur d’ions cationiques[14].
Une autre méthode chromatographique de purification et de pré-concentration des microcystines utilisent les principes d’immunoaffinité. À l’aide d’anticorps polyclonaux ou monoclonaux ajouté au support solide, on peut sélectivement retenir les molécules d’intérêt et les éluer par la suite[15].
Méthodes de criblage
Les essais sur animaux ont longtemps été la méthode de détection de la toxicité des efflorescences. Évidemment, ces méthodes sont pauvres en sélectivité et en sensibilité. Ces tests ne sont maintenant utilisés que pour comprendre le mécanisme d’action des toxines et tenter de dériver des normes au niveau des doses maximales tolérables quotidiennes[1].
Méthodes biochimiques
La détection des microcystines et des nodularines peut s’effectuer à l’aide de leur activité biochimique qui est en lien avec leur mécanisme de toxicité. Ces molécules sont des inhibiteurs de protéines de types phosphatases (PP1 ou PP2A) qui exécutent la déphosphorylation de phosphoprotéines intracellulaires. Il s’agit de mesurer l’activité d’une quantité fixe de phosphatases, en fonction de différentes concentrations de microcystines/nodularines, avec une quantité fixe de substrat. Initialement, le substrat utilisé était un radio-isotope du 32P, mais des variantes utilisant un substrat détectable par spectrophotométrie ont été développées pour pallier les problèmes reliés à l’utilisation de matériel radioactif[1],[14].
Un test semblable s’applique pour la détection de l’anatoxine-a(S) par le biais de son activité sur l’enzyme acétylcholinestérase. Ce test n’est pas exclusif à cette toxine car les pesticides organophosphorés exhibent le même pouvoir d’inhibition.ref>Agence française de sécurité sanitaire de l’environnement et du travail (AFSSET), Évaluation des risques liés à la présence de cyanobactéries et de leurs toxines dans les eaux destinées à l’alimentation, à la baignade l’eau de baignade et aux autres activités récréatives, juillet 2006.</ref>. Finalement, les saxitoxines peuvent être détectées à l’aide de leur activité biologique où ils agissent comme agent bloqueur des canaux sodium de la membrane cellulaire. Le test s’effectue par l’incubation de ces neurotoxines avec des cellules cancéreuses de type neuroblastes, qui démontrent l’expression génétique de canaux à sodium, et de deux agents qui favorisent l’ouverture et le processus de retour du sodium hors de la cellule. Ceci permet de maximiser l’activité de ces canaux et d’améliorer la sensibilité du test. Après la période d’incubation, on mesure le taux de survie des cellules par colorimétrie en fonction de la concentration de saxitoxines[14].
Méthodes immunologiques
Ces méthodes sont du type ELISA et elles sont basées sur le principe de reconnaissance de motifs structuraux présents sur une molécule définie, par des anticorps. La technique est de type compétition directe, où il se produit une compétition pour le site actif de l’anticorps entre une microcystine conjuguée à une enzyme (peroxydase ou phosphatase alcaline) et une microcystine contenu dans l’échantillon à analyser. Après le temps d’incubation, le substrat est ajouté et la concentration de microcystines-enzyme liées aux anticorps est évaluée par spectrophotométrie. Une coloration intense indique une faible quantité de microcystines libres dans l’échantillon et l’inverse implique une forte concentration de microcystines[1],[14].
Méthodes de confirmation
Ces méthodes sont exclusivement des techniques chromatographiques couplées à une gamme d’analyseurs tels la détection du rayonnement UV-visible, l’émission de fluorescence et l’enregistrement du spectre d’absorption avec des barrettes de diode (PDA), ainsi que les différents analyseurs utilisés en spectrométrie de masse (SM). Les techniques chromatographiques incluent les variantes de la chromatographie en phase liquide à haute performance (CLHP), de la chromatographie en phase gazeuse (CG) et de l’électrophorèse capillaire (EC). Les variantes de la SM sont attribuables aux techniques d’ionisation des analytes et des détecteurs utilisés. Les techniques d’ionisation sont l’électronébulisation (ESI), le bombardement par atomes rapides (FAB) et l’ionisation-désorption laser assistée par matrice (MALDI), alors que le triple quadripôle (QqQ) ainsi que le temps de vol (TOF) sont utilisés comme analyseur[1].
Les méthodes de CLHP sur phase inversée utilise une phase stationnaire C18 hydrophobe qui retient en ordre décroissant les microcystines et nodularines, les anatoxines, les cylindrospermopsines et les saxitoxines. Ces dernières sont peu ou pas retenues et un agent de pairages d’ions est souvent ajouté pour augmenter la rétention de celles-ci[1],[16] Les saxitoxines sont mieux séparées en CLHP sur phase normale avec une phase stationnaire constituée de silice et de groupement amide libre[14].
Les méthodes de détection basée sur le rayonnement nécessitent la présence de groupements chromophores sur les molécules. Dans certains cas, il faut effectuer une dérivation pré ou post-colonne de façon à ajouter un groupement chromophore à l’analyte, c’est le cas des saxitoxines qui sont détectées par fluorescence[3].
Les méthodes de SM utilisent majoritairement l’ESI comme méthode d’ionisation et la détection s’effectue avec le triple quadripôle qui est une méthode de spectrométrie de masse en tandem (MS/MS). Le triple quadripôle permet différents modes d’acquisition de spectres en fonction du patron de fragmentation désirés. On retrouve, le « multiple reaction monitoring » (MRM), le « precursor ion mode » et le « selected ion monitoring » (SIM). Cet ensemble de méthodes permet d’obtenir la sélectivité nécessaire pour détecter les analogues structuraux des différentes classes de cyanotoxines grâce au patron de fragmentation[17],[4].
Une autre méthode d’analyse utilise un produit de dégradation des microcystines et des nodularines résultant d’une oxydation. Ce produit est analysé par CG/SM ou CLHP et détecté par fluorescence après avoir effectué une dérivation post-colonne. Cette méthode a été développé dû au fait que dans les matrices animales, les hépatotoxines sont liées de façon covalente aux phosphatases, et c’est seulement celles qui sont libres qui sont détectées. Par conséquent, une procédure d’oxydation au permanganate de potassium et du métapériodate de sodium permet de libérer l’acide carboxylique MMPB (acide 2-méthyl-3-méthoxy-4-phénylbutyrique) de l’acide aminé ADDA[1],[18].
Les méthodes d’EC utilisent la mobilité d’une molécule chargée soumise à un champ électrique. Comme les cyanotoxines sont sous forme anioniques ou cationiques en fonction du pH de la matrice aqueuse, il est possible de les séparer par EC et de les détecter par UV-vis ou SM[19]. Une variante intéressante utilise l’ajout d’un surfactant à la matrice aqueuse et celui-ci agit comme phase pseudo-stationnaire. La séparation est dû aux différences de solubilisation des molécules à l’intérieur des micelles du surfactant et de la mobilité électrophorétique[20],[21].
La spectrométrie MALDI-TOF est en plein développement car elle ne nécessite pas la séparation du mélange au préalable par CLHP. De plus, cette méthode permet l’utilisation d’une très faible masse d’échantillon, et comme le triple quadripôle, on peut effectuer un balayage des ions produits (post source decay PSD) qui permet de détecter les fragments d’intérêts. Cependant, cette méthode provoque un bruit de fond important en fonction de la matrice qui est utilisée[22].
Avantages et inconvénients des méthodes
Le tableau suivant présente les limites de détection pour plusieurs méthodes analytiques[6],[3],[14].
Limite de détection des méthodes analytiques Méthode Cyanotoxines Limite de détection bio-essais (souris) Toutes 1 – 200 µg CLHP-UV Microcystines individuelles 0,02 µg/L CLHP-PDA Microcystines individuelles 0,02 µg/L CLHP-SM Microcystines individuelles 0,01 µg/L CLHP-fluorescence Saxitoxines individuelles 34 µg/L GC-SM Anatoxine-a 1 µg/L CLHP-SM/SM Cylindrospermopsine 0,2 µg/L CLHP- SM/SM Anatoxine-a 1 µg/L Inhibition des phosphatases (radioisotopes) Microcystines et Nodularines totales 0,02 µg/L Inhibition des phosphatases (colorimétrie) Microcystines et Nodularines totales 0,1 µg/L MMPB (GC-SM) Microcystines et Nodularines totales 10 µg/g de foie de poisson ELISA Microcystines et Nodularines totales 0,05 µg/L Les bio-essais sur les animaux ont l’avantage de détecter l’ensemble des cyanotoxines, mais ne démontrent que très peu de sélectivité face aux classes de cyanobactéries et encore moins pour différencier les variantes structurales à l’intérieur d’une même classe. De plus, la présence de résultats faux-positifs n’est pas négligeable et la reproductibilité des résultats est faible[23]. Malgré cela, ces tests présentent la possibilité de détecter la totalité des cyanotoxines, incluant les cyanotoxines inconnues qui pourront par la suite être investiguées avec des méthodes de confirmation de façon à déterminer leur structure[3].
Les méthodes chromatographiques démontrent la plus grande sélectivité et la meilleure quantification des différentes variantes des cyanotoxines. La détection du rayonnement est moins performante au niveau de la quantification, vu la plus faible sensibilité et sélectivité comparativement à la SM. Il est également à la portée des procédures de SM de déterminer la structure exacte des nouvelles toxines. Le manque de standards certifiés est présentement l’inconvénient majeur de l’ensemble de ces techniques qui fait en sorte que l’on ne détecte pas l’ensemble des cyanotoxines. De plus, les échantillons nécessitent une préparation préalable qui consomme du temps et génère un temps réponse relativement long[3].
Les méthodes immunologiques et biochimiques sont attrayantes vu la rapidité du temps réponse et la possibilité de détecter l’ensemble des variantes de la cyanotoxine étudiée. Cependant, ceci engendre nécessairement un manque d’information sur les variantes structurelles et dans le cas de l’ELISA, il y a un risque de réactivité croisée faisant en sorte qu’il est difficile de relier la concentration de microcystines détectées à la toxicité réelle de l’échantillon[6]. De façon générale les méthodes utilisées dépendent de l’objectif à atteindre et des coûts associés aux analyses. Ainsi, les méthodes immunologiques et biochimiques démontrent une sensibilité comparable aux méthodes de confirmation, à un moindre coût, et sont des méthodes de choix pour les analyses de routines. De plus, les qualifications du personnel technique sont moins exigeantes que dans le cas des méthodes de confirmation. Ces dernières sont principalement utilisées dans le cadre de recherche et développement[14].
Développement
Les méthodes de confirmation présentent la possibilité d’établir des méthodes d’analyses multi-toxines, qui après optimisation des conditions analytiques, permettent de minimiser les analyses et ainsi les établir comme méthodes de routine[24],[4],[17],[21].
L’étude de phénomène comme la bioaccumulation, et son impact sur la consommation d’organismes infectés, nécessite un protocole d’extraction des cyanotoxines intégrées à l’intérieur de matrices complexes. Un protocole connu sous le nom de « dispersion de la matrice sur phase solide » utilise les propriétés d’adsorption que possèdent des substances comme la silice, l’alumine et le sable. Ainsi, la matrice est mélangée avec l’adsorbant dans un mortier et l’échantillon résultant est déposé dans une petite colonne chromatographique et les analytes sont élués sélectivement[25],[26].
Les méthodes de préparation d’échantillons sont fastidieuses et plus particulièrement l’extraction sur phase solide, qui peut s’étendre sur une journée. Pour optimiser le temps réponse des analytes, une modification de l’appareillage chromatographique peut être accomplie de façon à effectuer la purification et la pré-concentration automatiquement. Pour ce faire, il faut relier une colonne de charge à la colonne analytique par un système de valve. Ensuite, l’échantillon liquide est élué sur la colonne de charge, à l’aide de pompes indépendantes du système chromatographique, et les analytes y sont sélectivement adsorbés, alors que l’éluant et les contaminants sont rejetés au rebut. Ensuite, on effectue une permutation des valves de façon à ce que la colonne de charge soit en aval de la colonne analytique et on utilise les pompes du système chromatographique pour éluer nos analytes à l’intérieur de la colonne analytique. Ceci permet de diminuer drastiquement le temps total d’analyse, soit moins d’une heure par échantillon. Cette procédure a été utilisée avec succès pour la quantification d’agents anti-infectieux dans les eaux usées[27] et il est envisageable qu’elle puisse très bien fonctionner pour les cyanotoxines.
Une nouvelle méthode d’ionisation des analytes pourraient drastiquement diminuer le temps d’analyse, soit la désorption thermique induite par diode laser couplée à l’ionisation chimique à pression atmosphérique.
 Schéma de la LDTD
Schéma de la LDTD
L’échantillon est dissous dans un solvant qui est par la suite déposé dans une plaque analytique à 96 puits. Le solvant est séché et l’échantillon solide passe en phase vapeur sous l’effet d’un bombardement laser. Ensuite, les analytes gazeux neutres sont ionisés chimiquement à pression atmosphérique. Ceci produit des ions moléculaires en moins de trois secondes qui sont par la suite analysés par spectrométrie de masse, pour un temps total inférieur à dix secondes. Les avantages de cette méthode, outre le temps excessivement bas d’analyse, est la possibilité de programmer des rampes d’élévation de température, ainsi que le temps de maintien de cette température, de façon à faire passer sélectivement les analytes en phase gazeuse. Ceci fait en sorte qu’il n’est plus nécessaire d’effectuer de séparation chromatographique avant l’analyse par spectrométrie de masse. De plus, aucune matrice et solvant ne sont utilisés, réduisant la présence de bruit de fond[28]. Cette technologie a été utilisée avec succès dans le cadre de l’étude de l’inhibition du cytochrome P450[29]. Il serait donc envisageable d’analyser les cyanotoxines par cette méthode à condition que les analytes se volatilisent avant de se dégrader sous l’effet de la chaleur.
Notes et références
- World Health Organisation (WHO), Toxic cyanobacteria in Water : A guide to their public health consequences, monitoring and management, 1ère edition, 1999.
- Ohno N, Morrison DC. Lipopolysaccharide interaction with lysozyme: binding of lipopolysaccharide to lysozyme and inhibition of lysozyme enzymatic activity. J Biol Chem 264:4434–4441. 1989.
- Agence française de sécurité sanitaire de l’environnement et du travail (AFSSET), Évaluation des risques liés à la présence de cyanobactéries et de leurs toxines dans les eaux destinées à l’alimentation, à la baignade l’eau de baignade et aux autres activités récréatives, juillet 2006.
- J. Dahlmann, W. R. Budakowski, B. Luckas, Liquid chromatography-electronspray ionisation mass spectrometry based method for the simultaneous determination of algal and cyanobacterial toxins in phytoplankton from marine waters and lakes by tentative structural elucidation of microcystins, Journal of Chromatography A 994: 45-574, 2003.
- Ministère de la santé et des services sociaux et Santé Canada, Risque à la santé publique découlant de la présence de cyanobactéries et de mycrocystines dans trois bassins versants du Sud-ouest québécois tributaire du fleuve Saint-Laurent, décembre 2001.
- C. Svrcek, D.W. Smith, Cyanobacteria toxins and the current state of knowledge on water treatment options: a review, J. Environ. Eng. Sci. 3: 155-184, 2004.
- Agence française de sécurité sanitaire de l’environnement et du travail (AFSSET), Évaluatio des risques liés à la présence de cyanobactéries et de leurs toxines dans les eaux destinées à l’alimentation, à la baignade l’eau de baignade et aux autres activités récréatives, juillet 2006.
- Syndicat français des entreprises de services et d’assainissement (SPDE), Cyanobactéries : les traitements, conférence introductive, journée cyanobactérie 6 juin 2005.
- Ministère de la santé et des services sociaux et Santé Canada, Risque à la santé publique découlant de la présence de cyanobactéries et de mycrocystines dans trois bassins versants du Sud-ouest québécois tributaire du fleuve Saint-Laurent, décembre 2001.
- World Health Organisation (WHO), Toxic cyanobacteria in Water : A guide to their public health consequences, monitoring and management, 1e edition, 1999.
- L. A. Lawton, C. Edwards, Review: Purification of microcystins, Journal of Cromatography A 912: 191-209, 2001.
- J. S. Metcalf, G. A. Codd, Microwave oven and boiling waterbath extraction of hepatotoxins from cyanobacterial cells, FEMS Microbiology Letters 184: 241-246 (2000).
- R. Aranda-Rodriguez, A. Tillmanns, F. M. Benoit, F. R. Pick, J. Harvie, L. Solenaia, Pressurized liquid extraction of toxins from cyanobacterial cells, Environmental toxicology, 20 (3), 390-396, Jun 2005.
- American Water Works Association (AWWA), Determination and significance of emerging algal toxins (cyanotoxins), 2007.
- R. Aranda-Rodriguez, C. Kubwado, F. M. Benoit, Extraction of 15 microcystins and nodularin using immunoaffinity columns, Toxicon 42 : 587-499, 2003.
- C. Dell’Aversano, G. K. Eaglesham, M. A. Quilliam, Analysis of cyanobacterial toxins by hydrophilic interaction liquid chromatography-mass spectrometry, Journal of Cromatography A 1028: 155-164, 2004.
- S. Hiller, B. Krock, A.Cembella, B. Luckas, Rapid detection of cyanobacterial toxins in precursor ion mode by liquid chromatography tandem mass spectrometry. Journal of mass spectrometry : 42 (9), 1238-1250, Sep 2007.
- J. L. Ott, W. W. Carmichael, LC/ESI/MS method development for the analysis of hepatotoxic cyclic peptide microcystins in animal tissues, Toxicon, 47 (7), 734-741, Jun 2006.
- P. Thibault, S. Pleasance, M. V. Laycock, Analysis of paralytic shellfish poisons by capillary electrophoresis, Journal of chromatography, 452, 483-501, 1991.
- N. Onyewuenyi, P. Hawkins, Separation of toxic peptides (microcystins) in capillary electrophoresis, with the aid of organic mobile phase modifiers, Journal of Chromatography A, 749, 271-277, 1996.
- G. Vasas, A. Gáspár, C. Páger, G. Surányi, C. Máthé, M. M. Hamvas, G. Borbely, Analysis of cyanobacterial toxins (anatoxin-a, cylindrospermopsin, microcystin-LR) by capillary electrophoresis, Electrophoresis, 25 (1), 108-115, Jan 2004.
- M. Welker, J. Fastner, M. Erhard, H. von Döhren, Applications of MALDI-TOF MS analysis in cyanotoxin research, Environmental toxicology, 17 (4), 367-374, Jan 2002.
- S. Pérez, D. S. Aga, Recent advances in the sample preparation, liquid chromatography tandem mass spectrometric analysis and environmental fate of microcystins in water, Trends Anal. Chem., Vol. 24, No 7, 2005.
- C. Dell’Aversano, G. K. Eaglesham, M. A. Quilliam, Analysis of cyanobacterial toxins by hydrophilic interaction liquid chromatography-mass spectrometry, Journal of Cromatography A 1028: 155-164, 2004.
- S. Bogialli, M. Bruno, R. Curini, A. Di Corcia, A. Laganá, B. Mari, Simple assay for analyzing five microcystins and nodularin in fish muscle tissue: hot water extraction followed by liquid chromatography-tandem mass spectrometry, Journal of agricultural and food chemistry, 53 (17), 6586-6592, Aug 2005.
- S. Bogialli, M. Bruno, R. Curini, A. Di Corcia, A. Laganà, Simple and rapid determination of anatoxin-a in lake water and fish muscle tissue by liquid-chromatography-tandem mass spectrometry, Journal of Chromatography A 1122: 180-185, 2006.
- P. A. Segura, C. Gagnon, S. Sauvé, A fully automated on-line preconcentration and liquid chromatography-tandem mass spectrometry method for the analysis of anti-infectives in wastewaters, Anal. Chim. Acta 604 : 147-157, 2007.
- LDTD™ Enhancing high-throughput analysis increasing analytical speed, Phytronix Technologies Product specification, 2007.
- J. Wu, C. S. Hughes, P. Picard, S. Letarte, M. Gaudreault, J-F Lévesque, D. A. Nicoll-Griffith, K. P. Bateman, High-Throughput Cytochrome P450 Inhibition Assays Using Laser Diode Thermal Desorption-Atmospheric Pressure Chemical Ionization-Tandem Mass Spectrometry, Anal. Chem. 79 : 4657-4665, 2007.
-











Wikimedia Foundation. 2010.