- Spéciation
-
Les mécanismes de
l'évolution biologiqueMécanismes non aléatoires :
Mécanismes aléatoires :
Conséquences de l'évolution :
- spéciation
- adaptation des espèces
- radiation évolutive
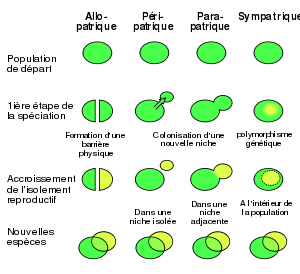 Comparaison des différents modes de spéciation.
Comparaison des différents modes de spéciation.
La spéciation est, en biologie, le processus évolutif par lequel de nouvelles espèces vivantes apparaissent. En chimie, la spéciation désigne l'ensemble des espèces chimiques sous lesquelles un élément se présente dans un environnement donné, cf. spéciation chimique. L'article traite ici de la spéciation en biologie.
Une espèce n'apparaît pas instantanément par une mutation conduisant à l'apparition d'un individu d'un type nouveau. Les espèces s'individualisent à partir de populations appartenant à une espèce d'origine (sauf pour le cas de certains végétaux, voir plus bas). Il s'agit donc de l'évolution de populations interfécondes - composant une même espèce, par définition - en populations non-interfécondes, c'est-à-dire isolées sur le plan reproducteur. Elles prennent ainsi le statut d'espèces biologiques vraies. La spéciation résulte de la sélection naturelle et/ou de la dérive génétique, qui sont les deux moteurs de l'évolution. Le concept de spéciation a été essentiellement développé par Ernst Mayr.
On distingue plusieurs types de spéciation, basés sur la répartition géographique des populations en divergence au cours du processus :
Sommaire
Spéciation allopatrique
Selon ce mode de spéciation, des populations initialement interfécondes évoluent en espèces distinctes car elles sont isolées géographiquement. C'est le mode de spéciation de loin le plus fréquent chez les animaux. Il se décline en trois modalités :
- Spéciation vicariante : une barrière géographique (rivière, montagne, vallée, océan, glacier...) coupe l'aire de répartition d'une espèce en plusieurs zones. Dans chacune des zones, chaque population évolue indépendamment des autres, pouvant donner naissance à une nouvelle espèce.
- Spéciation péripatrique ou spéciation par effet fondateur : un petit nombre d'individus fonde une nouvelle population en marge de l'aire de répartition de l'espèce d'origine, par exemple suite à la colonisation d'une île près de la côte. Cette nouvelle population de petite taille peut évoluer rapidement en une nouvelle espèce.
- Spéciation parapatrique : des populations en divergence ne sont pas totalement isolées géographiquement mais possèdent une zone de contact étroite. Les migrations entre populations sont cependant limitées puisque ces dernières se perpétuent dans des conditions environnementales différentes (gradient climatique par exemple). La sélection naturelle a donc un rôle important dans ce mode de spéciation.
Ces populations peuvent aussi revenir en sympatrie (contact secondaire), et peuvent même donner une descendance si les évolutions n'ont pas été trop importantes. Dans ce cas, il peut tout de même y avoir une spéciation s'il y a une faiblesse hybride, c'est-à-dire que la survie/reproduction de l'hybride est plus faible (même légèrement) que celle des deux populations. Dans ce cas les allèles conférant la capacité de reproduction avec les individus de l'autre population seront donc désavantagés et disparaîtront. On parle alors de mécanismes de renforcement qui apparaissent dès qu'il y a une amorce de reproduction (faiblesse hybride).
Les espèces Panthera leo et Panthera tigris illustrent parfaitement ce concept. Actuellement, ce sont deux espèces différentes, elles ne peuvent se reproduire l'une avec l'autre qu'en captivité. Lors du contact secondaire, les lions et les tigres pouvaient donc encore se reproduire, mais les tigrons et les ligres (hybride) présentent une faiblesse (système immunitaire défaillant pour le tigron...). Les lions se reproduisant avec des tigres ont donc eu moins de descendants et ceux-ci ont progressivement disparu. Actuellement aucun lion ne se reproduit avec un tigre en conditions naturelles. Il s'agit bien d'une spéciation.
Dans l'espèce Canis familiaris, on comprendra aisément que la fécondation réussie de femelles chihuahua par des mâles saint-Bernard pose une double difficulté exclusivement physique :
- la possibilité pratique de l'accouplement,
- celui d'une arrivée de la gestation à bon terme,
même si les gamètes sont compatibles in vitro : la raciation commencerait dans ce cas précis - sans l'existence d'intermédiaires - son cheminement vers une possible spéciation.
A contrario, certaines races de Canis lupus sont interfécondes avec Canis familiaris et les croisements donnent une descendance féconde. Les notions de race et d'espèce contiennent donc parfois une réduction simplificatrice de la complexité du vivant. (voir Classification). Il faut noter qu'actuellement cet exemple ne semble plus pertinent, les chiens (Canis familiaris) étant considérés comme une sous-espèce de Canis lupus sous le nom de Canis lupus familiaris.
Spéciation sympatrique
Des populations non isolées géographiquement peuvent évoluer en espèces distinctes. Ici, la sélection naturelle joue un rôle crucial dans la divergence des populations. Ce phénomène reste controversé, mais semble prouvé chez plusieurs poissons (notamment les cichlidés), ou chez certains insectes phytophages. La différenciation des espèces pourrait aussi résulter du conflit sexuel.
Variante : chez certains végétaux supérieurs (angiospermes), deux espèces normalement non-interfécondes peuvent engendrer des hybrides féconds, sous certaines conditions liées au doublement du nombre de chromosomes présent dans l'hybride formé (diploïdie). Il peut alors lui-même se perpétuer tout en étant incapable de se reproduire avec des individus appartenant aux espèces de ses parents. C'est donc le premier représentant d'une nouvelle espèce, sans faire intervenir un isolement géographique. On parle donc là aussi de spéciation sympatrique, qui semble très fréquente chez les espèces herbacées.
Un exemple plus rare de spéciation sympatrique est la divergence entre "résident" et "transient" chez l'Orque dans le pacifique[1]. Les deux formes habitent le même océan mais ne chassent pas les mêmes proies, n'ont pas les mêmes émissions vocales et ne se reproduisent pas entre eux. Des divergences entre les espèces peuvent aussi provenir des contraintes de micro-habitats contrastés. Ainsi, le putois présente un phénotype rare dit dark qui est directement dépendant des ruisseaux boisés et établit une convergence avec la physionomie du très rare vison d'Europe[2].
Vitesse de spéciation
L’histoire de l’évolution montre que les spéciations nécessitent généralement plusieurs centaines de milliers d’années. Toutefois, certaines nouvelles espèces de souris comme celles de Madère n'ont nécessité qu'entre 500 et 1000 ans pour apparaître (en fonction de l’origine viking ou portugaise qu’on leur attribue) ce qui représente environ 2000 à 4000 générations soit une vitesse extrêmement rapide.
En opposition avec cette vision ("gradualisme phylétique"), la théorie des équilibres ponctués (Stephen Jay Gould, Niles Eldredge) postule que cette histoire comprendrait de longues périodes d'équilibre, ponctuées de brèves périodes de changements importants comme la spéciation ou les extinctions.
Notes et références
- Burden et al.: Resident And Transient-Type Killer Wales SC/56/SM15.
- Thierry Lodé "Genetic divergence without spatial isolation in polecat Mustela putorius populations". J Evol Biol 14:228-236, 2001
Voir aussi
Articles connexes
- Évolution
- Écologie du paysage
- Fragmentation écopaysagère
- Insularisation écologique
- Variation clinale
- Conspécificité
- Vicariance
 Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant

Wikimedia Foundation. 2010.