- Trois règnes du vivant
-
Classification phylogénétique
La classification phylogénétique est un système de classification des êtres vivants. Elle tend à remplacer la classification traditionnelle en se basant uniquement sur les rapports de proximité évolutive entre espèces. L'œuvre de Willi Hennig Grundzüge einer Theorie der phylogenetischen Systematik[1] (Fondements d'une théorie de la systématique phylogénétique), publiée en Allemagne en 1950, est à l'origine de la classification phylogénétique. Ayant traduit son livre en anglais en 1966 Hennig trouva un plus large auditoire et révolutionna ainsi toute la systématique à partir de la fin des années 1960.
Sommaire
Méthode de classification
 La systématique phylogénétique établit trois types principaux de rapport au sein de groupes d'êtres vivants susceptibles de former, ou pas, un taxon valide et admis au sein de la classification. L'admissibilité des taxons se base uniquement sur le critère de la monophylie : en jaune les sauropsides (groupe monophylétique, car incluant un ancêtre et tous ses descendants). En bleu les reptiles (groupe paraphylétique, car n'incluant pas tous les descendants du même ancêtre, effectivement les oiseaux sont exclus de la définition des reptiles, mais partagent avec eux le même ancêtre commun). En rouge mammifères et oiseaux sont réunis sous le caractère de l'homéothermie, mais cela donne un groupe polyphylétique car ils n'ont pas développé ce caractère à partir du même ancêtre commun.
La systématique phylogénétique établit trois types principaux de rapport au sein de groupes d'êtres vivants susceptibles de former, ou pas, un taxon valide et admis au sein de la classification. L'admissibilité des taxons se base uniquement sur le critère de la monophylie : en jaune les sauropsides (groupe monophylétique, car incluant un ancêtre et tous ses descendants). En bleu les reptiles (groupe paraphylétique, car n'incluant pas tous les descendants du même ancêtre, effectivement les oiseaux sont exclus de la définition des reptiles, mais partagent avec eux le même ancêtre commun). En rouge mammifères et oiseaux sont réunis sous le caractère de l'homéothermie, mais cela donne un groupe polyphylétique car ils n'ont pas développé ce caractère à partir du même ancêtre commun.
La classification traditionnelle classait les espèces selon la présence ou l'absence d'une multitude de caractères (biologiques, phénotypiques, anatomiques, physiologiques, éthologiques, de comportement alimentaire etc) alors que la classification phylogénétique classe uniquement selon la méthode cladistique, exposée par Willi Hennig dans son ouvrage de 1950. Selon cette méthode seuls les caractères empiriquement observables et propres au rang d'espèce doivent être retenus, car, étant les seuls témoins de l'héritage ancestral de chaque espèce, ils sont les seuls qui peuvent donner un critère objectif de classification entre groupes distincts[2]. Ainsi les caractères phénotypiques (qui ne sont pas propres au rang d'espèce mais à celui de sous-espèce) peuvent servir à l'étude de populations actuelles concrètes, mais pas à l'analyse phylogénétique. La classification phylogénétique est donc un système de classification des êtres vivants d'abord fondé sur ce que les êtres vivants ont (au niveau morphologique comme au niveau moléculaire), et non pas, en première instance, sur ce qu’ils n’ont pas (on évitera alors « invertébrés ») ou sur ce qu’ils font (on évitera alors « vivipares », « fouisseurs »)[3]. Ce n'est une classification ni écologique ni anthropocentrique. Elle regroupe les êtres vivants sur un type particulier de ressemblances, celles des caractères évolutivement innovants au sein de l'échantillon d'espèces à classer. Elle s'oppose en cela aux classifications phénétiques qui ne trient pas la ressemblance et font des classements à partir de la ressemblance globale. Elle remplace la classification traditionnelle qui, elle, est fondée sur une foule de critères : morphologiques mais aussi écologiques, éthologiques, anthropocentriques, et de préférences nutritionnelles[4],[2],[5].
Le partage entre espèces d'un caractère ou d'un certains nombre de caractères jette sur ces mêmes espèces le soupçon d'une origine commune qui remonte jusqu'à l'existence d'un ancêtre commun, le premier à avoir acquis ce caractère ou ensemble de caractères. L'existence de l'ancêtre peut donc être découverte grâce à la méthode cladistique, mais pas son identité, qui reste cachée. Ainsi par exemple les oiseaux partagent tous un ancêtre commun, mais la découverte en 1861 d'un fossile comme Archaeopteryx, qui est le plus ancien oiseau connu, ne prouve pas que ce fossile en particulier soit l'ancêtre de tous les oiseaux. Effectivement une découverte future pourrait mettre à jour un oiseau fossile plus ancien qu'Archaeopteryx, mais à nouveau la certitude d'être en face d'un « ancêtre » est inexistante. Les rapports d'ancêtre à descendants (la généalogie) ne peuvent être identifiés en tant que tels que si l'identité même de l'ancêtre et des descendants est préalablement connue. Autrement dit, pour retracer la généalogie, la science de la classification devrait avoir la certitude de connaître toutes les espèces existantes et ayant existé. Comme ce n'est pas le cas, car la science est loin de pouvoir connaître la totalité des espèces vivantes et fossiles, la généalogie, même si elle a réellement eu lieu dans le passé, ne peut être retracée. Ce que la science de la classification peut retracer, avec ces mêmes éléments partiels que sont les quelques espèces fossiles et actuelles connues, ce sont les rapports de parenté entre espèces. Telle est la différence entre une généalogie (« qui est ancêtre de qui ») et une phylogénie (« qui est le plus proche parent de qui »). Les rapports phylogénétiques entre espèces connues constituent ainsi le seul critère objectif possible de classification[2].
La systématique moderne prend en compte tous les caractères héritables et même les pertes secondaires de caractère, pertes secondaires que l'embryologie, par exemple, peut mettre en lumière. Les caractères vont de ce qui est visible (anatomie et morphologie, fondement de la classification traditionnelle) jusqu'aux séquences d'ADN et d'ARN, en passant par les protéines et les données de la paléontologie. Le séquençage de certaines parties du génome, comme l'ADN des mitochondries ou l'ARN des ribosomes a permis dans les dernières années de faire des progrès importants dans la classification et de résoudre maints problèmes séculaires[6].
Les caractères et leurs ressemblances sont donc le point de départ de la méthode cladistique, mais de ces caractères ressemblants les seuls valables pour être attribués à un taxon (un taxon est un nom de groupe, comme mammifère, ou arthropode) sont ceux qui ont été hérités d'un ancêtre commun. Cette propriété d'être un caractère hérité se dit des caractères apomorphes, partagés par au moins deux taxons, ou au moins par deux espèces. Un caractère apomorphe est un caractère ancestral, une apomorphie, mais lorsqu'au sein d'une lignée d'êtres vivants l'évolution le transforme il devient le caractère propre d'un nouveau groupe. On dit dans ce cas que c'est un caractère propre dérivé, une synapomorphie. Par exemple la synthèse du phosphate de calcium (pour se fabriquer un squelette osseux) est une synapomorphie chez tous les crâniates[7], mais c'est une apomorphie chez les mammifères, qui eux ont leurs propres synapomorphies liées à leurs os, tout comme les oiseaux ont les leurs. Le caractère « phosphate de calcium » (ou « os ») est donc synapomorphique chez les crâniates et apomorphique chez les oiseaux et chez les mammifères. Oiseaux et mammifères sont donc forcément des crâniates mais avec le caractère phosphate de calcium la méthode cladistique ne peut nommer comme taxon valable au sein de la classification que les craniâtes eux-mêmes, c'est-à-dire le groupe ayant les caractères dérivés partagés et non pas les caractères primitifs partagés[8]. Oiseaux et mammifères seront donc définis selon leurs propres synapomorphies. Comme la construction de la classification se fait en subordonnant les taxons les uns aux autres il en résulte une organisation en forme d'arbre, où le vivant dans son ensemble est représenté par le tronc de l'arbre. Le tronc se divise d'abord en branches qui se divisent ensuite à leur tour en d'autres branches. Chacune de ces branches, appelées « clades » (du grec κλάδος, klados, « branche »), est un taxon disposant d'au moins une synapomorphie qui le caractérise et qui le rend valable au sein de la classification. Les points d'où bourgeonnent les branches les unes à partir des autres, les nœuds, représentent les ancêtres de chaque groupe. L'expression graphique d'un clade ou ensemble de clades est un cladogramme.
Les cladogrammes qui constituent le grand cladogramme qu'est l'arbre du vivant, de par le nombre gigantesque de caractères et d'espèces pris en compte, font appel à des algorithmes complexes exécutés par des logiciels spécialisés. Des algorithmes différents peuvent donner des résultats différents. Dans ce cas, celui qui répondra le plus au critère de parcimonie sera retenu. C’est-à-dire que de tous les arbres possibles, de tous les cladogrammes possibles, le plus parcimonieux sera celui demandant le minimum de transformations de caractères. L'arbre du vivant est, en effet, un ensemble de points de branchements, de niveaux. Chaque niveau étant assimilé à un nœud les nœuds sont obligatoirement un organisme théorique qui posséderait les synapomorphies partagées par les nœuds postérieurs dans le cladogramme. Si un nouvel organisme (fossile ou vivant) est découvert, possédant ou pas ces synapomorphies ou en possédant d'autres, un nouvel arbre doit être construit. À terme, si l'arbre restitue la totalité des relations de parenté, tous les branchements devraient être binaires. Le sens des dichotomies n'a rien à voir avec la sexualité (comme l'expression des mariages dans un arbre généalogique) ni avec des spéciations binaires (une ou deux espèces évoluant à partir d'une autre), car l'arbre n'est pas une généalogie (« qui descend de qui ») mais reflète uniquement un pouvoir explicatif maximal de l'arbre : un arbre totalement dichotomique réussit à restituer uniquement toutes les relations de parenté (« qui est plus proche de qui ») au sein d'un groupe d'organismes soumis à la méthode. La classification actuelle est continuellement remaniée en fonction de nouvelles informations[9], mais les exemples de classification peuvent toujours avoir à être modifiés car ils dépendent constamment de l'arrivée de nouvelles données empiriques. Par exemple selon la classification de Lecointre et Le Guyader la première division de l'ensemble du vivant est pour l'instant réduite à trois clades :
Savoir lesquels de ces trois groupes partagent un ancêtre commun qui les distingue du troisième est un sujet de recherche, comme ce l'est d'ailleurs avec tous les taxons non binaires (les « arbres non enracinés », ceux que la recherche n'a pas encore pu diviser en deux taxons de base). Certains chercheurs ont déjà proposé leur propre cladogramme, faisant de deux de ces trois clades un ensemble de deux groupes frères, ensemble qui serait à son tour le groupe frère du troisième. Par exemple Colin Tudge a proposé un arbre enraciné où les archées et les eucaryotes sont un ensemble de deux groupes frères, ensemble qui à son tour est le groupe frère des eubactéries[10]. La classification de Tudge n'est qu'un exemple car les chercheurs sont réellement divisés quant aux différentes positions prises à ce sujet.
Exemple de l'homme
Un exemple détaillé permet d'avoir une idée de la différence de résultat avec l'approche traditionnelle. Étant nous-mêmes des humains prenons l'exemple de l'homme, voici donc une partie des nœuds successifs permettant de classer l'homme telle qu'apparaissant dans le livre Classification phylogénétique du vivant, de Guillaume Lecointre et Hervé Le Guyader. Certains intermédiaires ont été omis arbitrairement par les auteurs pour montrer un condensé de l'arbre, le nombre de taxons mis en évidence par la méthode étant de très loin supérieur. La description associée à chaque clade est peu rigoureuse et vise juste à fixer les idées :
- Espèce Homo sapiens
- Genre Homo : inclut les espèces fossiles comme Homo neandertalhensis ou Homo erectus
- Hominines : les genres Australopithecus et Homo
- Homininés : les deux espèces de chimpanzés et les hominines
- Hominidés : gorille et Homininés (famille Hominidae dans la classification classique)
- Hominoïdés : on ajoute l'orang-outan
- Hominoïdes : on ajoute les gibbons
- Catarrhiniens : les singes de l'ancien monde
- Simiiformes : singes
- Haplorrhiniens : singes et tarsiers
- Primates : la plus grande partie de l'ancien ordre Primates dans la classification classique
- Euarchontes : s'ajoutent les Scandentiens (toupayes) et Dermoptères
- Euarchontoglires : s'ajoutent les Glires (lapins et rongeurs)
- Boréoeuthériens : s'ajoutent les Laurasiathériens (insectivores, carnivores et ongulés)
- Euthériens : s'ajoutent les Xénarthres et les Afrothériens pour compléter les mammifères placentaires
- Thériens : s'ajoutent les marsupiaux
- Mammifères : l'ancienne classe du même nom, les précédents plus les monotrèmes (ornithorynques et échidnés)
- Amniotes : s'ajoutent les anciennes classes des reptiles et des oiseaux
- Tétrapodes : s'ajoutent en gros les amphibiens
- Sarcoptérygiens : s'ajoutent dipneustes puis cœlacanthes
- Ostéichthyens : s'ajoutent les actinoptérygiens (la majeure partie des « poissons osseux »)
- Gnathostomes : s'ajoutent les chondrichtyens (requins et raies)
- Vertébrés : puis les lamproies
- Crâniates : s'ajoutent les myxines
- Chordés : s'ajoutent amphioxus et ascidies
- Deutérostomiens : s'ajoutent les échinodermes (étoiles de mer, oursins…) et hémicordés
- Bilatériens : animaux symétriques : s'ajoutent aux précédents, entre autres, les protostomiens qui incluent, entre beaucoup d'autres, différents groupes de vers, les arthropodes (insectes), les mollusques (escargots)
- Eumétazoaires : animaux organisés, s'ajoutent aux précédents les cnidaires par exemple
- Métazoaires : l'ancien règne animal, s'ajoutent aux précédents différents groupes d'éponges
- Holozoaires : s'ajoutent les Choanoflagellés et les Mésomycétozoaires, protozoaires plus proches parents des animaux que des champignons
- Opisthocontes : s'ajoutent, entre autres, les champignons
- Unicontes : s'ajoute la plupart des amibes
- Eucaryotes : Le domaine Eukaryota de la classification classique, êtres vivants à cellules à noyau ; s'ajoutent aux précédents listés ci-dessus la lignée verte (algues vertes, algues rouges, plantes à fleurs), la lignée brune (algues brunes…) et toute une collection de groupements d'espèces unicellulaires
De bas en haut de cette liste les taxons sont tout autant de clades qui incluent tous ceux qui se trouvent au-dessus d'eux. L'homme est ainsi un homininé comme l'est aussi le chimpanzé, un vertébré comme le sont aussi le chimpanzé et la baleine bleue, un bilatérien comme le sont aussi le chimpanzé, la baleine bleue et la mouche. Finalement il est aussi un eucaryote comme le sont aussi le chimpanzé, la baleine bleue, la mouche et la carotte. Les rangs taxinomiques parfois cités en marge (ordre, famille, embranchement etc) n'ont de sens que dans les classifications dites traditionnelles ou classiques. La classification phylogénétique ne conserve que les deux premiers rangs, l'espèce et le genre, car la nomenclature binominale elle-même est aussi conservée afin de pouvoir continuer à donner nom aux espèces[11].
Différences avec la classification traditionnelle
Une des caractéristiques de l'approche phylogénétique est que cette classification bouleverse toutes les classifications l'ayant précédée, autant les classifications de biologies fixistes (comme celles développées par Carl von Linné ou par Georges Cuvier) que celles qui suivirent Darwin et qui incluaient la sélection naturelle comme cause de la spéciation. La classification de Linné reposait sur l'adage que toutes les espèces sont apparues en même temps et que celles-ci étaient fixes, alors que la classification phylogénétique illustre les principes d'évolution et de sélection naturelle. Les classifications post-darwiniennes avaient déjà inclus ces mêmes principes évolutifs mais en établissant leur critère de classification sur les rapports d'ancêtre à descendant (généalogie) alors que la classification phylogénétique se base sur le critère de la plus proche parenté entre espèces (phylogénie). L'arrivée de la théorie de l'évolution introduisit l'idée que les taxons ont évolué les uns à partir des autres mais elle ne modifia pas immédiatement les critères de classification, qui jusqu'à l'arrivée de la cladistique proposée par Hennig restèrent essentiellement les mêmes que ceux du temps de Linné.
 Dans la classification classique, le groupe des reptiles ne comprend pas les oiseaux, or d'après la classification cladistique, les plus proches cousins des crocodiles sont les oiseaux, et non les lézards ou les serpents par exemple. Le regroupement des crocodiles, lézards et serpents sous le nom de reptile n'est donc pas pertinent.
Dans la classification classique, le groupe des reptiles ne comprend pas les oiseaux, or d'après la classification cladistique, les plus proches cousins des crocodiles sont les oiseaux, et non les lézards ou les serpents par exemple. Le regroupement des crocodiles, lézards et serpents sous le nom de reptile n'est donc pas pertinent. Arbre simplifié du vivant selon la classification phylogénétique : trois groupes principaux ou empires sont proposés : les bactéries, les archées et les eucaryotes.
Arbre simplifié du vivant selon la classification phylogénétique : trois groupes principaux ou empires sont proposés : les bactéries, les archées et les eucaryotes.La classification phylogénétique ne validant que des groupes caractérisés par des caractères dérivés propres (les synapomorphies) ces groupes sont aussi dits monophylétiques, c'est-à-dire d'une seule phylogénie, d'une seule phyliation : celle d'un ancêtre et de tous ses descendants. Les classifications qui précédèrent la classification phylogénétique ne tenaient pas compte de la phylogénie mais uniquement d'éléments qui pouvaient parfaitement être contingents ou anthroponcentriques (comme le comportement ou comme la privation de caractères humains[2],[5]), même dans le cas des classifications qui acceptaient la théorie de l'évolution. La classification classique, que ce soit sous son ancienne forme créationniste (Linné, Cuvier) ou sous sa forme évolutioniste post-darwinienne, a ainsi formé des taxons qui n'étant pas basés sur le principe d'un ancêtre et de tous ses descendants sont appelés paraphylétiques par la méthode cladistique. Les reptiles en sont un exemple connu. Le groupe d'animaux appelés « reptiles » partage les mêmes ancêtres que ces autres groupes qui étaient appelés « oiseaux » ou « mammifères » au sein de la même classification. Le critère de la monophylie (un ancêtre et tous ses descendants) étant appliqué le groupe des reptiles est identifié comme ne constituant pas un groupe naturel et il se voit donc chassé de la classification. Il est toujours en usage dans la langue quotidienne que de prétendre que les serpents, les crocodiles, les tortues ou les iguanes sont des « reptiles » mais aux yeux des méthodes et des découvertes modernes ce mot n'a plus la valeur de taxon qu'il avait auparavant dans la classification classique.
Une autre différence avec la systématique traditionnelle est que la systématique phylogénétique rejette toute catégorisation des niveaux hiérarchiques[12]. Pour des raisons pratiques, l'arbre du vivant donne lui-même la hiérarchie que tentaient de fournir les anciennes catégories qu'étaient les rangs taxinomiques du système linnéen. Ce système exprimait l'idée anthropocentrique et non objective d'une échelle des êtres, une hiérarchie dans laquelle l'homme était le couronnement[5], et ce autant dans le modèle fixiste créationniste que dans le modèle évolutionniste pré-phylogénétique. Au contraire la classification phylogénétique offre une vision des êtres vivants qui n'attribue pas de prééminence à certains êtres vivants sur certains autres. L'évolution les a produits en leur donnant la propriété d'être tous adaptés à leur milieu, ce qui les rend égaux face à une tentative de les organiser selon un quelconque ordre hiérarchique qui soit basé sur le critère d'un jugement de valeur. La hiérarchie de la classification phylogénétique est strictement celle de l'emboîtement des taxons les uns à l'intérieur des autres. Les vers de terre ou les cafards ne sont donc pas inférieurs aux êtres humains du point de vue de la classification scientifique, ils occupent leur position dans l'arbre du vivant tout comme les humains occupent la leur. Les jugements de valeurs appelant à une supériorité de l'espèce humaine sur les autres espèces ne se font donc plus dans le domaine des sciences naturelles mais uniquement dans les domaines non-scientifiques ou non-empiriques, ceux qui sont intrinsèquement liés à une idée de transcendance, comme par exemple la religion, la philosophie, la métaphysique ou voire même la politique, la sociologie, la déontologie de la médecine etc.
Citons enfin quelques exemples de changements spectaculaires par rapport à la classification traditionnelle :
- Les oiseaux sont des dinosaures, car tous les ancêtres des dinosaures sont aussi des ancêtres des oiseaux[8].
- Les plus proches parents vivants des oiseaux sont les crocodiles[8].
- Le concept de « reptile » est abandonné en tant que groupe valide naturel, car paraphylétique[8].
- Le concept de « poisson » est abandonné en tant que groupe valide, tout comme celui de reptile[13].
- Les ostéichthyens (anciennement les « poissons osseux ») sont plus proches des mammifères que des requins[14].
- Les plus proches parents des cétacés seraient les hippopotames[15]. Le concept d' « artiodactyles », ancien groupe incluant les hippopotames mais excluant les cétacés, est donc abandonné.
- L'ancien groupe des « algues » explose en tous sens, certaines étant groupées avec les plantes vertes, d'autres avec les bactéries[16],[17].
- Le concept de « protiste » est abandonné, au profit de groupes pouvant mélanger êtres multicellulaires et monocellulaires (par exemple les straménopiles regroupent les algues brunes - dont les kelp, jusqu'à 60 m de long - et les diatomées unicellulaires)[18],[16].
- La division entre Procaryotes et Eucaryotes est soumise à débat, au profit de la division provisoire en trois du vivant[19].
- L'application de cette classification aux angiospermes est illustrée par la classification APG (Angiosperms Phylogeny Group)[20].
Éléments conservés de la classification traditionnelle
Exemple de classification
présente dans les articles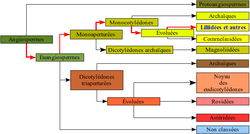
Classification classique puis phylogénétique d'une
plante à fleur, le Lis des Pyrénées. Lorsque la
classification phylogénétique suit la classification
classique, le nom du rang taxinomique (division,
classe, ordre, famille, etc) ainsi que le nom du
taxons (en latin) peuvent être gardés. Dans le
cas contraire (comme les angiospermes) le rang
est simplement appelé clade, un nouveau nom est
donné, et la classification classique n'est plus
suivi, elle peut néanmoins être de nouveau suivi à
partir d'un rang inférieur.Classification classique Règne Plantae Sous-règne Tracheobionta Division Magnoliophyta Classe Liliopsida Sous-classe Liliidae Ordre Liliales Famille Liliaceae Genre Lilium Classification phylogénétique Empire Eucaryota Règne Plantae Clade Angiospermes Clade Monocotylédones Ordre Liliales Famille Liliaceae Genre Lilium Nom binominal Lilium pyrenaicum
Gouan, 1773Parcourez la biologie sur Wikipédia : 





La classification phylogénétique a évincé les critères arbitraires et non objectifs de classification, les anthropocentrismes et les rangs taxinomiques. Elle conserve par contre :
- La nomenclature binominale en latin pour nommer les espèces. Deux rangs taxinomiques sont donc conservés, l'espèce et le genre, mais ils n'établissent pas une hiérarchie autre que celle du simple emboîtement des taxons.
- Le latin comme langue véhiculaire pour nommer les taxons.
- L'idée vague d'un arbre évolutif. Cette notion est conservée sous la forme des cladogrammes, mais les rapports entre taxons sont strictement phylogénétiques.
Notes
- ↑ HENNIG Grundzüge einer Theorie der phylogenetischen Systematik, Deutscher Zentralverlag, Berlin 1950.
- ↑ a , b , c et d LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 13, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 6, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 6, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ a , b et c LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 17, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 39, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ http://perezyvan.free.fr/Gabriel_Neve/PDFcours2006/TP2_2006.pdf Méthodes de Classification Phylogénétique, page 6.
- ↑ a , b , c et d LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 16, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ Voir le site treebase qui fait référence en la matière
- ↑ (en) Colin Tudge, The Variety of Life, Oxford University Press, 2000
- ↑ « L’espèce n’est qu’une collection monophylétique d’individus, définie au mieux par une synapomorphie, au pire par la moyenne et la variance de paramètres mesurés » in LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 18, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 23, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 520, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 321, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 448, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ a et b LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 517, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 518, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 118, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ LECOINTRE Guillaume et LE GUYADER Hervé, Classification phylogénétique du vivant, page 50, 2e édition, 2002, Belin, Paris, ISBN 2-7011-2137-X
- ↑ http://www.botanique.org/classification/angiosperm-phylogeny-group-apg-ii/rubrique576.html rubrique APG II du site bonatique.org
Liens internes
- Cladistique
- Phylogénie
- Arbre phylogénétique
- Classification scientifique des espèces
- Systématique
- Taxonomie
- Théorie de l'évolution
- Caractère ancestral et caractère dérivé
- Classification phylogénétique de Guillaume Lecointre et Hervé Le Guyader
- Eubacteria (classification phylogénétique)
- Photo-guide taxinomique du monde animal
Liens externes
- (en) Accueil de l'Angiosperm Phylogeny Group
- Arbre de la vie
- svt 44
- Treebase
- Projet Tree of Life
- Méthodes de classification phylogénétique
- Théorie des chaînes
- arbre taxonomique du Muséum national d'Histoire naturelle : [1]
Taxinomie Règne - Sous-règne - Infra-règne -
Superembranchement - Embranchement / Division - Sous-embranchement -
Superclasse - Classe - Sous-classe - Infraclasse -
Superordre - Ordre - Sous-ordre - Infraordre
Superfamille - Famille - Sous-famille - Tribu -
Genre - Sous-genre - Section - Série -
Espèce - Sous-espèce -
Variété ou Race - Forme ou TypeBiologie - Nomenclature - Nom binominal  Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant Portail de la biologie
Portail de la biologie
Catégories : Phylogénétique | Classification



Wikimedia Foundation. 2010.