- Tissu méristématique
-
Méristème
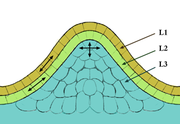 Schéma de la représentation en couches d'un méristème apical.
Schéma de la représentation en couches d'un méristème apical.
Chez les embryophytes (plantes vertes), un méristème est un tissu biologique constitué de cellules indifférenciées (ou peu différenciées) formant une zone de croissance où ont lieu les divisions cellulaires (mitoses). On distingue habituellement les méristèmes primaires, qui assurent la croissance de la plante en longueur, au niveau de la tige, des feuilles ou des racines, et les méristèmes secondaires, responsables de la croissance en diamètre des organes de certaines plantes, comme le tronc.
Sommaire
Méristèmes et développement
Seul tissu végétal capable de créer de nouvelles cellules, le méristème est essentiel au développement des végétaux. Il crée ainsi toute la plante : il est à l'origine de la tige, des feuilles, des racines, des fleurs (chez les plantes à fleur) ou encore des branches. Il est en outre responsable de la disposition particulière des feuilles ou des branches autour de la tige, ce qu'on appelle la phyllotaxie.
Les méristèmes dans le monde végétal
Selon les différents groupes, les méristèmes sont différents.
Mousses et ptérydophytes
Les mousses et les ptéridophytes (dont les fougères) possèdent une unique cellule apicale de forme tétraédrique, qui présente donc quatre faces. À chaque cycle cellulaire, cette cellule se divise et donne une nouvelle cellule sur une de ses faces, puis chaque nouvelle cellule est produite par chaque face, et ce de manière cyclique.
Gymnospermes
Les gymnospermes (dont les conifères) possèdent un tissu méristématique constitué de deux couches de cellules (si l'on considère l'organisation en couches) : une première couche en surface dont les cellules ont une division anticline uniquement, c'est-à-dire perpendiculairement à l'axe considéré, et une couche en dessous dont les divisions sont anticlines et périclines, c'est-à-dire selon l'axe considéré.
Angiospermes
Les plantes à fleur possèdent un tissu méristématique similaire aux gymnospermes, mais avec une couche intermédiaire (L2) entre la couche externe (L1) et la couche interne (L3), dont les cellules ont une division majoritairement anticline. (voir plus loin l'organisation des tissus méristématiques)
Croissance indéfinie
Bien que chaque plante croîsse selon ses propres règles, chaque méristème peut continuer à croître tant qu'il est en vie : chez bon nombre de plantes la croissance méristématique est potentiellement indéterminée, ce qui suppose que la forme complète de la plante n'est pas déterminée à l'avance, comme c'est le cas chez les animaux bilatériens qui disposent d'un plan d'organisation défini.
Histogenèse et organogenèse
Les méristèmes forment des tissus (histogenèse) tels que des tissus conducteurs, xylèmes et phloèmes, et des organes (organogenèse), comme les feuilles, eux -même constitués de tissus avec une organisation particulière.
Le méristème racinaire et les méristèmes apicaux de tiges produisent en continu des tissus, qui permettent à la racine et à la tige de croître sans pour autant créer de nouveaux organes. De même, les méristèmes secondaires produisent des tissus, notamment pour la croissance en épaisseur.
La formation d'organes sous-entend la formation au niveau du méristèmes d'ébauches (des ébauches foliaires par exemple), c'est-à-dire de massifs de cellules programmées pour donner un organe particulier, qui vont croître jusqu'à former l'organe en question.
La croissance des tissus se fait par allongement des cellules : grâce à la pression engendrée par l'eau (pression de turgescence), les cellules et leur paroi s'allongent.
On peut donc distinguer de manière générale :- Les méristèmes racinaires et secondaires sont uniquement histogènes ;
- Les méristèmes apicaux sont à la fois histogènes et organogènes.
Les différents méristèmes
Il existe différents types de méristèmes liés aux différents rôles que jouent les méristèmes au cours du développement, et surtout au type de tissus spécialisés qu'ils engendrent.
Méristèmes primaires
Les méristèmes primaires sont les premiers à se mettre en place ; ils forment tous les tissus primaires de la plante. Ils permettent une croissance en longueur.
Méristèmes secondaires
Les plantes herbacées (dont les monocotylédones) ne font pas de croissance secondaire. Celle-ci permet la croissance en diamètre de la tige, qui peut aller jusqu'à la formation d'un tronc. Les méristèmes secondaires créent en effet notamment du bois d'une part, du liège d'autre part. Cependant, cette croissance secondaire n'a lieu qu'après la mise en place de la structure primaire de la plante.
Le méristème secondaire principal est le cambium. Présent uniquement chez les dicotylédones, il est aussi appelé assise génératrice libéro-ligneuse (AGLL). Dans les tiges, ce tissu est situé entre le xylème primaire, à différenciation centrifuge dans la tige et centripéte dans la racine d'une part, et d'autre part le phloème primaire, à différenciation centrifuge. Il va ainsi créer des tissus conducteurs secondaires ; les cellules qui le constituent effectuent des divisions radiales de manière à créer :
- vers l'intérieur : du xylème secondaire, dont la particularité est d'être lignifié. Elles constituent ainsi en plus des vaisseaux un tissu de soutien formant le bois.
- vers l'extérieur : du phloème secondaire, que l'on appelle liber (tissu de soutien, mais aussi conducteur : il conduit la sève descendante, dite "sève élaborée" qui va des feuilles jusqu'aux racines).
Les vaisseaux primaires finissent écrasés par la croissance des tissus secondaires.
Le deuxième méristème secondaire est le phellogène ou assise génératrice subéro-phellodermique. Celui-ci, à l'instar du cambium, effectue des divisions radiales créant :- vers l'intérieur : du phelloderme. Ce phelloderme a pour fonction de régénérer le phellogène.
- vers l'extérieur : du suber, plus communément appelé liège. Le liège à pour rôle la protection de la plante et s'ajoute à celle de l'épiderme.
 figure 1 : section de tronc d'un chêne pédonculé (Quercus robur) apparemment âgé de 21 ans
figure 1 : section de tronc d'un chêne pédonculé (Quercus robur) apparemment âgé de 21 ansAu fur et à mesure des années, la plante croît en diamètre en créant progressivement de nouveaux vaisseaux ; les précédents sont remplacés au même rythme. Ils disparaissent dans le cas du phloème, mais le xylème, lui, reste en place et devient un tissu de soutien uniquement, constituant ainsi le bois de cœur (duramen). Ces croissances sont d'autant plus efficaces que la saison le permet ; ainsi dans les zones tempérées peut-on observer des anneaux plus ou moins foncés correspondant à l'intensité de l'activité de la zone cambiale.
Les zones foncées sont donc créées lors de saisons de faible croissance, attribuable à un manque de ressources pour le cambium, comme un manque d'eau ou le passage d'un hiver. Alors que les zones plus claires, créées lors de saisons de forte croissance, correspondent à des vaisseaux d'épaisseur plus importante créés quand le cambium dispose de bonnes ressources.
L'alternance de périodes favorables (printemps/été) et défavorables (automne/hiver) est donc à l'origine de la formation de deux types de bois :
- Le bois initial, constitué en début de saison de végétation, en général de couleur claire ;
- Le bois final, constitué en fin de saison de végétation, de couleur plus sombre. C'est addition de ces 2 couches de bois qui forme le cerne.
Ainsi le décompte des cernes des arbres peut-il donner une idée particulièrement précise de l'âge de la plante : c'est ce principe qui est utilisé dans la dendrochronologie (figure 1).
Organisation du tissu méristématique
Cellules
Les cellules méristématiques ont des fonctions analogues aux cellules souches des animaux : elles sont peu ou pas du tout différenciées et sont capables de continuer la division cellulaire indéfiniment. La grande différence existant entre les cellules souches et les cellules spécialisées, qui n'ont pas le potentiel de différenciation des cellules souches, est beaucoup moins marquée chez les plantes : les cellules différenciées des plantes sont capables de se dédifférencier et de reconstruire de nouveaux tissus ou organes. Le bouturage est un exemple de ces capacités.
Ces cellules sont indifférenciées, ce qui se traduit notamment par un matériel cytoplasmique qui remplit complètement la cellule, bien que les vacuoles soient extrêmement réduites. Le cytoplasme ne contient aucun plaste, même s'ils sont présents sous une forme rudimentaire. Les cellules méristématiques sont de plus étroitement accolées les unes aux autres, séparées par des parois pectocellulosiques très fines et ne présentant aucune cavité intercellulaire.
Structure du méristème
Le méristème apical primaire peut être divisé en couches ou en domaines :
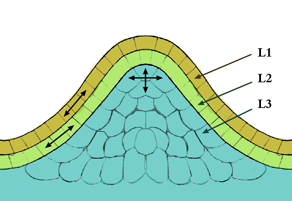
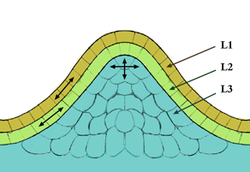 figure 2 : organisation en couches d'un méristème apical
figure 2 : organisation en couches d'un méristème apicalCouches
L'organisation en couche prend en compte les lignées cellulaires qui constituent chacune une couche différente (figure 2).
La couche de cellule la plus externe qui forme l'épiderme est appelée couche L1, et la couche sous-jacente est la couche L2. Le tissu le plus interne, qui est plus qu'une couche de cellules, forme la couche L3, qui formera le parenchyme médullaire. Les couches L1 et L2 qui ont toutes deux une division anticline, forment la tunica
 du méristème, tandis que la couche L3, qui a des divisions à la fois anticline et péricline, forme le corpus du méristème.
du méristème, tandis que la couche L3, qui a des divisions à la fois anticline et péricline, forme le corpus du méristème.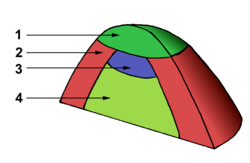 figure 3 : organisation en zones d'un méristème apical
figure 3 : organisation en zones d'un méristème apical
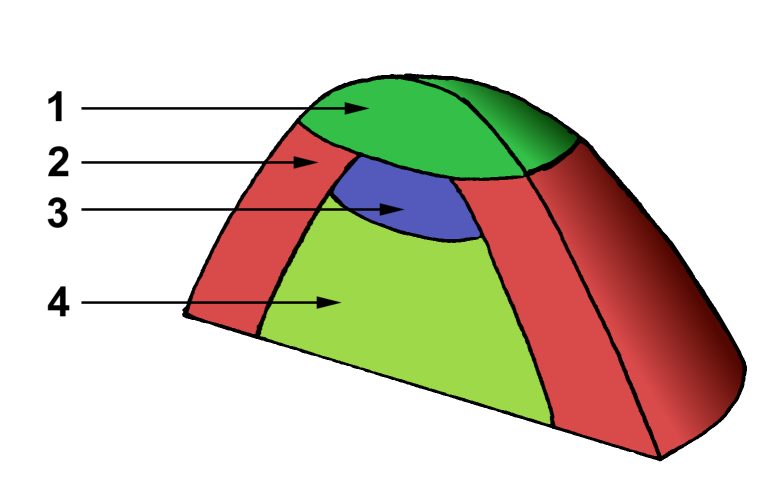
1 - zone centrale
2 - zone périphérique
3 - méristème médullaire
4 - tissus médullairesDomaines
Une autre présentation du méristème consiste à le diviser en domaines (ou zones) selon la fonction des cellules. Certains considèrent ainsi que le méristème apical est plutôt un ensemble de méristèmes spécialisés dans la formation d'un type de cellule spécifique (figure 3).
La première zone, la zone centrale (ZC) (1), est aussi appelée centre quiescent en raison de la faible activité mitotique de ses cellules, qui forment ainsi des cellules totalement indifférenciées. Celles-ci se divisent et migrent vers la zone périphérique (ZP) du méristème (2). Les cellules de cette zone périphérique sont plus actives et forment les primordia foliaires qui constitueront des organes comme les feuilles (rôle organogène). Sous la zone centrale se trouve le méristème médullaire (3), une zone de forte croissance cellulaire qui assure principalement la création des tissus internes : les parenchymes et les tissus conducteurs primaires notamment (4). Son rôle est donc histogène.
La différence principale qui existe entre le méristème apical et le méristème racinaire réside dans le fait que le MAC est à la fois histogène et organogène alors que le MAR est uniquement histogène.
Régulation de l'activité du méristème
L'activité permanente du méristème requiert une régulation précise, sans quoi le méristème pourrait par exemple s'épuiser ou au contraire produire trop de cellules. En outre, les différents méristèmes doivent avoir une identité déterminée (MAC, MAR, méristème floral…). Tout cela est réalisable à l'aide de signaux.
Biotechnologies
Le méristème est un tissu important en biotechnologie. En effet, comme on ne trouve pas de virus dans les méristèmes d'une plante infectée, on peut régénérer une plante saine à partir d'une plante malade en faisant une culture in vitro de méristème.
Ce procédé est en outre très utilisé en horticulture pour produire des plantes d'un même génotype donné. C'est une technique de multiplication asexuée (ou végétative).
Notes et références
Voir aussi
Liens internes
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire Portail de la botanique
Portail de la botanique
Catégories : Histologie végétale | Cellule végétale

Wikimedia Foundation. 2010.