- Donneur universel de sang
-
Groupe sanguin
Un groupe sanguin est une classification de sang reposant sur la présence ou l'absence de substances antigéniques héritées à la surface des globules rouges (hématies). Ces antigènes peuvent être des protéines, des glucides, des glycoprotéines ou des glycolipides, selon le système de groupe sanguin, et certains de ces antigènes sont également présents à la surface d'autres types de cellules de différents tissus.
Les divers groupes sanguins sont regroupés en systèmes. Appartiennent à un même système de groupes sanguins l'ensemble des épitopes ou phénotypes résultant de l'action des divers allèles d'un même gène ou de gènes étroitement liés.
Le sang est un tissu liquide que l’on peut facilement prélever sur un individu sain pour le transfuser à un individu malade. Or, malgré une composition cellulaire identique de ce tissu, il existe une variabilité, ou polymorphisme des divers éléments du sang entre les individus, ce qui rend impossible la transfusion entre certains groupes de personnes. On dit des personnes qui présentent une même caractéristique qu’elles appartiennent au même groupe sanguin. Jusqu’à une époque récente, ces caractéristiques ont été mises en évidence grâce à des anticorps spécifiques d’un épitope, déterminant antigénique reconnu spécifiquement par un anticorps. Ces épitopes, déterminant divers phénotypes, sont génétiquement transmis.
La découverte du système ABO, le premier de ces systèmes, en 1900, par Landsteiner[1] a permis de comprendre pourquoi certaines transfusions sanguines étaient couronnées de succès, alors que d'autres se terminaient tragiquement.
Sommaire
Bases d'immunologie
Les antigènes sont des molécules qui couvrent la surface de toutes les cellules de l'organisme et participent à son identité. Elles sont les cibles des anticorps lorsqu'elles sont identifiées comme étrangères. Mais les antigènes concernent aussi bien des substances extérieures à l'organisme et contre lesquelles réagissent les anticorps : le pollen, la poussière, certains aliments ou médicaments, ou les poils léchés d'animaux.
Les anticorps sont des molécules produites par les lymphocytes B du système immunitaire qui réagissent avec les antigènes n'appartenant pas à l'organisme. Elles attaquent le non-soi. Certains anticorps sont fabriqués « à la demande » (défense contre les bactéries...), d'autres existent naturellement dans l'organisme (ce qui fut découvert avec le système ABO).
Lorsqu'un anticorps se fixe spécifiquement à un antigène situé à la surface des globules rouges, il provoque l'agglutination, parfois l'hémolyse, de ces derniers. Cette agglutination peut être soit immédiate, et c'est ainsi que le système ABO a été découvert, soit « aidée » par une technique d'agglutination artificielle, et c'est ainsi, qu'après les travaux de Coombs, qui a produit et utilisé une antiglobuline, un grand nombre d'anticorps et de systèmes de groupes sanguins ont été découverts.
Classification
Ces différences antigéniques entre les individus définissent les différents groupes sanguins et peuvent porter aussi bien sur les éléments figurés du sang, globules rouges, globules blancs, plaquettes, que sur les protéines circulantes, en particulier les immunoglobulines. Le terme groupe sanguin ayant été appliqué aux seuls groupes connus avant les années 1950, à savoir aux groupes érythrocytaires, et ce terme étant souvent compris et en règle générale utilisé de façon restrictive dans cette acception, ce sont ces derniers qui seront traités dans la suite du présent article. Enfin, historiquement, ce sont les transfusions d'érythrocytes qui ont posé des problèmes cliniques d'incompatibilité, les autres éléments du sang n'étant que peu impliqués dans des accidents transfusionnels immédiats d'origine immunologique.
Nous dirons cependant un mot sur chacun des autres systèmes de groupes, en adressant le lecteur aux articles traitant de façon plus détaillée chacune de ces questions, ou y faisant référence, comme simple polymorphisme ou allotypie.
Groupes sanguins leucocytaires
C'est en travaillant avec des anticorps anti-leucocytes, et en tentant d'identifier des groupes leucocytaires, que Jean Dausset a découvert le système HLA. Il s'agissait en fait des antigènes d'histocompatibilité présents sur toutes les cellules de l'organisme.
Les leucocytes, portent également des antigènes spécifiques, soit aux différentes catégories de lymphocytes, soit aux polynucléaires. Ces derniers portent divers antigènes regroupés en 5 systèmes, HNA1, HNA2, HNA3, HNA4 et HNA5 (HNA pour Human Neutrophil Alloantigen).
Les anticorps dirigés contre les globules blancs, susceptibles d'être contenus dans un plasma transfusé, qu'il s'agisse d'anti HLA ou d'anti HNA, peuvent induire un accident transfusionnel grave, le TRALI (transfusion related acute lung injury) qui consiste en une atteinte œdémateuse pulmonaire.
Voir aussi transfusion sanguine et incompatibilité fœto-maternelle.
Groupes sanguins plaquettaires
Il s'agit des systèmes HPA (Human Platelet Antigens), au nombre de 6 : HPA1, HPA2, HPA3, HPA4, HPA5, et HPA15, tels que définis par le Comité de Nomenclature des Plaquettes, PNC (Platelet Nomenclature Committee) créé en 2003 en association avec l'ISBT et l'ISTH (Société Internationale de Thrombose et Hémostase). Le plus connu de ces systèmes étant le système HPA1, suivi du système HPA5, dont les anticorps anti HPA1-a et HPA5-b sont impliqués respectivement dans 80 % et 10 % des cas d'incompatibilités fœto-maternelles plaquettaires.
Un anticorps dans l'un de ces systèmes entraîne :
- en cas d'incompatibilité fœto-maternelle, une thrombopénie chez le fœtus et le nouveau-né, causant parfois des hémorragies intracrâniennes qui peuvent être graves.
- chez l'adulte, une transfusion inefficace en cas de transfusion de plaquettes incompatibles. Cette transfusion peut être exceptionnellement suivie d'un purpura post-transfusionnel où sont non seulement détruites immédiatement les plaquettes transfusées, mais également, par un mécanisme discuté, les propres plaquettes du patient.
Groupes sériques
Il s'agit des groupes Am, Gm, Km des immunoglobulines A, G, et de la chaîne légère Kappa, ainsi que du groupe ISf (Inhibiteur San Francisco, situé sur la chaîne lourde des IgG1). Ces systèmes, dont le premier a été découvert par Grubb et Laurell, sont déterminés grâce à une antiglobuline, par une technique d'inhibition d'agglutination. La technique est indiquée dans les articles : "discuter:Robin Coombs", et dans le paragraphe allotypie dans "discuter:anticorps".
Groupes érythrocytaires
Il s'agit des premiers groupes sanguins qui ont été découverts (ABO, MNS), et le terme groupes sanguins, utilisé de façon isolée, désigne en règle et de façon restrictive les groupes érythrocytaires, sinon on utilise le terme groupe plaquettaire, leucocytaire, ou sérique.
Groupes sanguins (érythrocytaires)
Les principaux groupes sanguins sont ceux qui définissent les systèmes ABO, Rhésus et Kell, mais il en existe beaucoup d'autres. Ces trois systèmes sont les plus importants, en pratique. Le premier, ABO, car il entraîne un accident transfusionnel immédiat en cas de transfusion incompatible, et de ce fait a été le premier découvert. Le second, Rhésus, car l'immunogénicité de deux de ses antigènes (D, et c, surtout) entraîne très fréquemment des immunisations sources d'accidents ultérieurs et d'incompatibilités fœto-maternelles. Le troisième système, Kell, car l'antigène Kell est très immunogène, moins cependant que l'antigène RH1, D, et donne de ce fait, mais moins fréquemment, les mêmes complications.
La détermination du groupe dans ces trois systèmes en ABO (A, B, AB ou O), en Rhésus (+ ou -), ou en Kell (+ ou -) se base, comme pour tous les systèmes, sur les caractéristiques des antigènes présents à la surface des érythrocytes et, pour le système ABO, sur les anticorps présents dans le sang.
Nous donnons ici la liste des différents systèmes définis et référencés par l'ISBT en août 2008, avec dans l'ordre leur numéro, leur dénomination initiale ou commune, leur dénomination abrégée (symbole) officielle, la nature de l'épitope ou de l'élément qui le porte, la localisation chromosomique. Enfin, dans chaque système, un numéro à 3 chiffres est attribué à chaque spécificité antigénique. Ainsi, dans le ABO quatre spécificités sont référencées : A=001, B=002, AB=003, A1=004. Dans le système MNS nous arrivons au numéro 046, et dans le RH nous dépassons le numéro 050...
Classification des groupes sanguins
Classification des groupes sanguins N° Dénomination initiale ou commune Dénomination abrégée (symbole) officielle Nature de l'épitope ou de l'élément qui le porte Localisation chromosomique 001 ABO ABO ose (N-acétylgalactosamine, galactose) 9q34.2 002 MNS MNS GPA / GPB (glycophorines A et B) 4q31.21 003 P P1 glycolipide 22q11.2-qter 004 Rhésus RH protéines RHD / RHCE 1p36.11 005 Lutheran LU IgSF (apparenté aux immunoglobulines) 19q13.32 006 Kell KEL glycoprotéine 7q34 007 Lewis LE ose (fucose) 19p13.3 008 Duffy FY protéine (ECR ou récepteur de chimiokine, et des Plasmodium vivax et Plasmodium knowlesi) 1q23.2 009 Kidd JK protéine (transporteur d'urée) 18q12.3 010 Diégo DI glycoprotéine (bande 3, AE 1, ou échangeur d'anions) 17q21.31 011 Cartwright YT protéine (AChE, acétylcholinestérase) 7q22.1 012 Xg XG glycoprotéine Xp22.33 013 Scianna SC glycoprotéine 1p34.2 014 Dombrock DO glycoprotéine (fixée à la membrane par le GPI ou glycosyl-phosphatidyl-inositol) 12p12.3 015 Colton CO aquaporine 1 7p14.3 016 Landsteiner-Wiener LW IgSF (apparenté aux immunoglobulines) 19p13.2 017 Chido/Rodgers CH/RG C4A C4B (fractions du complément) 6p21.3 018 Hh H ose (fucose) 19q13.33 019 Kx XK glycoprotéine Xp21.1 020 Gerbich GE GPC / GPD (glycophorines C et D) 2q14.3 021 Cromer CROM glycoprotéine (DAF ou CD55, régulatrice des fractions C3 et C5 du complément, liée à la membrane par un GPI) 1q32.2 022 Knops KN glycoprotéine (CR1 ou CD35, capteur d'immun-complexes) 1q32.2 023 Indian IN glycoprotéine (CD44 fonction d'adhésion ?) 11p13 024 OK OK glycoprotéine (CD147) 19p13.3 025 RAPH MER2 glycoprotéine transmembranaire 11p15.5 026 John Milton Hagen JMH protéine (liée à la membrane par un GPI) 15q24.1 027 Ii I polyoside ramifié (I) / non ramifié (i) 6p24.2 028 Globoside P glycolipide 3q26.1 029 GIL GIL aquaporine 3 9p13.3 030 Rh-associated glycoprotein RHAG 6p21-qter Il existe, n'appartenant pas à ces 30 systèmes, d'autres antigènes présents sur les érythrocytes. Ces antigènes sont classés dans deux séries.
La première, série numérotée 700, regroupe les antigènes de faible incidence, rencontrés chez moins de 1% des individus.
La seconde, série numérotée 901, regroupe les antigènes de haute incidence, rencontrés chez plus de 90% des individus, et très souvent plus de 99% pour la plupart d'entre eux.
Les antigènes rares, qu'ils appartiennent à un système, comme l'antigène Vw du système MNS, (système 002, antigène 009) ou à la série 700 comme les antigènes Peters (700.018) ou Rasmussen (700.040) sont appelés antigènes privés, et les anticorps correspondant anti-privés.
Les antigènes fréquents, qu'ils appartiennent à un système, comme l'antigène RH46, (système 004, antigène 046) ou à la série 901, comme les antigènes Vel (901.001) ou Ata (901.003) sont appelés antigènes publics, et les anticorps correspondant anti-publics.
ABO et RH, modèles de groupes sanguins érythrocytaires
Ces deux systèmes sont les plus importants, tant dans la pratique médicale, que pour leur intérêt historique, car ils ont fourni les bases génétiques, immunologiques pour toutes les études ultérieures des autres systèmes.
Le système ABO
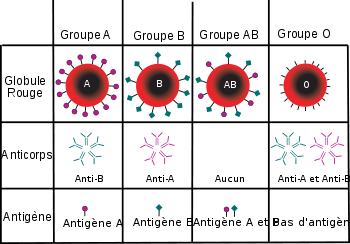
 Article détaillé : Système ABO.
Article détaillé : Système ABO.Découvert en 1900 par Landsteiner, le système ABO permet de classer les différents groupes sanguins selon
- La présence ou non d’antigènes A ou B à la surface des globules rouges.
Ainsi les globules rouges du groupe sanguin A possèdent des antigènes A, ceux du groupe B des antigènes B, ceux du groupe AB des antigènes A et B alors que ceux du groupe O ne contiennent pas d’antigènes de type A ni de type B.
- La présence ou non d'anticorps anti-A ou anti-B dans le sérum. La présence d’antigènes d’un certain type impliquant l’absence d’anticorps de cette spécificité (sous peine de formation d’un complexe anticorps-antigènes !).
- Ces deux recherches, d'antigènes définissant l'épreuve de Beth-Vincent, et d'anticorps définissant l'épreuve de Simonin-Michon sont obligatoires et doivent être concordantes pour établir un groupe sanguin ABO. Une exception toutefois chez le nouveau-né de moins de six mois dont les anticorps ne sont pas bien développés, et chez lequel ne sont donnés que des résultats non définitifs.
Le système Rhésus
Article détaillé : Groupe Rhésus.Ce système, expliquant certains problèmes indépendant du système ABO, accidents transfusionnels et la maladie hémolytique du nouveau-né, fut découvert en 1940 par Landsteiner et Wiener.
Le système Rhésus permet de classer les groupes sanguins selon la présence ou non d’antigène D à la surface des globules rouges (Rhésus est le nom d'un macaque qui a servi à fabriquer le premier sérum-test permettant de tester les groupes Rhésus).
Dans la pratique médicale courante, on distingue les individus rh- qui ne portent pas l'antigène D, ou RH1 dans la nomenclature internationale, sur la surface de leurs hématies et les individus Rh+, qui présentent l'antigène D. En règle générale, les sujet rh- n'ont pas d'anticorps anti-D dans leur plasma. Une transfusion est alors possible sans conséquence immédiate.
Cet anticorps n'apparaît qu'après une transfusion non iso-rhésus (transfusion d'un sang D+ RH1 à un sujet D-) ou une grossesse après la naissance d'un enfant Rh+ chez une femme rh-. On dit alors qu'il s'agit d'un anticorps irrégulier. Dans ce dernier cas, la transfusion d'un sang Rhésus positif D+ entraîne une réaction hémolytique (qui détruit les hématies) par incompatibilité Rhésus.
Ce système de groupe sanguin comporte de nombreux autres antigènes à côté de l'antigène D=RH1. En particulier les antigènes C (RH2), E (RH3), c (RH4) et e (RH5). Certains de ces antigènes peuvent entraîner les mêmes complications transfusionnelles ou fœtales que l'antigène D, en particulier l'antigène c (RH4), qui lui est en règle présent chez un sujet rh négatif
Répartition des groupes
En France, les groupes sanguins se répartissent de la manière suivante (exemple A+ prédomine avec 39%):
Répartition des groupes sanguins dans la population française[2] Rhésus Groupe sanguin Total O A B AB Rh+ 37 % 39 % 7 % 2 % 85 % Rh- 6 % 6 % 2 % 1 % 15 % Total : 43 % 45 % 9 % 3 % 100% Au Canada, en 2006 les groupes sanguins se répartissent de la manière suivante :
Répartition des groupes sanguins dans la population canadienne en 2006 Rhésus Groupe sanguin Total O A B AB Rh+ 39% 36 % 7,5 % 2,5 % 85 % Rh- 7 % 6 % 1,5 % 0,5 % 15 % Total : 46 % 42 % 9 % 3 % 100% Cette répartition peut présenter de sensibles différences selon les origines ethniques :
- chez les aborigènes d'Australie par exemple, on compte 68 % de O et 32 % de A
- chez les Inuits, 86 % de O
- chez les Asiatiques, on compte une plus forte proportion de groupe B.
Exemples de répartitions :
Exemples de répartition dans le monde Groupe sanguin Répartition mondiale O + 38% A + 34% B + 9% O - 7% A - 6% AB + 3% B - 2% AB - 1% Exemples de répartition par type de population Population O A B AB Allemande 41% 43% 11% 5% Belge 44% 45% 8% 3% Britannique 47% 42% 8% 3% Basque 56% 40% 3% 1% Indienne du Pérou 100% 0% 0% 0% Mayas 97% 1% 1% 1% Indienne d'Amérique 96% 4% 0% 0% Oyirad (Russie) 26% 23% 41% 11% Tchouvache (Asie centrale) 30% 29% 33% 7% De ces répartitions, nous pouvons calculer les fréquences géniques dans les diverses populations. Ce type de calculs est valable pour chaque système de groupe sanguin, par application de la loi de Castle-Hardy-Weinberg, article où le système ABO est pris pour exemple.
Compatibilité
La compatibilité entre les groupes sanguins d'un donneur et d'un receveur se pose lors des transfusions sanguines. Une transfusion échouera si des anticorps rencontrent des cellules présentant les antigènes correspondants. Une réaction immunologique (agglutination et hémolyse) se déclencherait alors très rapidement pour détruire ces cellules. Les conséquences peuvent aller d'une transfusion inefficace sans signe clinique, à une réaction clinique légère (angoisse, frisson), grave (état de choc, hémoglobinurie, insuffisance rénale), ou dramatique (Choc, Coagulation intravasculaire disséminée) conduisant au décès.
Elle se pose également en cas de grossesse pour les femmes Rhésus négatif portant un fœtus de rhésus positif. S'il s'agit d'une première grossesse, en général les choses se passent bien si la mère n'a pas été immunisée antérieurement par l'antigène D, RH1. Sinon, du fait que les anticorps peuvent franchir la barrière placentaire, les globules rouges du fœtus sont détruits plus ou moins massivement : c'est la maladie hémolytique du nouveau-né, ou MHNN. Cette maladie peut présenter tous les stades de gravité. Bénigne et n'entraîner qu'un simple ictère (jaunisse) et une anémie passagère, plus importante et nécessiter des transfusions, voire une exsanguino-transfusion à la naissance, majeure demandant un accouchement provoqué ou une césarienne avec exsanguino-transfusion immédiate, gravissime demandant des transfusions in utero pour éviter le décès de l'enfant, voire décès in utero de l'enfant avant toute intervention possible. Ces derniers cas sont devenus très rares depuis la prévention de l'immunisation des femmes par une injection d'anticorps anti-D à l'accouchement d'un enfant Rh Positif. Restent cependant les autres spécificités, dont les plus fréquentes sont l'anti-c (RH4), et l'anti-K1, qui causent également des maladies hémolytiques du nouveau-né.
Présence des anticorps
Type d’anticorps présents selon le groupe sanguin Sang Anticorps présents anti-A anti-B groupe O 
groupe A groupe B groupe AB Dans le système ABO, on retrouve dans le sang de toutes les personnes des anticorps spécifiques des antigènes qu'ils ne possèdent pas sur leurs globules. Ainsi une personne de groupe B développera naturellement des anticorps anti-A et une personne du groupe O développera des anticorps anti-A et des anticorps anti-B. Ces anticorps sont dits réguliers, car présents chez tous les individus, sauf chez le nouveau-né.
Ces anticorps naturels apparaissent, dans le système ABO, dès les premiers mois de la vie. Ce sont des immunoglobulines de classe IgM, agglutinants et agissant à froid : ce sont des anticorps complets.
Dans le système Rhésus, il n'y a pas d'anticorps présents naturellement. Ils n'apparaissent en règle générale qu'après une première sensibilisation, par grossesse ou transfusion (certains tels l'anti-E ou l'anti-Cw pouvant toutefois être "naturels"). Ces anticorps apparus après sensibilisation sont dits irréguliers. Ce sont des immunoglobulines de classe IgG, actifs à 37 °C, et qui ne sont mis en évidence que par des techniques d'agglutination artificielle - techniques à l'antiglobuline ou aux enzymes. Ces anticorps, ne provoquant pas spontanément d'agglutination -mais pouvant provoquer une hémolyse in vivo- sont dits incomplets.
Transfusion de globules rouges
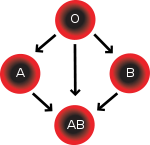 Compatibilité ABO des transfusions de globules rouges:
Compatibilité ABO des transfusions de globules rouges:
Les donneurs O (donneurs universels) peuvent donner aux receveurs O, A, B et AB ; les donneurs A peuvent donner aux receveurs A et AB ; les donneurs B peuvent donner aux receveurs B et AB ; les donneurs AB ne donnent qu'aux receveurs AB.Les globules rouges (ou concentré érythrocytaire) sont extraits de dons de sang. En effet, le sang n'est plus que rarement transfusé dans son intégralité. Il est le plus souvent traité et séparé en ses composants.
Ainsi le concentré érythrocytaire ne contient que peu de plasma, donc peu d’anticorps. Les problèmes d’incompatibilité de la transfusion de plasma (voir ci-dessous) ne sont donc pas posés.
Lors d’une transfusion de globules rouges, il faut veiller à ne pas transmettre au receveur des cellules sanguines sur la surface desquelles se présentent des antigènes que le receveur n’a pas. Un traitement enzymatique, publié en 2007 et actuellement en expérimentation, permet d'éliminer les antigènes A et B des érythrocytes, et de les transformer en hématies de groupe O. Cette possibilité ne deviendra effective que dans quelques années, et, au début, pour des cas très particuliers, sangs dépourvus d'un antigène public présent chez plus de 99 % des donneurs, par exemple.
Les deux tableaux suivants doivent donc être respectés lors d'une transfusion :
Compatibilité système ABO Type du
receveurType de sang recevable O A B AB O A B AB Compatibilité système RH Type du
receveurType de sang recevable Rh+ rh- Rh+ rh- Ainsi, pour le système ABO, et en ne considérant que le groupe rhésus standard (antigène RH1), les sujets AB+ sont considérés comme receveurs universels, et les O- comme donneurs universels de globules rouges.
En ce qui concerne le système RH, cette règle est toujours considérée comme valable en cas d'urgence vitale, et en l'absence de groupe connu. Mais dès que l'on connaît le groupe du patient, il est souhaitable de respecter les autres antigènes de ce système, en particulier l'antigène c (RH 4), afin de ne pas immuniser les femmes jeunes en particulier.
Transfusion de plasma
Le plasma est un des composants du sang. Il est recueilli lors d'un don en plasmaphérèse et peut être utilisé (en France) pour transfusion directe, à condition d'avoir été prélevé chez un homme jamais transfusé (pour éviter la présence d'anticorps anti-HLA ou anti HNA susceptibles d'induire un TRALI, et souvent présents chez les sujets transfusés et les femmes ayant eu des grossesses). Les plasmas extraits de dons de sang total ou par plasmaphérèses chez des femmes, ne peuvent être utilisé (en France) que pour préparer des médicaments dérivés du sang, albumine, immunoglobulines, fibrinogène...ou des réactifs de laboratoire.
Comme le plasma contient des anticorps en fonction de son groupe dans le système ABO, les globules rouges du receveur ne doivent pas présenter les antigènes correspondants. Le plasma de donneurs de groupe AB ne contenant pas d'anticorps convient à tous les receveurs.
Les plasmas contenant un anticorps, le plus souvent naturel dans un autre système (anti-P1, anti-Lewis, anti-M...), hors système Rhésus, ne sont pas utilisés en France. En ce qui concerne le système Rhésus, les plasmas contenant un anti-D (donc issus de femmes immunisées) sont réservés, en France, pour la fabrication des immunoglobulines anti-D, ou d'autres médicaments dérivés du sang. Ces plasmas ne sont donc pas perfusés directement à des malades. Il est clair que dans des pays où seraient utilisés des plasmas contenant un anti-D, le receveur ne pourrait être qu'un sujet rhésus négatif, tout comme le concentré érythrocytaire issu d'une donneuse ayant un anti-D ne peut être transfusé qu'à un patient rh négatif.
En ce qui concerne leur immunogénicité, les plasmas frais congelés, ayant subi une inactivation virale S.D. (solvant-détergent) associée à une filtration ne sont pas immunogènes. Les plasmas qui étaient sécurisés par quarantaine et utilisés jusqu'en septembre 2008 et qui sont remplacés maintenant par le plasma viro-atténué au bleu de méthylène (PVA-BM)ou par amotosalen, technologie Intercept (PVA-IA), peuvent contenir quelques hématies susceptibles, sinon de provoquer une immunisation primaire décelable, du moins de l'induire ou de relancer une immunisation secondaire. D'où la préférence qu'ont certains médecins de respecter pour ces plasmas (PVA-BM ou PVA-IA) une règle de compatibilité Rhésus identique à celle des globules rouges, sans que le bénéfice de cette attitude soit réellement documenté.
Les plasmas viro-atténués par méthode S.D. (solvant-détergent), ayant été filtrés, ne contiennent plus de stromas globulaires et ne peuvent être immunogènes. Pour ces plasmas, nous ne tenons pas compte du groupe Rhésus.
Le donneur et le receveur doivent donc respecter les tableaux suivants :
Compatibilité de groupe ABO Type du
receveurType de plasma recevable O A B AB O A B AB Compatibilité de Rhésus Type du
receveurType de plasma recevable Rh+ rh- Rh+ rh- Ainsi, alors que les personnes de groupe AB sont des receveurs universels de globules rouges dans le système ABO, ils sont donneurs universels de plasma. A contrario, si les personnes de groupe O (ne possédant pas les antigènes A ou B) sont des donneurs universels de globules rouges dans le système ABO, ils sont receveurs universels de plasma, possédant déjà les deux anticorps.[3]
Impasses transfusionnelles
Certains malades posent d'énormes problèmes transfusionnels. Il s'agit en particulier de sujets dépourvus d'un antigène public, sujets Vel négatifs, RH:-46 ou KEL:-2, par exemple. On parle de sujets présentant un groupe sanguin rare ou "phénotype érythrocytaire rare". En cas d'urgence vitale, ces personnes peuvent être transfusées une fois avec du sang classique si elles n'ont pas l'anticorps correspondant à leur groupe sanguin rare, mais ne peuvent plus l'être dès lors qu'elles sont immunisées, sauf par du sang rare identique au leur. De même certaines personnes ayant développé de nombreux anticorps ne peuvent recevoir que du sang d'un phénotype compatible dans les divers systèmes concernés, d'où leur rareté. Ces malades doivent donc dans la mesure du possible participer à un protocole d'auto-transfusion en cas d'intervention chirurgicale programmée, et faire si leur état de santé le permet des dons de sang qui seront conservés congelés à la Banque Nationale des Sangs de Phénotype Rares (BNSPR - Etablissement Français du Sang Ile de France - Créteil). Tous les patients et donneurs présentant un groupe sanguin rare sont suivis par le Centre National de Référence pour les Groupes Sanguins (CNRGS-Paris), département de l'Institut National de la Transfusion Sanguine (INTS - Paris).
Génétique des groupes sanguins
Les groupes sanguins érythrocytaires sont définis grâce aux différences observées entre les individus à la surface des érythrocytes. Il s’agit donc de caractères allotypiques, c'est-à-dire différents d’un individu à l’autre à l’intérieur d’une même espèce.
Ces différences portent sur la présence, l’absence ou l’agencement spatial à la surface des érythrocytes de sucres ou oses (systèmes ABO, P…), ou de protéines (systèmes Rh, Kell…). Autant de différences qui peuvent constituer un épitope antigénique pour quelqu’un qui ne le possède pas.
Ces caractères sont génétiquement transmis selon les lois de Mendel.
Génétique des systèmes ABO et Rhésus
Système ABO
Caractérisé par deux sucres possibles à la surface de l’érythrocyte, soit un galactose (antigène B) soit une N-acétyl-galactosamine (antigène A). Ces sucres sont fixés sur une substance de base, appelée substance H, elle-même osidique. La présence de chacun de ces sucres est due à une enzyme spécifique codée par un gène lui-même spécifique : une variante A pour l’antigène A, B pour l’antigène B. La présence d’une enzyme inactive, due à un codon stop, pour ce gène ne permet pas l’ajout d’un sucre à cette substance de base H, qui reste donc en l’état. Cette enzyme inefficace, sans activité a été appelée "0" (O venant de ohne, sans, en allemand), et a donné le groupe O.
Ainsi le système ABO est caractérisé par un gène dont il existe trois allèles (variantes du gène) A, B, et O.
Ce gène est porté par un autosome (par opposition aux chromosomes sexuels X ou Y). Tout individu possède donc deux copies du gène, l’un venant de son père et l’autre de sa mère, à un même locus, c’est-à-dire à un emplacement défini sur le chromosome. En l’occurrence, pour le système ABO, sur le chromosome 9.
Lorsque le sujet possède à la fois l'allèle A et le B, les deux sucres se trouvent alors sur l’érythrocyte et le sujet est de groupe AB. Lorsqu’il ne possède que 2 allèles O, il sera de groupe O, s’il possède un ou deux A et pas l'allèle B, il sera A, s’il possède un ou deux allèles B et pas le A, il sera B.
Ainsi, un couple de parents, dont la mère est génétiquement A / O, donc de groupe A, et le père B / O, donc de groupe B pourra avoir des enfants de quatre groupes différents. Si chacun des parents transmet son allèle O, l’enfant sera génétiquement O / O, donc de groupe O. Si le père transmet l'allèle O et la mère le A, l’enfant sera A / O, donc de groupe A. Si le père transmet l'allèle B et la mère le O, l’enfant sera B / O, donc de groupe B. Si la mère transmet l'allèle A et le père celui B, l’enfant sera alors A / B, donc de groupe AB.
Système Rhésus
Il s’agit là d’un système de protéines. Deux gènes sont situés à des locus très proches l’un de l’autre sur le chromosome n°1, et sont donc transmis ensemble d’une génération à la suivante. Ces deux gènes résultent d’une duplication d’un gène originel, et synthétisent deux protéines très proches ayant la même structure et la même fonction ; si l’une est absente, l’autre la remplace, ce qui peut expliquer la grande quantité de protéines D chez les sujets ayant une délétion au locus CE (donc de phénotype D--, soit RH:1,-2,-3,-4,-5 en nomenclature internationale), ou les réactivités différentes des hématies selon le nombre de chacun des épitopes présents, lors d'une recherche d'anticorps irréguliers. Au premier locus, locus D, se trouve soit l’allèle D, qui synthétise la protéine Rhésus D définie par la présence de l’antigène D ou RH1, soit un emplacement vide dénommé d, qui ne synthétise rien. Au second locus, locus CE, se trouve un gène qui synthétise une seconde protéine qui ne porte pas l'épitope D. Mais cette seconde protéine présente deux autres épitopes principaux. L’un des ces épitopes définit les antigènes C ou c, le second les antigènes E ou e. La même protéine peut donc avoir quatre combinaisons possibles d'épitopes : ce, Ce, cE, CE.
Ainsi, en combinant l’ensemble des ces possibilités, nous obtenons 8 agencements possibles, ou haplotypes, sur un même chromosome. Quatre des ces agencements comportent le gène D qui définira un sujet Rhésus positif standard. Il s’agit des haplotypes Dce, DCe, DcE, DCE. Quatre des ces agencements ne comportent pas le gène D. Il s’agit des haplotypes dce, dCe, dcE, dCE.
Le même raisonnement que pour les gènes du système ABO s’applique aux haplotypes du système Rhésus. Ainsi deux parents Rhésus positif de génotype D / d, donc hétérozygotes au locus D, pourront avoir un enfant rhésus négatif de génotype d / d.
Génétique des autres systèmes
L'ensemble des autres systèmes de groupes sanguins suit les mêmes lois génétiques. Cependant des particularités spécifiques à chacun des systèmes existent. Système Lewis par exemple, la synthèse de ses antigènes dépendant de deux systèmes génétiques (Lewis, avec ses allèles Le, le et système H avec ses allèles H, h), ou les systèmes Xg ou Kx dont les gènes sont situés sur le chromosome X, et non sur un chromosome autosomal
Anomalies apparentes et filiation
Dans chaque système de groupe sanguin nous pouvons être confrontés à des anomalies apparentes de transmission.
Ainsi, dans le système ABO, nous savons que l’antigène A résulte d’un sucre (ose) fixé par une enzyme sur une substance de base, également osidique, dite substance H. Cette même substance H résulte de l’action d’un gène H, que des très rares sujets ne possèdent pas. Ces sujets sont de génotype h/h, possédant en double dose l’allèle inactif h de H. Ces sujets sont dits de groupe « Bombay », du nom de la localité où cette particularité a été décrite. Ces sujets n’ont donc pas de substance H sur leurs globules rouges, et ont un anticorps anti-H dans leur plasma, ce qui interdit ou rend dangereuse toute transfusion non isogroupe (non "Bombay"). N’ayant pas cette substance H, même si ces sujets "Bombay" possèdent le gène A ou le gène B, les substances A ou B ne pourront être fabriquées, et ces sujets seront en apparence de groupe O. Leurs enfants héritant de ce parent d’un gène h et d’un gène A ou B, et de l’autre parent d’un gène normal H (dans le système Hh) et d’un gène O par exemple (dans le système ABO) pourront à nouveau exprimer le gène A ou B qui leur a été transmis par le premier parent et seront de groupe A ou B normal.
Le même problème peut se poser dans tout autre système où il existe un allèle amorphe, une délétion, une mutation ou un système inhibiteur. Il existe ainsi un rarissime haplotype rhnull dans le système Rhésus. Cet haplotype, qui ne synthétise aucune des deux protéines RH, ni RHD, ni RHCE, est noté RH:---. Supposons un père déterminé comme D+, C+, E-, c-, e+, c'est-à-dire possédant les antigènes D, C, et e, et ne possédant pas les antigènes c et E. Nous en déduisons le génotype vraisemblable de ce père comme étant DCe / DCe, ou DCe / dCe. Or, ce père, uni à une femme de génotype dce / dce, pourra avoir un enfant D-, C-, E-, c+, e+, c'est-à-dire ne possédant pas l’antigène attendu C. Cet enfant sera considéré à tort comme de génotype dce/dce. Nous constatons alors une apparente exclusion de paternité, l'enfant étant supposé avoir reçu un haplotype dce qui n'existe pas chez son père. Or ceci peut être parfaitement expliqué par le génotype DCe / --- de ce père, qui a transmis son haplotype "---" à son enfant dont le génotype réel est dce / --- .
En conclusion, une anomalie apparente de transmission d’un groupe sanguin ne permet en aucune façon à elle seule de conclure à une exclusion de paternité ou de maternité. Une telle conclusion doit s’appuyer sur plusieurs systèmes, et maintenant sur la biologie moléculaire (analyse directe au niveau des chromosomes).
Anomalies, curiosités et pathologies
Antigènes faibles
Dans tous les systèmes nous pouvons voir des antigènes faibles, souvent signalés par un astérisque, ou un f -faible- en indice, ou un w -weak- en indice, sur les résultats du laboratoire, tels les A*, B*, E* ou FY1* ou JK1w. Il est même parfois impossible de mettre ces antigènes en évidence par les techniques habituelles de groupage. Sont alors utilisée des techniques de fixation-élution, voire de biologie moléculaire si besoin.
Il en est ainsi des antigènes A faibles ou B faibles (A3, Ax, Am...B3, Bx...) pour lesquels c'est la faiblesse ou l'absence d'anticorps anti-A ou d'anti-B à l'épreuve de Simonin-Michon qui attire l'attention, et évite que ces groupes ne soient, à tort, étiquetés O. Cet antigène A, ou B, est cependant présent sur les érythrocytes, mais n'est pas mis, ou mal mis en évidence lors de l'épreuve globulaire de Beth-Vincent.
Dans le système RH, les antigènes D faibles sont encore appelés Du.
Tous les autres antigènes de groupe sanguin peuvent être affaiblis, pour diverses raisons, mutation du gène, manque de substrat, gène inhibiteur...Ainsi, comme pour le Rhnull, existe un phénotype Lunull, donc Lu(a-, b-), dû soit à la présence d'un gène amorphe en double dose, cas où aucun antigène LU ne peut être mis en évidence, soit, le plus souvent, à l'action d'un gène inhibiteur. Il s'agit souvent d'un gène autosomique IN(Lu) actif en simple dose, cas où une très faible quantité d'antigène peut être mise en évidence sur les érythrocytes. Ce gène IN(Lu) provoque une forte dépression des antigènes Lutheran, para-Lutheran et AnWj (Anton), et un affaiblissement des antigènes de certains autres systèmes de groupes sanguins, P1, i, Indian, Knops. Existe également, dans quelques familles, un second gène inhibiteur Luthéran nommé XS2, lié au chromosome X, le gène normal étant nommé XS1, dont l'action est légèrement différente de In(Lu) sur les autres antigènes de groupe sanguin.
Certains antigènes de groupe sont connus pour donner, au laboratoire, des réactions très variables d'un individu à l'autre, tel l'antigène P1 chez l'adulte, ou donnent des réactions plus faibles chez le sujet hétérozygote que chez l'homozygote (effet de dose, antigènes M, N, S, Jka...), ou ne sont pas développés à la naissance et apparaissent progressivement en l'espace de deux ou cinq ans, comme les antigènes Lewis ou P1.
Système Lewis
Certaines femmes Le(a-, b+) ou Le(a+, b-), pour trente pour cent d'entre elles, perdent pendant leur grossesse l'antigène Lewis qu'elles possèdent. Elles apparaissent donc comme Le(a-, b-) et développent un anticorps naturel anti-Lewis, anti-Lea, anti-Leb et/ou anti-Lex. Un mois au plus après l'accouchement, cet anticorps a disparu et ces femmes ont retrouvé leur phénotype Lewis normal. Cette perte d'antigène est sans conséquence pour l'érythrocyte, car la substance Lewis est une substance (glycosphingolipide) qui n'appartient pas à la membrane de l'érythrocyte, mais est une substance soluble (que l'on trouve dans le plasma, la salive, les larmes, le lait, le sperme...) adsorbée passivement sur l'érythrocyte.
La substance Lewis n'est pas détectée sur l'érythrocyte du fœtus ni du nouveau-né qui est donc Le(a-, b-) à la naissance. Il apparaît Le(a+, b-) à l'âge d'un mois environ, puis Le(a+, b+) avant de devenir Le(a-, b+) vers l'âge de deux ans si tel doit être son phénotype définitif, lorsqu'il est génétiquement Le (gène Le) et Sécréteur (gène Se), du moins chez les caucasiens. Ceci explique, entre autres raisons, que les anti-Lewis développés chez la mère n'ont aucune conséquence pour le fœtus.
Chimères hématopoïétiques
Lorsque les placentas de deux jumeaux dizygotes fusionnent et permettent une circulation croisée entre les fœtus, chacun d'entre eux possède alors ses propres cellules souches ainsi que celles de son jumeau ou de sa jumelle. Il y a greffe, tolérance immunitaire et les deux lignées cellulaires cohabitent chez le même individu. Dans chaque système nous pouvons observer une double population cellulaire due à la différence de groupe selon l'origine des cellules. Certains érythrocytes appartenant en propre à l'individu seront, par exemple, A, Rh+, K-, d'autres, venant de son jumeau pourront être B, rh-, K+. Parfois, en cas de mort précoce du second embryon, le chimérisme est une découverte fortuite pour l'individu survivant. C'est un cas qui peut poser problème dans les recherches en paternité, voire simuler une exclusion de maternité, le patrimoine génétique des cellules circulantes n'étant pas le même que celui des cellules somatiques ou germinales.
Parfois même il y a fusion précoce entre les deux œufs et il n'en résulte qu'un seul individu, ce qui ne pose pas de problème si les œufs sont du même sexe. Il en résulte un individu unique qui possède donc deux types de cellules, et pas seulement les cellules hématopoïétiques, chaque lignée cellulaire ayant son propre patrimoine génétique.
Les mêmes images de double population se voient régulièrement au laboratoire après transfusion, et dans le cas de greffes de moelle thérapeutiques. Cette double population est visible avant prise totale de la greffe, et réapparaît en cas de rejet.
Perte d'un antigène de groupe sanguin
Dans certaines affections préleucémiques, anémies réfractaires en particulier, certaines lignées d'érythrocytes peuvent perdre, ou plutôt ne plus synthétiser, certains antigènes de groupes sanguins. Par exemple, un sujet connu de groupe AB, peut avoir trois types de globules en circulation, à savoir des globules AB, A, et O, la première lignée n'étant pas atteinte, la seconde ayant perdu une enzyme, et la troisième en ayant perdu deux. Nous avons alors affaire à ce que nous nommons des doubles populations érythrocytaires. Ce constat est parfois un élément étiologique de l'anémie, bien avant les autres éléments cliniques.
Cette perte d'antigènes de groupes sanguins peut s'accompagner d'une perte d'autres enzymes érythrocytaires (adélinate kinase), sans parler des atteintes chromosomiques possibles des autres lignées myéloïdes.
B acquis
Lors d'infections du tube digestif, lors de cancers coliques en particulier, certains germes libèrent une enzyme, une désacétylase, qui transforme la N-acétyl-galactosamine, qui constitue la substance A du groupe ABO, en galactosamine. Certains réactifs anti-B, reconnaissant normalement uniquement le galactose, réagissaient alors comme si le groupe possédait la substance B. Les réactifs maintenant commercialisés sont contrôlés et ne présentent plus, en principe, cette réaction croisée, qui pouvait être source d'erreur entre des mains inexpérimentées, faisant déterminer comme AB un sujet de groupe A. Dès la fin de l'infection, l'anomalie disparaît progressivement.
Autres pathologies
- Protéine Kell non exprimée chez le sujet de phénotype McLeod dont le gène XK, situé en Xp21.1, est génétiquement lié de façon étroite aux gènes de la rétinite pigmentaire (RP), de la granulomatose chronique (CGD) et de la myopathie de Duchenne (DMD), selon la séquence Xpter-DMD-XK-CGD-RP-Xcent. Une délétion à cet endroit expliquant la possible survenue de ces affections chez les sujets de phénotype McLeod, qui présentent par ailleurs une acanthocytose importante et une anémie hémolytique souvent bien compensée.
- RHnull et hémolyse chronique.
- Système FY et résistance au paludisme.
- Agammaglobulinémie et problème de détermination de groupe sanguin ABO -discordance entre les épreuves globulaires et sériques.
- Les taux de vWF et F VIII (facteurs de coagulation von Willebrand et antihémophilique A) sont plus faibles chez les sujets de groupe sanguin O. Les résultats d'un dosage doivent donc être interprétés en fonction du groupe sanguin.
Génétique des populations
Les fréquences géniques des allèles des groupes sanguins, calculées grâce à la loi de Castle-Hardy-Weinberg, ont permis l'essor de la génétique des populations. Grâce à elle, nous pouvons suivre les migrations et les filiations des diverses populations du globe.
Notes et références
- ↑ K. Landsteiner (1900), Zur Kenntnis der antifermentativen, lytischen und agglutinierenden Wirkungen des Blutserums und der Lymphe, Zbl Bakt., 27, 357-362.
- ↑ Source: Bloodbook
- ↑ AFSSaPS : Publications, Bonnes Pratiques, Transfusion
Voir aussi
Bibliographie
- Human blood groups, Geoff Daniels, Blackwell Science Ltd, 1995
- Bases moléculaires des antigènes des groupes sanguins, Jean-Pierre Cartron et Philippe Rouger, Masson, 1998
- The blood group antigen, Marion E. Reid et Christine Lomas-Francis, Facts Book, Elsevier Academic Press, 2° édition, 2004.
- Nomenclature of human platelet antigens, Vox Sanguinis(2003) 85, 240-245
Articles connexes
- Anticorps irréguliers
- Antiglobuline
- Croyance japonaise en l'influence du groupe sanguin sur la personnalité
- Hématologie
- Transfusion sanguine
Liens et documents externes
- (en) International Society for Blood Transfusion (ISBT)
- (en) National Center for Biotechnology Information (NCBI)
- (en) Platelet Nomenclature Commitee (PNC)
- (en) Antigènes plaquettaires, nomenclature, Vox Sanguinis 2003
- (en) Systèmes HNA, TCB juin 2001, p242-247
- (en) Systèmes HNA, National Institute Health (pdf)
- (en) Systèmes érythrocytaires
 Portail de la médecine
Portail de la médecine Portail de la biologie
Portail de la biologie
Catégories : Immuno-hématologie | Groupage sanguin


Wikimedia Foundation. 2010.