- Daspletosaurus
-
 Dasplétosaure
Dasplétosaure
Daspletosaurus sp. Classification Règne Animalia Classe Archosauria Ordre Saurischia Sous-ordre Theropoda Famille Tyrannosauridae Genre Daspletosaurus
Russell, 1970[1]Nom binominal Daspletosaurus torosus
Russell, 1970[1]
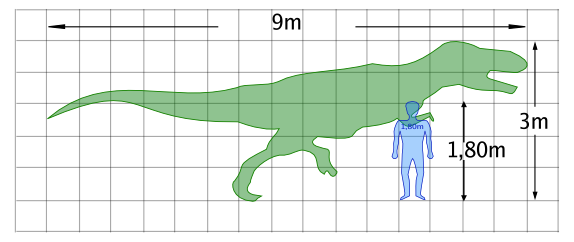
Taille d'un Daspletosaurus sp. comparé à l'homme

D'autres documents multimédia
sont disponibles sur Commons
Daspletosaurus (le Dasplétosaure, qui signifie lézard effrayant) est un genre de tyrannosauridé, moins connu que le célèbre Tyrannosaurus rex, qui vivait dans la partie ouest de l'Amérique du Nord il y a entre 77 et 74 Ma, durant la fin du Crétacé. Les fossiles de la seule espèce décrite (D. torosus) proviennent de la Red Deer River, près de Steveville, dans l'Alberta, bien que des nouvelles espèces possibles de l'Alberta et du Montana sont dans l'attente d'être décrites. En incluant ces espèces, le genre Daspletosaurus est le genre de tyrannosauridé le plus riche en espèces. Comme la plupart des tyrannosauridés connus, c'était un prédateur bipède de plusieurs tonnes, qui possédait des douzaines de larges dents. Daspletosaurus avait également les courts membres antérieurs typiques des tyrannosauridés, bien qu'ils furent plus longs que pour les autres genres.Daspletosaurus était au sommet de la chaîne alimentaire, chassant probablement des gros dinosaures comme le ceratopsidé Centrosaurus et l'hadrosaure Hypacrosaurus. Dans certaines régions, Daspletosaurus a coexisté avec un autre tyrannosauridé, Gorgosaurus, mais il existe des preuves de différenciation des niches écologiques entre ces deux genres. Bien que les fossiles de Daspletosaurus soient plus rares que d'autres tyrannosauridés, les spécimens disponibles permettent des analyses de la biologie de ces animaux, comme le comportement social, le régime alimentaire et développement de l'individu.
Sommaire
Description
 Reconstitution d'un Dasplétosaurus
Reconstitution d'un Dasplétosaurus
Bien qu'étant grand par rapport aux prédateurs modernes, le Daspletosaurus n'était pas le plus grand tyrannosauridé. Les adultes pouvaient atteindre une longueur de 8-9 mètres du museau jusqu'à la queue[2]. Il pesait en moyenne 2,5 tonnes[2][3],[4] mais pouvait atteindre de 1,8[5] à 3,8[6] tonnes.
Le Daspletosaurus avait un crâne massif qui pouvait mesurer plus de 1 mètre de long[2]. Les os étaient solides, et les os nasaux au-dessus du museau étaient fusionnés. Les ouvertures larges dans le crâne réduisaient son poids. Un Daspletosaurus adulte était armé de six douzaines de dents longues mais à section perpendiculaire ovale plutôt qu'en forme de lame. Contrairement à ses autres dents, celles du prémaxillaire au bout de la mâchoire supérieure ont une section perpendiculaire en forme de D. Cette hétérodontie se retrouve chez tous les tyrannosauridés. Les seules caractéristiques du crâne sont la surface externe rugueuse du maxillaire (l'os de la mâchoire supérieure) et les crêtes autour des yeux, sur l'os lacrymal, l'os postorbitaire, et l'os jugal. L'orbite était un grand ovale, intermédiaire entre la forme circulaire de l'orbite de Gorgosaurus et la forme en "trou de serrure" de celle de Tyrannosaurus[7],[8],[9].
Daspletosaurus avait la même morphologie que les autres tyrannosauridés, avec un cou court en forme de S, supportant le crâne massif. Il marchait sur ses deux membres postérieurs, qui se terminaient en pied à quatre orteils, bien que le premier orteil (l'hallux) ne touchait pas le sol. Par rapport aux membres postérieurs, les membres antérieurs étaient extrêmement petits et ne comportaient que deux doigts, bien que Daspletosaurus possède les plus longs membres antérieurs par rapport à la taille du corps, parmi les tyrannosauridés. Une lourde et longue queue servait de contre-poids à la tête et au torse, plaçant le centre de gravité au dessus des hanches[2][9].
Classification et systématique
Cladogrammes des Tyrannosauridae montrant la position de Daspletosaurus
Carr et al. 2005[10] Tyrannosauridae void Albertosaurinae
Tyrannosaurinae void Daspletosaurus
void *Note: Carr et al. utilisent le nom Tyrannosaurus bataar
Currie et al. 2003[11] Tyrannosauridae void Albertosaurinae
Tyrannosaurinae void void Daspletosaurus
void Daspletosaurus appartient à la sous-famille des Tyrannosaurinae parmi la famille des Tyrannosauridae, avec les genres Tarbosaurus, Tyrannosaurus et Alioramus. Les animaux de cette sous-famille sont connus, à l'exception d'Alioramus, pour leur squelette robuste avec proportionnellement des crânes plus gros et de plus longs fémurs que ceux de l'autre sous-famille, les Albertosaurinae[9][11].
Daspletosaurus est généralement considéré comme étant proche parent de Tyrannosaurus rex, ou même un ancêtre direct par anagénèse[12]. Gregory Paul réassigna D. torosus au genre Tyrannosaurus, créant ainsi le Tyrannosaurus torosus[3], mais cela n'a pas été encore généralement accepté[7][9]. Beaucoup de chercheurs pensent que les genres Tarbosaurus et Tyrannosaurus sont des taxons frères, voire le même genre, et Daspletosaurus un parent plus ancestral[9][10]. D'un autre côté, Philip John Currie et des collègues ont trouvé que Daspletosaurus était plus apparenté au Tarbosaurus et à d'autres tyrannosauridés asiatiques comme Alioramus, plutôt qu'au Tyrannosaurus nord-américain[11]. La systématique de Daspletosaurus sera surement plus claire une fois que les espèces seront toutes décrites.
Découverte
Le spécimen type de Daspletosaurus torosus (CMN 8506) est un squelette partiel constitué du crâne, une épaule, un membre antérieur, le pelvis, un fémur, toutes les vertèbres du cou au bassin, et les onze premières vertèbres de la queue. Il a été découvert en 1921 par Charles Mortram Sternberg, qui pensa qu'il appartenait à une espèce de Gorgosaurus. Il fallu attendre 1970 pour que le fossile soit entièrement décrit par Dale Russell, qui en fit le type d'un nouveau genre, Daspletosaurus, du grec δασπλητo-/daspleto- ("effrayant") et σαυρος/sauros ("lézard")[13]. L'espèce type est D. torosus, qui correspond à "musclé", en latin[2]. A part le spécimen type, il n'y a qu'un autre individu bien connu, un squelette complet découvert en 2001. Tous deux proviennent de la formation géologique appelée formation d'Oldman, dans l'ensemble de formations "Judith River Group" d'Alberta. Un spécimen de la formation de Horseshoe Canyon à Alberta a été réassigné à Albertosaurus sarcophagus[8]. La formation d'Oldman s'est déposée pendant l'étage du Campanien moyen, au Crétacé supérieur, entre 77 et 76 Ma[14].
Autres espèces
Deux ou trois espèces supplémentaires ont été attribuées au genre Daspletosaurus au fur et à mesure des années, bien qu'en 2007 aucune de ces espèces n'ait été proprement décrite et ait un nom scientifique. En attendant, elles sont toutes sous le nom Daspletosaurus sp., ce qui n'implique pas qu'elles soient de la même espèce[8][9].
 D'abord attribué à Gorgosaurus libratus, ce crâne (AMNH 5434) fut vendu au Muséum Field à Chicago et est maintenant exposé en tant que FMNH PR308. Il a été réassigné à Daspletosaurus torosus par Thomas Carr en 1999.
D'abord attribué à Gorgosaurus libratus, ce crâne (AMNH 5434) fut vendu au Muséum Field à Chicago et est maintenant exposé en tant que FMNH PR308. Il a été réassigné à Daspletosaurus torosus par Thomas Carr en 1999.Avec l'holotype, Russell désigna un spécimen récolté par Barnum Brown en 1913 comme paratype de D. torosus. Ce spécimen (AMNH 5438) est constitué de fragments de membres antérieurs, du pelvis et de quelques vertèbres associées. Il a été découvert dans la partie supérieure de la formation Oldman[8]. Cette section a été renommée en formation de Dinosaur Park, qui date d'environ 76–74 Ma[14]. En 1914, Brown ramassa un squelette presque complet; quarante ans plus tard l'American Museum of Natural History le vendit au Muséum Field à Chicago. Il a été préparé pour être exposé à Chicago et étiqueté en tant que Albertosaurus libratus pendant de nombreuses années. Mais après plusieurs moulages de caractéristiques du crâne, notamment des dents, le spécimen (FMNH PR308) fut réattribué à Daspletosaurus[7]. Au total, huit individus ont été collectés dans la formation de Dinosaur Park, la plupart dans les limites du parc provincial Dinosaur. Phil Currie pense que les spécimens de Dinosaur Park représentent une nouvelle espèce de Daspletosaurus, qui diffère par certaines caractéristiques du crâne. Des images de ces nouvelles espèces ont été publiées mais elles sont toujours dans l'attente d'un nom et d'une description complète[8].
Un nouveau spécimen de tyrannosaur (OMNH 10131), incluant des fragments de crâne, des côtes et des parties des membres postérieurs, a été rapporté du Nouveau-Mexique en 1990 et attribué à l'ancien genre Aublysodon[15]. Des auteurs ont plus tard réattribué ce spécimen, et quelques autres du Nouveau-Mexique, à une autre espèce de Daspletosaurus[8][9][16]. Cependant, des recherches publiées en 2010 montrent que cette espèce, du Hunter Wash Member de la formation Kirtland, est en fait un tyrannosauroïdé plus primitif, et a été classé dans le genre Bistahieversor[17]. Il y a actuellement des divergences quant à l'âge de la formation Kirtland, que certains attribuent au Campanien supérieur[18], tandis que d'autres le pensent plus récent, de l'étage du Maastrichtien[19].
En 1992, Jack Horner et des collègues publièrent un rapport préliminaire sur un tyrannosauridé du Campanien supérieur, de la formation de Two Medicine dans le Montana, qui a été interprété comme une espèce intermédiaire entre Daspletosaurus et Tyrannosaurus[12]. Un autre squelette partiel fut rapporté de la partie supérieure de cette formation en 2001, avec les restes d'un hadrosaure juvénile dans sa cavité abdominale. Il fut attribué à Daspletosaurus mais à aucune espèce en particulier[20]. Les restes d'au moins trois autres Daspletosaurus ont été décrits dans un niveau riche en os de la formation Two Medicine[21]. Ces spécimens n'ont pas été décrits en détail, mais Currie pense que tout le matériel de la formation Two Medicine représente une troisième espèce de Daspletosaurus[8].
Paléobiologie
Coexistence avec Gorgosaurus
A la fin du Campanien, en Amérique du Nord, Daspletosaurus était contemporain du tyrannosauridé de la sous-famille des Albertosaurinae Gorgosaurus. C'est une des rares coexistences de genres de tyrannosaures connues. Dans les guildes de prédateurs modernes, des prédateurs de tailles similaires sont séparés dans des niches écologiques différentes par des caractères anatomiques, comportementaux ou géographiques différents qui limitent la compétition[22]. Plusieurs études ont tenté d'expliquer la différenciation des niches écologiques de Daspletosaurus et Gorgosaurus.
Dale Russell émit l'hypothèse que Gorgosaurus, moins solide et plus commun, devait chasser les hadrosaures, tandis que Daspletosaurus, plus robuste et moins commun, devait s'être spécialisé dans la chasse des cératopsidés, moins abondants mais avec une meilleure défense, qui les aurait rendu plus difficiles à chasser[2]. Cependant, un spécimen de Daspletosaurus (OTM 200) de la formation Two Medicine contient les restes d'un hadrosaure juvénile dans sa cavité abdominale[20]. Les mâchoires plus grandes et plus larges des tyrannosaurinés comme Daspletosaurus sont plus fortes que celles des albertosaurinés comme Gorgosaurus, bien que la force des dents des deux groupes soit similaire. Cela semble montrer une différence dans le mode d'alimentation ou le régime alimentaire[23].
D'autres auteurs ont suggéré que la compétition était limitée par une séparation géographique. A la différence d'autres groupes de dinosaures, il semble qu'il n'y ait pas de corrélation avec la distance de la mer. Ni Daspletosaurus ni Gorgosaurus était plus abondant que l'autre à des altitudes hautes ou basses[22]. Cependant, bien qu'il y ait un chevauchement, Gorgosaurus semble plus commun vers le nord, et les espèces de Daspletosaurus plus abondantes dans le sud. Ce modèle se retrouve dans d'autres groupes de dinosaures. Les cératopsiens cératopsinés et les hadrosaures hadrosaurinés sont aussi plus communs dans la formation Two Medicine et dans le sud-ouest de l'Amérique du Nord pendant le Campanien. Thomas Holtz pense que ces modèles peuvent montrer des préférences écologiques partagées entre les tyrannosaurinés, cératopsinés et hadrosaurinés. Holtz note qu'à la fin du Maastrichtien, les tyrannosaurinés comme Tyrannosaurus rex, les hadrosaurinés et les cératopsinés comme Triceratops étaient répandus à travers l'ouest de l'Amérique du Nord, alors que les albertosaurinés et les centrosaurinés étaient éteints, et les lambeosaurinés étaient très rares[9].
Comportement social
 Squelette exposé au Muséum Field à Chicago
Squelette exposé au Muséum Field à ChicagoUn jeune spécimen des espèces de Daspletosaurus de la formation de Dinosaur Park (TMP 94.143.1) présente des traces de morsure sur le crâne, infligées par un autre tyrannosaure. Les marques ont guéri, ce qui indique que l'animal a survécu à la morsure. Un Daspletosaurus adulte de cette même formation (TMP 85.62.1) montre des marques similaires de morsure d'un tyrannosaure, ce qui montre que ces attaques n'étaient pas dirigées seulement contre les jeunes. Bien que ces morsures puissent être attribuées à d'autres espèces, l’agression intraspécifique, dont la morsure faciale, est très commune parmi les prédateurs. Ces morsures à la face se retrouvent chez d'autres tyrannosaures comme Gorgosaurus et Tyrannosaurus, ainsi que dans d'autres groupes de théropodes comme Sinraptor et Saurornitholestes. Darren Tanke et Phil Currie émirent l'hypothèse que les morsures sont dues à la compétition intraspécifique pour le territoire ou les ressources, ou pour la dominance dans un groupe social[24].
Des preuves que Daspletosaurus vivait en groupe sociaux ont été trouvées dans un niveau riche en os de la formation Two Medicine dans le Montana. Il contenait les restes de trois Daspletosaurus, dont un adulte, un juvénile, et un autre individu d'une taille intermédiaire. Au moins cinq hadrosaures sont préservés au même endroit. La géologie de la région prouve que les restes n'ont pas été charriés pas un cours d'eau mais que tous ces animaux ont été ensevelis au même endroit simultanément. Les restes des hadrosaures sont dispersés et portent de nombreuses marques de dents de tyrannosaur, ce qui indique que les Daspletosaurus étaient en train de se nourrir des hadrosaures au moment de la mort. La cause de la mort est inconnue. Currie imagine que les dasplétosaures formaient une meute, bien que cela ne puisse pas être affirmé de façon certaine[21]. D'autres scientifiques restent sceptiques quant aux preuves des groupes sociaux de Daspletosaurus et d'autres grands théropodes[25]. Brian Roach et Daniel Brinkman ont suggéré que les interactions sociales de Daspletosaurus devaient ressembler à celles du dragon de Komodo, où des individus non-coopératifs se massent sur des carcasses, s'attaquant souvent parfois même allant jusqu'au cannibalisme[26].
Développement
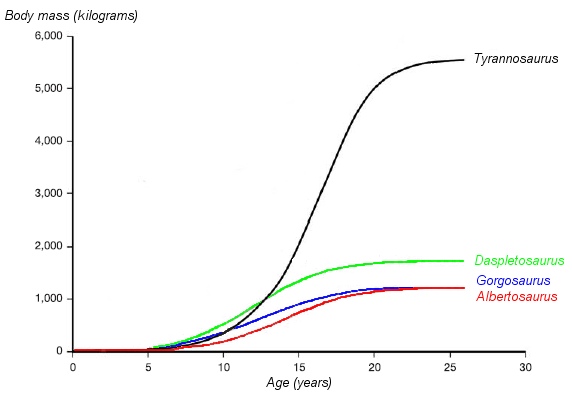
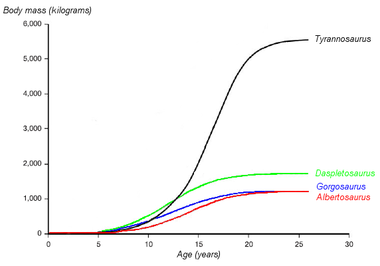 Graphique montrant les courbes de croissance hypothétiques (masse en fonction de l'âge) de quatre tyrannosauridés. Daspletosaurus est représenté en vert. Basé sur Erickson et al. 2004.
Graphique montrant les courbes de croissance hypothétiques (masse en fonction de l'âge) de quatre tyrannosauridés. Daspletosaurus est représenté en vert. Basé sur Erickson et al. 2004.Les paléontologues Gregory Erickson et des collègues ont étudié la croissance et le développement des tyrannosauridés. Des analyses de l'histologie des os peut déterminer l'âge d'un spécimen à sa mort. Les taux de croissance peuvent être examinés quand les âges de différents individus sont comparés à leurs masses sur un graph. Erickson a montré qu'après une longue période en tant que juvéniles, les tyrannosaurs subissent des poussées de croissance importantes pendant environ quatre ans, vers la moitié de leur vie. La phase de croissance rapide se termine par la maturité sexuelle. Erickson a seulement examiné les Daspletosaurus de la formation de Dinosaur Park, mais ces spécimens montrent le même modèle. Comparé aux albertosaurinés, les Daspletosaurus montrent un taux de croissance plus rapide, dû à sa taille adulte plus importante. Le taux de croissance le plus important pour les Daspletosaurus est 180 kilogrammes par an, basé sur une estimation de la masse de 1800 kilogrammes pour un adulte. D'autres auteurs ont suggéré des poids plus importants pour les Daspletosaurus adultes; ce qui changerait la magnitude du taux de croissance mais pas le modèle dans son ensemble[5].
Par la tabulation du nombre de spécimens de chaque groupe d'âge, Erickson et ses collègues purent tirer des conclusions sur l'histoire d'une population d'Albertosaurus. Leur analyses ont montré que bien que les juvéniles soient rares dans l'enregistrement fossile, les individus subadultes en phase de croissance rapide et les adultes sont beaucoup plus courant. Cela pourrait être des biais dus à la préservation ou à la récolte, mais Erickson émit l'hypothèse que la différence était due à une faible mortalité des individus juvéniles au delà d'une certaine taille, ce qui s'observe chez des grands mammifères actuels comme l'éléphant. Cette faible mortalité peut résulter d'un manque de prédation, puisque les tyrannosaures surpassaient tous les prédateurs contemporains en taille dès leur deuxième année. Les paléontologues n'ont pas trouvé assez de restes de Daspletosaurus pour faire une analyse similaire, mais Erickson note que la même tendance générale semble s'appliquer[27].
Paléoécologie
Tous les fossiles de Daspletosaurus ont été trouvés dans des formations datant du Campanien moyen à supérieur, dans le Crétacé supérieur, il y a entre 77 et 74 millions d'années. Depuis le milieu du Crétacé, l'Amérique du Nord a été séparée en deux par la voie maritime intérieure de l'Ouest, avec une grande part du Montana et de l'Alberta sous la surface. Cependant, la surrection des Montagnes Rocheuses pendant l'orogenèse laramienne à l'ouest, qui commença pendant la période du Daspletosaurus, força la mer à se retirer vers l'est et le sud. Des rivières s'écoulèrent depuis les montagnes jusque la mer, charriant du sédiment qui forma la formation Two Medicine, le groupe de formations de la Judith River, et d'autres formations sédimentaires de la région. Il y a environ 73 millions d'années, la mer commença à revenir vers l'ouest et le nord, et toute la région fut couverte par la mer de Bearpaw, qui déposa dans l'ouest des Etat-Unis et du Canada les massifs schistes de Bearpaw[28],[29],[30].
Daspletosaurus vivait dans une vaste plaine d’inondation près de la côte ouest de la mer intérieure. De grandes rivières coulaient dans la plaine, l'inondant occasionnellement, recouvrant la région de nouveau sédiment. Quand l'eau était abondante, la région pouvait accueillir beaucoup de plantes et de vie animale, mais des sécheresses périodiques ont aussi frappé la région, causant des extinctions en grand nombre dont les fossiles sont préservés dans des niveaux très fossilifères des sédiments de la formation Two Medicine et des formations de la Judith River, incluant le niveau à Daspletosaurus[31]. Des conditions similaires existent actuellement dans l'Est de l'Afrique[32]. Des éruptions volcaniques à l'ouest recouvrent périodiquement la région de cendres, causant une forte mortalité, tout en enrichissant le sol pour la pousse de futures plantes. Ce sont ces niveaux à cendres qui permettent une datation radiométrique précise. La fluctuation des niveaux marins a resulté en une variété d'autres environnements à des endroits et des moments différents dans les formations du groupe de la Judith River, incluant des habitats marins profonds et côtiers, des littoraux, des deltas et des lagons, en plus des plaines d'inondation[29]. la formation The Two Medicine fut déposée à plus en hauteur, et plus loin dans les terres que les deux autres formations[30].
 Reconstitution d'un Daspletosaurus torosus mangeant un cératopsien
Reconstitution d'un Daspletosaurus torosus mangeant un cératopsienL'enregistrement excellent de vertébrés fossiles de Two Medicine et Judith River résulte de la combinaison d'une vie animale abondante, des catastrophes naturelles périodiques, et le dépôt de grandes quantités de sédiment. Beaucoup de poissons d'eau douce et d'estuaire sont représentés, dont des requins, des raies, des esturgeons, et d'autres. Le groupe de formations de la Judith River préserve les restes de beaucoup d'amphibiens et reptiles aquatiques, dont des grenouilles, des salamandres, des tortues, des Champsosaurus et des crocodiliens. Des lézards terrestres, dont des Teiidae, des Scincidae, des varans et des Anguidae, ont également été découverts. Des ptérosaures Azhdarchidae, et des neornithes comme Apatornis survolaient cette région, tandis que Avisaurus, un énantiornithe, et plusieurs variétés de multituberculés, marsupiaux et mammifères placentaires courraient entre les pattes de Daspletosaurus et d'autres dinosaures[29].
Dans la formation d'Oldman, Daspletosaurus torosus a pu chasser des hadrosaures comme Brachylophosaurus et Hypacrosaurus, des ornithopodes comme Orodromeus, des cératopsiens comme Centrosaurus, des pachycephalosaures, des ornithomimidés, des thérizinosaures et probablement des ankylosaures. Les autres prédateurs étaient composés de Troodontidae, d'oviraptorosaures, du dromaeosauridé Saurornitholestes et peut-être d'un tyrannosaure albertosauriné (le genre est inconnu pour le moment). Les formations de Dinosaur Park et Two Medicine ont des faunes comparables à celle d'Oldman, avec la particularité de Dinosaur Park de préserver une gamme unique de dinosaures[29]. L'albertosauriné Gorgosaurus vivait avec des espèces de Daspletosaurus dans les environnements de la formation de Dinosaur Park et du haut de la formation Two Medicine[22]. Les jeunes tyrannosaures devaient remplir les niches écologiques entre les tyrannosaures adultes et les théropodes plus petits[2][9][23][33]. Un dentaire de Saurornitholestes a été découvert dans la formation de Dinosaur Park. Il porte des marques de dents d'un jeune tyrannosaure, peut-être Daspletosaurus[34].
Annexes
Articles connexes
- Voir Liste des dinosaures
Liens externes
Références taxinomiques
- Référence The Paleobiology database : Daspletosaurus, Russell (1970) (en)
Notes et références
- D. A. Russell. 1970. Tyrannosaurs from the Late Cretaceous of western Canada. National Museum of Natural Sciences, Publications in Paleontology 1:1-34
- Dale A. Russell, « Tyrannosaurs from the Late Cretaceous of western Canada », dans National Museum of Natural Sciences Publications in Paleontology, vol. 1, 1970, p. 1–34
- (en) Gregory S. Paul, Predatory Dinosaurs of the World, New York, Simon & Schuster, 1988 (ISBN 978-0-671-61946-6), p. 464pp
- Per Christiansen, « Mass prediction in theropod dinosaurs », dans Historical Biology, vol. 16, no 2-4, 2004, p. 85–92 [lien DOI]
- GM Erickson, Gregory M., « Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs », dans Nature, vol. 430, no 7001, 2004, p. 772–775 [lien PMID, lien DOI]
- François Therrien, « My theropod is bigger than yours... or not: estimating body size from skull length in theropods », dans Journal of Vertebrate Paleontology, vol. 27, no 1, 2007, p. 108–115 [texte intégral, lien DOI]
- Thomas D. Carr, « Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Coelurosauria) », dans Journal of Vertebrate Paleontology, vol. 19, no 3, 1999, p. 497–520 [lien DOI]
- Philip J. Currie, « Cranial anatomy of tyrannosaurids from the Late Cretaceous of Alberta », dans Acta Palaeontologica Polonica, vol. 48, no 2, 2003, p. 191–226 [texte intégral [PDF]]
- (en) Thomas R. Holtz, The Dinosauria, Berkeley, University of California Press, 2004 (ISBN 0-520-24209-2), « Tyrannosauroidea », p. 111–136
- Thomas D. Carr, « A new genus and species of tyrannosauroid from the Late Cretaceous (middle Campanian) Demopolis Formation of Alabama », dans Journal of Vertebrate Paleontology, vol. 25, no 1, 2005, p. 119–143 [texte intégral, lien DOI]
- Philip J. Currie, « Skull structure and evolution in tyrannosaurid phylogeny », dans Acta Palaeontologica Polonica, vol. 48, no 2, 2003, p. 227–234 [texte intégral [PDF]]
- John R. Horner, « Marine transgressions and the evolution of Cretaceous dinosaurs », dans Nature, vol. 358, no 6381, 1992, p. 59–61 [lien DOI]
- (en) Henry G. Liddell, & Scott, Robert, Greek-English Lexicon, Oxford, Oxford University Press,, 1980, Abridgede éd. (ISBN 978-0-19-910207-5) (LCCN 33031054)
- David A. Eberth, « Tectonic, stratigraphic, and sedimentologic significance of a regional discontinuity in the Upper Judith River Group (Belly River wedge) of southern Alberta, Saskatchewan, and northern Montana », dans Canadian Journal of Earth Sciences, vol. 30, 1993, p. 174–200 [lien DOI]
- Thomas M. Lehman, « A partial skeleton of the tyrannosaurid dinosaur Aublysodon from the Upper Cretaceous of New Mexico », dans Journal of Paleontology, vol. 64, no 6, 1990, p. 1026–1032
- (en) Thomas D. Carr, Dinosaurs of New Mexico, 2000, « A review of Tyrannosauridae (Dinosauria: Coelurosauria) from New Mexico », p. 113–146
- T.D. Carr, « Bistahieversor sealeyi, gen. et sp. nov., a new tyrannosauroid from New Mexico and the origin of deep snouts in Tyrannosauroidea », dans Journal of Vertebrate Paleontology, vol. 30, no 1, 2010, p. 1–16 [lien DOI]
- (en) Robert M. Sullivan, Late Cretaceous vertebrates from the Western Interior, 2006, « The Kirtlandian land-vertebrate "age" - faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of North America », p. 7–29
- (en) Michael J. Ryan, Encyclopedia of Dinosaurs, San Diego, Academic Press, 1997 (ISBN 0-12-226810-5), « Kirtland Formation », p. 390–391
- David J. Varricchio, « Gut contents from a Cretaceous tyrannosaurid: implications for theropod dinosaur digestive tracts », dans Journal of Paleontology, vol. 75, no 2, 2001, p. 401–406 [lien DOI] DOI: 10.1666/0022-3360(2001)075<0401:GCFACT>2.0.CO;2
- (en) Philip J. Currie, The Carnivorous Dinosaurs, Bloomington, Indiana University Press, 2005 (ISBN 978-0-253-34539-4), « An unusual multi-individual tyrannosaurid bonebed in the Two Medicine Formation (Late Cretaceous, Campanian) of Montana (USA) », p. 313–324
- James O. Farlow, « Body size overlap, habitat partitioning and living space requirements of terrestrial vertebrate predators: implications for the paleoecology of large theropod dinosaurs », dans Historical Biology, vol. 16, no 1, 2002, p. 21–40 [lien DOI]
- Eric Snively, « Fused and vaulted nasals of tyrannosaurid dinosaurs: implications for cranial strength and feeding mechanics », dans Acta Palaeontologica Polonica, vol. 51, no 3, 2006, p. 435–454 [texte intégral [PDF]]
- Darren H. Tanke, « Head-biting behavior in theropod dinosaurs: paleopathological evidence », dans Gaia, vol. 15, 1998, p. 167–184 [texte intégral [PDF]] [not printed until 2000]
- David A. Eberth, « Were large theropods gregarious? », dans Journal of Vertebrate Paleontology, vol. 21, no (Supplement to 3 - Abstracts of Papers, 61st Annual Meeting of the Society of Vertebrate Paleontology), 2001, p. 46A[published abstract only]
- Brian T. Roach, « A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs », dans Bulletin of the Peabody Museum of Natural History, vol. 48, no 1, 2007, p. 103–138 [texte intégral, lien DOI]
- Gregory M. Erickson, « Tyrannosaur life tables: an example of nonavian dinosaur population biology », dans Science, vol. 313, no 5784, 2006, p. 213–217 [lien PMID, lien DOI]
- Joseph M. English, « The Laramide Orogeny: what were the driving forces? », dans International Geology Review, vol. 46, no 9, 2004, p. 833–838 [texte intégral, lien DOI]
- (en) David A. Eberth, Encyclopedia of Dinosaurs, San Diego, Academic Press, 1997 (ISBN 0-12-226810-5), « Judith River Wedge », p. 199–204
- (en) Raymond R. Rogers, Encyclopedia of Dinosaurs, San Diego, Academic Press, 1997 (ISBN 0-12-226810-5), « Two Medicine Formation », p. 199–204
- Raymond R. Rogers, « Taphonomy of three dinosaur bonebeds in the Upper Cretaceous Two Medicine Formation of northwestern Montana: evidence for drought-induced mortality », dans PALAIOS, PALAIOS, Vol. 5, No. 5, vol. 5, no 5, 1990, p. 394–413 [texte intégral, lien DOI]
- Howard J. Falcon-Lang, « Growth interruptions in silicified conifer woods from the Upper Cretaceous Two Medicine Formation, Montana, USA: implications for palaeoclimate and dinosaur palaeoecology », dans Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 199, 2003, p. 299–314 [lien DOI]
- James O. Farlow, « Speculations about the diet and foraging behavior of large carnivorous dinosaurs », dans American Midland Naturalist, American Midland Naturalist, Vol. 95, No. 1, vol. 95, no 1, 1976, p. 186–191 [lien DOI]
- Jacobsen, A.R. 2001. Tooth-marked small theropod bone: An extremely rare trace. p. 58-63. In: Mesozioc Vertebrate Life. Ed.s Tanke, D. H., Carpenter, K., Skrepnick, M. W. Indiana University Press.
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Daspletosaurus » (voir la liste des auteurs)
 Portail de la paléontologie
Portail de la paléontologie Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant Portail des dinosaures
Portail des dinosaures Portail de la zoologie
Portail de la zoologie
Catégories :- Theropoda
- Dinosaure du Crétacé
- Formation Oldman
- Formation Dinosaur Park





Wikimedia Foundation. 2010.