- Mycorises
-
Mycorhize
La mycorhize (du grec myco ; champignon et rhiza ; racine) est le résultat de l'association symbiotique entre des champignons et les racines des plantes.
 Fructification (ou carpophore) de l'Amanite tue-mouches ; c'est la partie visible du champignon, qui est en réalité essentiellement souterrain
Fructification (ou carpophore) de l'Amanite tue-mouches ; c'est la partie visible du champignon, qui est en réalité essentiellement souterrain
 Culture du champignon mycorrhizien associé à Woollsia pungens (éricacées).
Culture du champignon mycorrhizien associé à Woollsia pungens (éricacées). L'orge, comme beaucoup d'autres graminées, croît mieux grâce à des endomycorrhizes
L'orge, comme beaucoup d'autres graminées, croît mieux grâce à des endomycorrhizesDans cette association, les hyphes d’un champignon colonisent les racines d’une plante. Les hyphes sont l'organe principal des champignons (rappelons ici que ce que l'on appelle couramment "champignon" (pied + chapeau) que l'on cueille, n'est qu'un organe éphémère du champignon, le carpophore, où se déroule la reproduction sexuée). Les hyphes se présentent comme de fins filaments, capables d'explorer un très grand volume de sol (1000 m de filaments mycéliens pour 1 m de racine).
Histoire de la mycorhization et génétique
Les études génomiques récentes laissent penser que la mycorhization arbusculaire, la plus courante, associant des végétaux supérieurs et des champignons du groupe des Glomus (courants sur toute la planète) date d'au moins 400 millions d'années. Elle concerne aujourd'hui 80% des plantes terrestres et est associée à un programme génétique qui semble particulièrement stable, peut-être inchangé depuis des millions d'années, qu'on retrouve notamment chez beaucoup de légumineuses telles que haricot, pois, soja qui produisent le même type de nodosités symbiotiques fixatrices d'azote gazeux. L'étude du séquençage génétique déterminant la nature intra- ou extracellulaire des échanges de matière (qui conditionne leur quantité et leur qualité) a permis d'éclairer les mécanismes d'interactions en jeu.
Les fabricants d'OGM pourraient chercher à commercialiser les séquences codantes permettant cette symbiose, qui pourrait permettre à d'autres cultures de se passer d'engrais chimiques azotés[1].
Types de mycorhization
On distingue deux types principaux de mycorhizes, définis par les relations physio-anatomiques entre les deux partenaires :
- Les ectomycorhizes : ce sont des associations fréquentes entre les arbres des régions tempérées (comme les Fagacées, les Pinacées ou les Bétulacées) et des Ascomycètes ou des Basidiomycètes.
Le champignon s’associe d’abord aux racines fines à croissance déterminée, dépourvues de poils absorbants. Puis, il enveloppe la racine d’un manteau d’hyphes, le manchon mycorhizien. D'autres hyphes croissent entre les cellules dans la partie externe du parenchyme cortical, formant ainsi l'interface symbiotique ou « réseau de Hartig ». La symbiose modifie la physionomie de la racine mycorhizée : elle se renfle, cesse de croître et peut se ramifier de façon abondante. La coiffe et le méristème apical sont alors réduits.
- Les endomycorhizes : il en existe deux types :
- - les endomycorhizes à vésicules et arbuscules (AM) ;
- - les endomychorizes à pelotons.
Dans les deux cas, les hyphes pénètrent dans la paroi des cellules (de l’écorce). La paroi des hyphes est donc en contact avec la membrane plasmique de la cellule racinaire, sans la traverser. La surface de contact peut être augmentée par la formation de ramification. Les racines ne sont pas déformées.
-
- Les endomycorhizes à arbuscules (www.kent.ac.uk/bio/beg): c’est le cas le plus répandu. Elles impliquent des champignons Gloméromycètes ayant perdu la reproduction sexuée. Les hyphes s’étendent dans le parenchyme cortical de la racine, formant des vésicules contenant des réserves, et des arbuscules.
- Les endomychorizes à pelotons intracellulaires : les hyphes forment des amas dans les cellules corticales. Elles impliquent des basidiomycètes, en symbiose avec les Orchidacées.
- Les endomycorhizes éricoïdes : les hyphes forment des pelotons dans des racines transitoires de faible diamètre. Elles impliquent des Ascomycètes ou Basidomycètes (en symbiose avec les Ericales).
- Les ectendomychorizes : elles sont aussi appelées mycorhizes de type arbutoïde. Le champignon forme des pelotons intracellulaires et un manteau autour de la racine. C’est le cas chez les Ericales.
On trouve également des ectendomychorizes monotropoïdes, chez les Ericales non chlorophylliennes. Les hyphes forment des pelotons dans les cellules superficielles de la racine.
Dans cette symbiose, le réseau dense et étendu des hyphes des champignons mycorhiziens aide la plante à obtenir des sels minéraux présents dans le sol (principalement l'eau, le phosphore et l'azote), tout en la protégeant d’attaques d’autres organismes pathogènes. Plusieurs études ont démontré que sans association avec un champignon symbiotique, la plante poussera plus lentement (voire pas du tout, comme dans le cas du Pin noir sur sol calcaire) et sera beaucoup plus susceptible d’être la victime d’une infection. En retour, le champignon bénéficie de la photosynthèse de la plante sous forme de matière organique riche en énergie (sucres) essentielle à sa survie. La qualité de l'humus en est améliorée, au bénéfice d'autres espèces et du maintien ou de la constitution et amélioration du sol.
Quelques plantes de la famille des Gymnospermes comme les Podocarpaceae, les Araucariaceae et les Phyllocladaceae, sont connues pour présenter des renflements racinaires envahis par des champignons endomycorhiziens, appelées myconodules ou pseudonodules
Ectomycorhizes
Les ectomycorhizes ne pénètrent pas à l’intérieur de la paroi cellulaire de la plante, mais entourent simplement les racines, formant un manteau de mycélium et un réseau entre les parois des cellules de la racine.
Ils sont typiquement formés par un champignon de la division des Basidiomycètes, Ascomycètes, ou des Zygomycètes et les racines d’une plante ligneuse.
Les plantes faisant partie de l’ordre des Ericales et de la famille des Orchidacées forment aussi des mycorhizes. Ces associations, plus rares, sont cependant différentes des deux formes précédemment mentionnées.
Endomycorhizes
Les endomycorhizes (aussi appelés champignons mycorhiziens à vésicules et arbuscules), tirent leur nom de ce qu’ils forment comme structures en forme d’arbuscules ou de vésicules à l’intérieur même de la cellule de la plante. S'ils traversent bien la paroi, ils ne pénètrent cependant pas la membrane plasmique de la cellule végétale, se contentant de provoquer une invagination de la membrane de celle-ci. Cela a pour effet d’accroître la surface de contact entre l'hyphe et la cellule de la plante et ainsi faciliter l’échange de métabolites entre les deux partenaires.
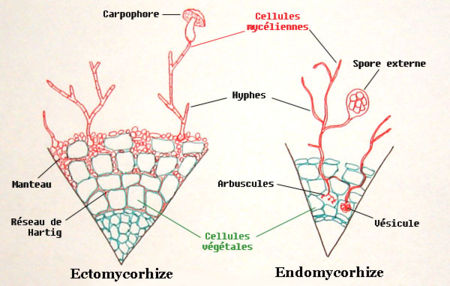 Il y a deux types principaux de mycorhizes : les endomycorhizes et les ectomycorhizes dont voici des vues en coupe très schématiques.
Il y a deux types principaux de mycorhizes : les endomycorhizes et les ectomycorhizes dont voici des vues en coupe très schématiques.Les endomycorhizes sont formés uniquement par des champignons de la division des Glomeromycètes. Ils sont associés avec les plantes herbacées et ligneuses.
Les endomycorhizes colonisent environ 90% des plantes vasculaires terrestres, c'est-à-dire plus de 400 000 espèces. Il existe cependant moins de 200 espèces de champignons endomycorhiziens. Ces champignons ne sont donc pas très spécifiques dans leurs relations de symbiose. Étant peu spécifique, chaque espèce doit posséder un grand potentiel d'adaptabilité et une large diversité génétique afin de lui permettre de s’adapter aux différentes conditions environnementales auxquelles elle doit faire face.
Ils se reproduisent uniquement asexuellement.
Ils sont aussi uniques au point de vue génétique puisque leurs spores possèdent plusieurs noyaux génétiquement différents [1].
Dialogue plante-hôte champignons
Les inducteurs de la plante-hôte
Des chercheurs ont découvert récemment que la plante-hôte secrète des molécules de type strigolactone induisant également la germination de graines de striga (phénomène connu depuis quarante ans chez cette plante parasite).(symbiose fixatrice d'azote).
(les strigolactones n'ont rien à voir avec les facteurs nod et la chitine (cook et al. 1966, besserer et al. 2006, Akiyama et al 2005).)La voie de signalisation Myc
La voie de signalisation Myc est peu connue. Les facteurs Myc (par analogie avec les facteurs Nod) restent à identifier.
La voie Myc présente des étapes communes avec la voie Nod: au niveau des récepteurs de type SYMRK, RLK, de DMI1et DMI2 et des oscillations calciques. Les seules différences sont la fréquence et l’intensité des oscillations calciques, ainsi que la réponse de DMI3, d’où l’intérêt de connaître ses substrats. Les endomycorhizes étant apparues avant les endosymbioses fixatrices d’azote, les chercheurs émettent l’hypothèse que la voie de transduction du signal mycorhizien a été recrutée par les bactéries fixatrices d’azote.
Histoire des mycorhizes
L'endosymbiote originel (un Gloméromycète semble-t-il), serait apparu il y a environ 450 millions d'années au Paléozoïque, probablement au même moment que les premières plantes terrestres. Des fossiles de la flore de Rhynie (Aglaophyton...), d'environs 400 millions d'années, comportent des mycorhizes morphologiquement identiques aux Glomales. Ceci laisse penser que les mycorhizes ont été l’instrument d'une colonisation accélérée des terres émergées, par leur capacité à extraire l’eau et les minéraux du sol.
Ces premières associations ont aussi pu permettre la constitution d'un sol résistant mieux aux intempéries, stockant mieux l'eau, tout en améliorant la résistance des plantes au stress hydrique, leur tenue au vent et au ruissellement, ainsi peut-être que leur résistance aux trop fortes intensités lumineuses, comme c'est le cas des mycophycobiontes de certaines algues de l'estran (ces symbioses, avec des ascomycètes principalement, sembleraient être apparues secondairement).
Cette symbiose correspond à un saut macroévolutif (bond de l'évolution) puisque les fonctions d’un des partenaires s’associent aux fonctions de l’autre, avec des effets fonctionnels multipliés. En effet on passe de l’état algue et champignon à celui de plante mycorhizée terrestre (ce qui va à l'encontre du gradualisme Darwinien).
A l’heure actuelle, 85 % des Archégoniates, ainsi que des Ptéridophytes et des Hépatiques, sont endomycorhizés par des Glomales. Cela suppose que la symbiose avec les Glomales est la plus ancienne chez les Archégoniates et qu’elle aurait permis l’impressionnante radiation de ces derniers (diversité, lignification…).
Les autres familles de Gloméromycètes (Acaulosporaceae et Gigaspora ) sont apparues plus tard vers -250/-230 millions d’années. Elles possèdent des capacités supérieures pour l’exploitation des ressources minérales des sols.
Les ectomycorhizes, quant à elles, seraient peut-être apparues au Crétacé mais les plus vieux fossiles connus ne datent que de l’Eocène. Elles ont permis la colonisation de sols antérieurement défavorables. C’est également à cette époque que des symbioses fixatrices d’azotes sont apparues.
La symbiose ectomycorhiziennes est apparue de nombreuses fois indépendamment dans différents clades des champignons (Gloméromycètes, Ascomycètes, Basidiomycètes) et de Spermaphytes (Gymnospermes, Angiospermes). On constate qu’un certain nombre de plantes contractant une symbiose mycorhizienne, plus récente d’un point de vue évolutif, sont susceptibles de contracter aussi une association à mycorhizes à arbuscules. Il semble donc qu’il y ait eu des innovations évolutives dans ce type de symbiose. Ces innovations expliquent sans doute le nombre limité de plantes capables les contracter.
L'apparition d'ectomycorhizes a été corrélée deux fois à des radiations évolutives des plantes : - Au Crétacé (apparition des pinacées et des rosidées) - Lors de la transition Éocène-Oligocène (apparition des forêts '« actuelles »' de l'hémisphère Nord). Plus récemment encore d’autres formes d’endomycorhizes et d’ectomycorhizes à pelotons sont apparues chez les Ericales. Leurs symbiotes fongiques ont des capacités saprophytes encore plus fortes permettant à la plante de réingérer directement le carbone organique par le biais du champignon, dans les sols où l’activité de minéralisation est très faible (landes, haute montagne, écosystèmes froids…) : la symbiose permet un couplage de niveaux trophiques.
Les angiospermes sans mycorhizes (Brassicacées, Chénopodiacées...) les auraient perdus secondairement.
Importance écologique de la mycorhization
Les mycorhizes sont à l'origine des écosystèmes les plus complexes, et en particulier des forêts tropicales qui poussent souvent sur des sols pauvres. Leurs mycéliums forment des réseaux interconnectés qui influencent le fonctionnement des écosystèmes (cycles biogéochimiques, composition des communautés végétales, alimentation carbonée des plantules pendant leur développement, modification de la compétition...) en permettant ou augmentant des flux importants de carbone organique et de minéraux (azote, phosphore, eau...) via le sol. Ils constituent un des éléments les plus dynamiques de la symbiose mycorhizienne. Ces transferts sont si efficaces, qu'ils remettent en cause le concept de spéciation par compétition pour les nutriments entre les plantes d'un écosystème, en particulier pour la capture des phosphates par les racines (ils permettent de se passer des fertilisants phosphatés) ou pour la résistance à la sécheresse. Ils sont pourtant encore peu exploités en horticulture, agriculture et foresterie, ou pour la dépollution de certains sols pollués.
La colonisation des systèmes racinaires, le potentiel mycorhizogène du sol et la dépendance mycorhizienne des plantes sont inversement corrélés avec la teneur de la solution du sol en ions phosphates; De plus, ce résultat n'est pas lié à une forme d'engrais phosphaté, qu'il soit organique ou minéral, puisque les plantes n'absorbent que des ions en solution. L'enrichissement de cette solution devient directement responsable du fait que la plante bien nourrie ne favorise plus le développement des mycorhizes. Dans certaines situations, les niveaux de phosphore atteints deviennent incompatibles avec l'installation des mycorhizes.[2]
Les mycorhizes interagissent avec diverses bactéries du sol (dont Pseudomonas) appelées bactéries auxiliaires à la mycorhization (en anglais MHB: Mycorrhizal Helper Bacteria). Les mycorhizes interagissent aussi avec les autres mycorhizes et les autres champignons et avec certains prédateurs et parasites aériens des plantes :
- L'attaque des plantes par des herbivores provoque une modification rapide des communautés mycorhiziennes (les espèces demandant le moins de carbone sont favorisées) mais la nature de la population mycorhizienne modifie également (positivement ou négativement) les capacités de défense des plantes.
- Les communautés mycorhiziennes répondent (par modification des abondances spécifiques relatives) aux modifications de l'environnement suivant qu'elles soient plus ou moins favorable à l'hôte ou au symbiote et les communautés bactériennes sont modifiées par la variation des exsudats entre racines mycorhizées et non-mycorhizées.
- La diversité des champignons mycorhiziens à arbuscules du sol contrôle la composition des communautés végétales par un effet direct du champignon sur la valeur sélective des individus qu’ils colonisent (effet bénéfique, neutre, négatif ou même suppressif). Ce phénomène est lié à la préférence d’hôte existant pour chaque champignon. Cette préférence résiderait dans l’adéquation entre fonctions écologiques exercées par le champignon et besoins de la plante hôte.
Prise en compte dans la gestion des forêts et des cultures
Un cortège mycorhizien optimal agirait comme un « engrais » sans ses inconvénients, et gratuitement, en se passant de pesticides, fongicides et amendements. Après de premiers essais en Australie dès les années 1920, des entreprises ont développé des souches d'inoculation adaptées à plusieurs espèces commerciales, mais il faudrait qu'elles soient aussi adaptées au contexte du lieu d'implantation. Plusieurs chercheurs estiment que les souches de champignons symbiotes génétiquement les mieux adaptées au terrain, sont celles qui sont indigènes ([3].
L'activité sylvicole et agricole peuvent perturber ou modifier (négativement ou positivement) la microflore fongique et sa capacité à mycorhizer ;
- Le retournement du sol (labour), le tassement par les engins endommagent la communauté ectomycorhizienne
- La fertilisation azotée (qui se fait de plus en plus en sylviculture aussi) diminue le nombre de carpophores et altère la composition en espèces (mesuré sur Picea abies par Peter et al.[4], en réduisant le nombre de mycorhizes [5], et en changeant les types mycorhiziens (Brandbrud, 1995 ; Karen, 1997)
- Les coupes rases endommagent la communauté mycorhizienne[6]. Jones estime que le changement de composition des communautés ectomycorhiziennes est plus en cause qu'une diminution du taux de colonisation des racines. Le tassement du sol pourrait être en jeu aussi, de même à long terme que l'exportation de presque toute la biomasse ligneuse.
- Les coupes forestières d’éclaircie[7],[8], ainsi que l’irrigation[9] augmentent le nombre de carpophores produits avec des effets encore mal compris sur la composition et biodiversité de la communauté fongique.
- Bien entendu l'utilisation de fongicide affecte cette communauté. La présence de fongicides dans les eaux météoritiques (pluies, brumes, rosées, neige, etc.) est avérée, mais ses impacts sur les champignons restent mal compris.
Les changements bio-chimiques et biologiques, ainsi que microclimatique induit par les grandes coupes rases, (ou même par les moissons dans le cas de l'agriculture) pourraient avoir des impacts sous-estimés, liés aux impacts directs sur le sol, sur l'eau, mais aussi à la perte d’inoculum fongique (même si les racines restent dans le sol, les gros bois mort deviennent rares ou absents, et les rémanents sont souvent rassemblés).
De nombreuses expériences de mycorhization contrôlée ont montré que la régénération naturelle ou artificielle bénéfiçiait de la présence ou inoculation de champignons symbiotes adaptés aux plantes et au contexte. Inversement la trufficulture n'a pu s'exporter en Nouvelle-zélande qu'avec les plantes hôtes mycorhizées. De même les sapins de Noël européens n'y ont poussé qu'avec leurs symbiotes. Certains symbiotes sont d’une efficacité spectaculaire : C'est un accroissement de 60% du volume total de Pseudotsuga menziesii qui a été permis en 10 ans sur les boisements issus de plants ectomycorhizés en pépinière avec une souche Laccaria bicolor dite S238N, par rapport aux arbres non inoculés [10].
Précautions : Le risque existe de concurrence avec des espèces locales, voire de pollution génétique. Plusieurs suivis ont montré que de plans inoculés en pépinières avaient en général perdu ce symbiote au profit d'autres issus du sol local, mais ce pourrait ne pas toujours être le cas[11].
La symbiose
Contrairement à une idée répandue, la relation mycorhizienne n'est pas exclusivement symbiotique car les transferts de substances du champignon à la plante n'augmentent pas toujours la valeur sélective (qui est le critère de définition de la symbiose). Les bénéfices, qui varient hautement en fonction du génotype des partenaires et de l'environnement sont difficiles à évaluer, particulièrement pour les plantes pérennes. Elles pourraient bénéficier de la symbiose uniquement pour de courtes périodes à différents moments de leur vie. Il existe en fait un continuum de relations entre les partenaires, de la symbiose au parasitisme en passant par la saprotrophie (la nature de la relation est descriptible en fonction du rapport coût/bénéfice).
Par exemple, Tricholoma matsutake est symbiotique, parasite ou saprophyte suivant son stade de développement et les conditions du milieu ; les mycorhizes des orchidées sont parasites de ligneuses ; Tuber melanosporum (la truffe) tue les plantes qui entourent son hôte...
En fait, la mycorhization est continuellement instable en raison des conflits d'intérêts entre les partenaires et de la pression de sélection qui pousse chaque partenaire à être le plus parasitique possible. Un parasitisme trop important peut conduire à la rupture de l'association. Au-cours du temps la « symbiose » serait apparue et aurait été rompue plusieurs fois : l'association est réversible. Les champignons saprophytes pourraient ainsi être des symbiotes ayant perdu leur hôte. Ainsi, dans certaines associations mycorhiziennes, l’un des partenaires semble exploiter l'autre (soit par nature, soit en fonction des conditions). On entend ici par « exploiteur » un individu qui obtient un bénéfice (augmentation de la valeur selective) sans réciprocité. On peut citer comme exemple les plantes mycohétérotrophes (plus de 200 espèces connues chez les orchidacées, les gentianacées, les burmanniacées...), des plantes aux capacités photosynthétiques nulles ou réduites qui obtiennent leur carbone de leur partenaire mycorhizien, lui même l'obtenant d’un partenaire chlorophyllien au-travers du réseau mycélien.
L'exploitation d'un des partenaires de la symbiose par l'autre suppose des compensations pour l'individu exploité:
- uniformité génétique: la symbiose permet de maintenir l'uniformité génétique de l'espèce qui est parfois plus intéressante évolutivement que la variabilité (c'est le cas des mycorhizhes à vésicules et arbuscules et des éricoïdes).
- transmission verticale: la valeur sélective de l'exploité est liée à celle de son partenaire. C'est par exemple le cas pour les Glomales qui ont perdu toute reproduction sexuée depuis 400 millions d'années et sont dépendant de l'hôte pour leur survie.
- interaction de voisinage: lorsque la densité d'exploiteur est grande, l'exploité peut se réfugier sur les hôtes les moins exploitants et diminuer la valeur sélective des hôtes plus exigeants.
- commensalisme
- promiscuité: capacité à s'associer à plusieurs partenaires (cas des mycohétérotrophes)
Les mécanismes de compensation pourraient être important dans la stabilisation de la symbiose. De manière générale les symbioses se caractérisent par une diminution de la dérive génétique et du taux de spéciation (Cf.théorie du Roi rouge) mais ce n’est pas toujours le cas dans la mycorhization : plus l'association est exploiteuse plus elle est spécifique car l'exploité développe des résistances qui doivent être contournées par l'exploiteur (Théorie de la reine rouge). Dans le cas présent, la pression sélective sur l'hôte aurait conduit à l'apparition de la lignine et favorisé la croissance des tissus vasculaires.
Enfin, la logique et de nombreux indices laissent soupçonner une origine parasitique des mycorhizes :
- près de la moitié des gènes dont l'expression est modifiée par la mycorhization le sont de la même manière lors d'une attaque de parasites fongiques (à titre d'exemple, l’élicitation des chitinases) ;
- au-cours de l'évolution il y a eu contre-sélection des gènes de défense et augmentation des gènes de symbiose. La stimulation des défenses basales (non spécifiques) de la plante par les mycorhizes serait donc un vestige de l'ancienne relation parasitaire entre les partenaires.
- Bien d'autres exemples de symbiose ou d'interactions durables semblent avoir pour origine le parasitisme.
Recherche
Un programme scientifique (FUNDIV), piloté par Jean Garbaye (INRA Nancy) s'intéresse en France depuis 2006 à la conservation et à l'exploitation de la diversité fonctionnelle des communautés d’ectomycorhizes dans les écosystèmes forestiers, et étudie le risque posé par l'introduction volontaire ou non de souches fongiques allogènes [12]
Notes et références de l'article
- ↑ "SymRK defines a common genetic basis for plant root endosymbioses with AM fungi, rhizobia and Frankia bacteria", Gherbi et al., Proceedings of the National Academy of Sciences USA - 04/03/2008, UMR DIAPC, IRD)
- ↑ Fortin, Plenchette et Piché - "Les mycorhizes, la nouvelle révolution verte", quae, 2008, p.96
- ↑ Trappe J.M. 1977. Selection of fungi for ectomycorrhizal inoculation nurseries. Annual Review of Phytopathology 15: 203-222. ;
Navratil S. 1988. The state of the art in mycorrhizal research in Alberta and Saskatchewan. In Proceedings of the Canadian Workshop on Mycorrhizae in Forestry, May 1-4, 1988. M. Lalonde & Y. Piché (Ed). Centre de recherche en biologie forestière, Faculté de foresterie et géodésie, Université Laval, Ste-Foy, Qc. ;
Perry P.D., Molina R. & Amaranthus M.P. 1987. Mycorrhizae, mycorrhirospheres and reforestation. Canadian Journal of Forest Research 17: 929-940. - ↑ Peter M., Ayer F. & Egli S. 2001. Nitrogen addition in a Norway spruce stand altered macromycete sporocarp production and below-ground ectomycorrhizal species composition. New Phytologist 149: 311-325
- ↑ (Newton et Pigott, 1991)
- ↑ Perry P.D. 1995. Self-organizing systems across scales. Trends in Ecology and Evolution 10: 241-244,
Kranabetter, J.M., & Wylie, T. 1998. Ectomycorrhizal community structure across forest openings on naturally regenerated western hemlock seedlings. Canadian Journal of Botany 76: 189-196.,
Jones M.D., Durall D.M. & Cairney W.G. 2003. Ectomycorrhizal fungal communities in young forest stands regenerating after clearcut logging. Tansley review. New Phytologist 157: 399-422.,
Lazaruk l., Kernaghan G., Macdonald S.E. & Khasa D.P. 2005. The impact of partial forest harvesting on the ectomycorrhizae of Picea glauca in northwestern Alberta. Canadian Journal of Forest Research, 35: 1-13 - ↑ Waters J.R., McKelvey K.S., Zabel C.J. & Oliver W.W. 1994. The effects of thinning and broadcast burning on sporocarp production of hypogeous fungi. Canadian Journal of Forest Research 24: 1516-1522, McKelvey K.S., Zabel C.J. & Oliver W.W. 1994. The effects of thinning and broadcast burning on sporocarp production of hypogeous fungi. Canadian Journal of Forest Research 24: 1516-1522
- ↑ Egli S. & Ayer F. 1997. Est-il possible d’améliorer la production de champignons comestibles en forêt ? L’exemple de la réserve mycologique de La Chenéaz en Suisse. Revue Forestière Française 49: 235-243)
- ↑ Le Tacon F., Delmas J., Gleyze R. & Bouchard D. 1982b. Influence du régime hydrique du sol et de la fertilisation sur la fructification de la truffe noire du Périgord (Tuber melanosporum Vitt.) dansle Sud-Est de la France. Acta Oecologica 3-4 : 291-306.
Wiklund K., Nilsson L.O. & Jacobson S. 1995. Effect of irriguation, fertilization and artificial drought on basidioma production in a Norway spruce stand. Canadian Journal of Botany 73: 200-208. - ↑ Selosse M.A., Bouchard D., Martin F. & Le Tacon F. 2000. Effect of Laccaria bicolor strains inoculated on Douglas-fir (Pseudotsuga menziesii) several years after nursery inoculation. Canadian Journal of Forest Research 30: 360-371.
- ↑ Thèse d'André Gagné, Étude moléculaire du cortège ectomycorhizien de plantations de conifères sur des sites forestiers après coupes à blanc, Université Laval, 2005
- ↑ GARBAYE Jean - Vulnérabilité fonctionnelle des communautés d'ectomycorhizes dans les forêts feuillues de plaine plaine face à l'introduction de souches fongiques allogènes : approche par modélisation expérimentale. (Colloque 2006 Invasions biologiques / Voir Bibliographie)
Compléments
Articles connexes
Bibliographie
- Jean-André Fortin, Christian Plenchette, Yves Piché, Les mycorhizes - La nouvelle révolution verte Éditions Quæ et Multimondes, Québec, 2008, 148 p.
- Victoria GOMEZ ROLDAN, "Rôle des strigolactones dans la symbiose mycorhizienne à arbuscule.", PhD, Université Paul-Sabatier, Toulouse, France, 2009.
Liens et documents externes
 Portail de la botanique
Portail de la botanique Portail de la mycologie
Portail de la mycologie
Catégories : Mycologie | Physiologie végétale




Wikimedia Foundation. 2010.