- Modèle logistique
-
Dynamique des populations
La dynamique des populations s’intéresse au développement numérique de toutes les populations d’êtres vivants, et plus particulièrement de celles des animaux sexués. Les répartitions de poids, la composition par âge des individus, l'environnement, la biologie des groupes, et les processus qui influent sur ces changements font également partie de son champ d'étude. Ces études ont pour but, outre de prévoir les accroissements ou diminutions des populations, de comprendre les influences environnementales sur les effectifs des populations. Des études sur ces sujets sont incontournables par exemple pour la gestion de la pêche, la gestion cynégétique, le management des zones protégées, le contrôle des populations d'animaux dit nuisible...
Sommaire
Démographie
La démographie se réserve l’étude des populations humaines, qui diffèrent des populations animales non pas tant par les valeurs des paramètres biologiques (durée de vie, intervalle entre générations, etc.), que par la conscience qu’en a chaque individu, et qui permet l’interrogation des intéressés. Les individus ont un âge, c’est-à-dire une date de naissance. Ils sont les sujets d’« événements », tels que mariage et procréation, dont le repérage suppose soit une enquête auprès des intéressés (recensement ou sondage), soit des institutions appropriées (état civil). Ce repérage exige que soit connu le nom des individus, qui disparaît ensuite dans le traitement des données. Les relevés des lieux de naissance, de séjour, de décès, supposent également, notamment pour l’étude des migrations, un découpage géographique reconnu. La qualité et le progrès des connaissances démographiques dépendent donc des institutions administratives nécessaires à la collecte des données et à la garantie du secret.
L’étude de la précision des effectifs de populations humaines, obtenus par enquête ou par calcul, ou celle de la datation des événements démographiques, en particulier naissances et décès, met ainsi en jeu la psychologie des individus, leur confiance ou leur méfiance envers les enquêteurs et agents recenseurs, leur mémoire sélective des événements familiaux, leur degré d’adhésion aux institutions classificatoires. Un « in-dividu » ne peut par définition être divisé. Le traitement « en continu », avec des décimales d’effectifs, s’oppose au traitement « discret », en nombres entiers. La confrontation des deux approches, nécessaires l’une et l’autre, conduit à mettre le doigt sur l’absolue singularité de l’individu humain. L’imprécision n’est pas une imperfection de la notion de population, c’en est un attribut.
Fluctuation dans le temps des populations naturelles
Populations stables
L'étude de la dynamique des populations s’articule autour de la notion de « population stable » : une population dans laquelle la probabilité de procréer et de mourir ne dépend que de l’âge tend vers une répartition par âge constant, indépendante des conditions initiales, et dite « population stable » (Lotka). Si les régimes de fécondité et de mortalité s’équilibrent, l’effectif est de plus constant. c’est le cas particulier de la « population stationnaire ».
Une population est dite stable lorsqu'elle ne varie pas dans le temps. Il s'agit d'un équilibre dynamique : le taux de natalité est égale à celuis de mortalité. On peut parler de population stable suivant différentes échelles de temps. En effet, la stabilité d'une population est quelque chose de très relatif. Ainsi, la population de coquelicot dans un champ pourra être qualifiée de stable sur trois ou quatre ans, mais à l'échelle d'une année la population varie énormément.
Populations cycliques
Fluctuations saisonnières
Une population enregistre des fluctuations saisonnières lorsque il existe différentes tailles au fil des saisons. C'est le cas des flux migratoires ou lorsque que l'on observe un pic de mortalité après la reproduction.
Fluctuations annuelles
Fluctuations pluriannuelles
C'est le cas des lemmings qui suivent en plus d'une fluctuation annuelle, suivent un variation tous les 4 ans.
Autres types de fluctuations
Il existe aussi des fluctuations apériodiques ou éruptives comme par exemple les invasions.
Les modèles de croissance de populations
Loi de Malthus
Article détaillé : Thomas Malthus.Forme discrète : Malthus considère que s'il n'y a pas de pression du milieu (si le milieu n'est pas limitant) la population grandira de façon exponentielle. Par exemple, la population, en s'affranchissant de facteur limitant (guerre, famine, …) observe une croissance qui suit une loi géométrique, illimitée et donc exponentielle :
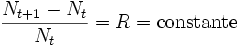 ,
,avec Nt l'effectif de la population étudiée à l'instant t.
Forme continue : La population N(t) est associée à l'équation différentielle
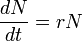
avec r, une constante, représente le taux intrinsèque d'accroissement naturel. Il s'en suit :
N(t) = N0exp(rt). Croissance avec facteur limitant
Modèle de Verhulst
Article détaillé : Modèle de Verhulst.Régulation des populations
La densité-dépendance
Influence des facteurs dépendant de la densité
Compétition intra-spécifique
La compétition intraspécifique correspond à la recherche par les individus d'une même espèce des ressources du milieu identiques.
Compétition inter-spécifique
La définition et le rôle de la compétition interspécifique ont fait l'objet de nombreux travaux contradictoires, voire polémiques.
Interaction positives entre les espèces
voir les articles : commensalisme, mutualisme, coopération, symbiose, prédation, parasitisme
Outils d'acquisition de données
Les populations animales sont plus difficiles à étudier car souvent mobiles et discrètes. De nombreuses espèces sont discrètes ou ne sont actives que la nuit, ou vivent à grande profondeur, ce qui rend leur observation difficile.
Des modèles existent, mais qui demandent souvent à être confirmés par des observations de terrains. Ceci se faisait autrefois par le piégeage et/ou via des indices de présences (chant, terrier, empreinte (avec ou sans "piège à trace"), épreinte, marquage du territoire, poils, pelote de réjection, etc.). Une technique souvent utilisée consiste à évaluer les évolutions spatiotemporelles de population par la méthode dite de capture-recapture, qui est néanmoins stressante pour l'animal, et qui pour cette raison pose problème pour des espèces vulnérables ou menacées.De méthodes récentes permettent un monitoring plus fiable et moins impactant sur les populations ; Elles peuvent combiner les avantages de deux progrès techniques :
- Les appareils automatique de détection (dont écoute ou vision nocturne ou sous-marine, amplification lumineuse, vision infrarouge et autres dispositifs de mesure automatique, compteurs d'entrée sortie d'un nid, d'une tannière, d'un gîte collectif, etc)
- Les analyses d'ADN, qui permettent de mieux évaluer - sans contact direct avec l'animal, via les poils ou des excréments - l'état et la dynamique des populations[1]. Par exemple, le Waterford institute of technology a dans les années 2000 mis au point une méthode utilisant un simple tube de plastique (diamètre : 100 mm, longueur : 250 mm ou 500 mm), positionné presque verticalement et au fond duquel est présenté un appât (aile de poulet). Seule la martre est assez agile pour aller y chercher l'appât. L'appareillage, comprend un dispositif (bande collante) collectant un ou quelques poils de la martre ayant pénétré dans le tuyau, ce qui permet par l'étude de l'ADN de ce poil, en utilisant la logique de la méthode « capture-recapture » d'évaluer le sex-ratio, la population locale de cet animal (On a par exemple constaté que les martres visitent chaque nuit les tubes qu'elle ont repéré, tant qu'il contiennent de la nourriture). On peut enregistrer automatiquement l'heure à laquelle la martre vient manger, les conditions de température ou d'hygrométrie, etc. Un dispositif infrarouge associé peut aussi déclencher un appareil photo ou une caméra (infrarouge ou à amplification lumineuse) et/ou compter les allers-et-venues (jusqu'à 5000 "évènements" mémorisables sur 6 mois). Ces études permettent de mieux évaluer la sensibilité d'une espèce animale discrète à la fragmentation de leur territoire, au roadkill, le type d'habitat qu'elles fréquentent, qu'elles choisissent pour élever leurs petits. Elles donnent des informations précises sur la présence/absence de l'espèce, le sexe-ratio, les liens familiaux des individus se nourrissant au même endroit)[2]. Divers programmes d'études[3][4], et de mieux la connaître y compris dans des milieux à forte naturalité comme la forêt de Bialowieza en Pologne[5]. Cette connaissante améliore les chances de réussite des programmes de réintroduction[6] ou de confortement des populations[7] la concernent ou sont en projet en Europe.
Voir aussi
Sources
- Dajoz R. (1974) Dynamique des populations. Masson et Cie, Paris, 301 p.
- ↑ Statham, M., Turner, P.D. and O’Reilly, C. 2005. Use of PCR amplification and restriction enzyme digestion of mitochondrial D-loop for identification of mustelids in Ireland. Irish Naturalists Journal 28: 1-6.
- ↑ Dispositifs mis au point par le Laboratoire Molecular Ecology (Dr Peter Turner, Dr Catherine 0'Reilly, Mark Statham, Tom Roche & Jacinta Mullins) et testé dans le Kent (Wildwoodland).
- ↑ Balharry, E. A., G. M. McGowan, H. Kruuk, and Halliwel, E. 1996. Distribution of pine martens in Scotland as determined by field survey and questionnaire. SNH Survey and Monitoring Report No. 48. Scottish Natural Heritage, Edinburgh, UK.
- ↑ Hughes, D. 1993. Mammal survey of five species: Red Squirrel, Grey Squirrel, Pine Marten, Irish Hare and English Hare. Ulster Wildlife Trust and Forest Service DANI, Crossgar, Northern Ireland.
- ↑ Zalewski, A., W. Jedrzejewski, and B. Jedrzejewska. 1995. Pine marten home ranges, numbers and predation on vertebrates in a deciduous forest (Bialowieza National Park, Poland). Annales Zoologici Fennici 32: 131-144.
- ↑ Bright, P. W., and Harris, S. 1994. Reintroduction of the pine marten: feasibility study (étude de faisabilité). English Nature / Contract Report.
- ↑ Bright, P.W. and Smithson, T.J. 1997. Species recovery programme for the Pine Marten in England: 1995-96. English Nature Research Reports Volume 240
Catégories : Écologie des populations | Démographie
Wikimedia Foundation. 2010.