- Campylobacter Jejuni
-
Campylobacter jejuni
Campylobacter jejuni
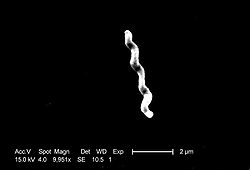
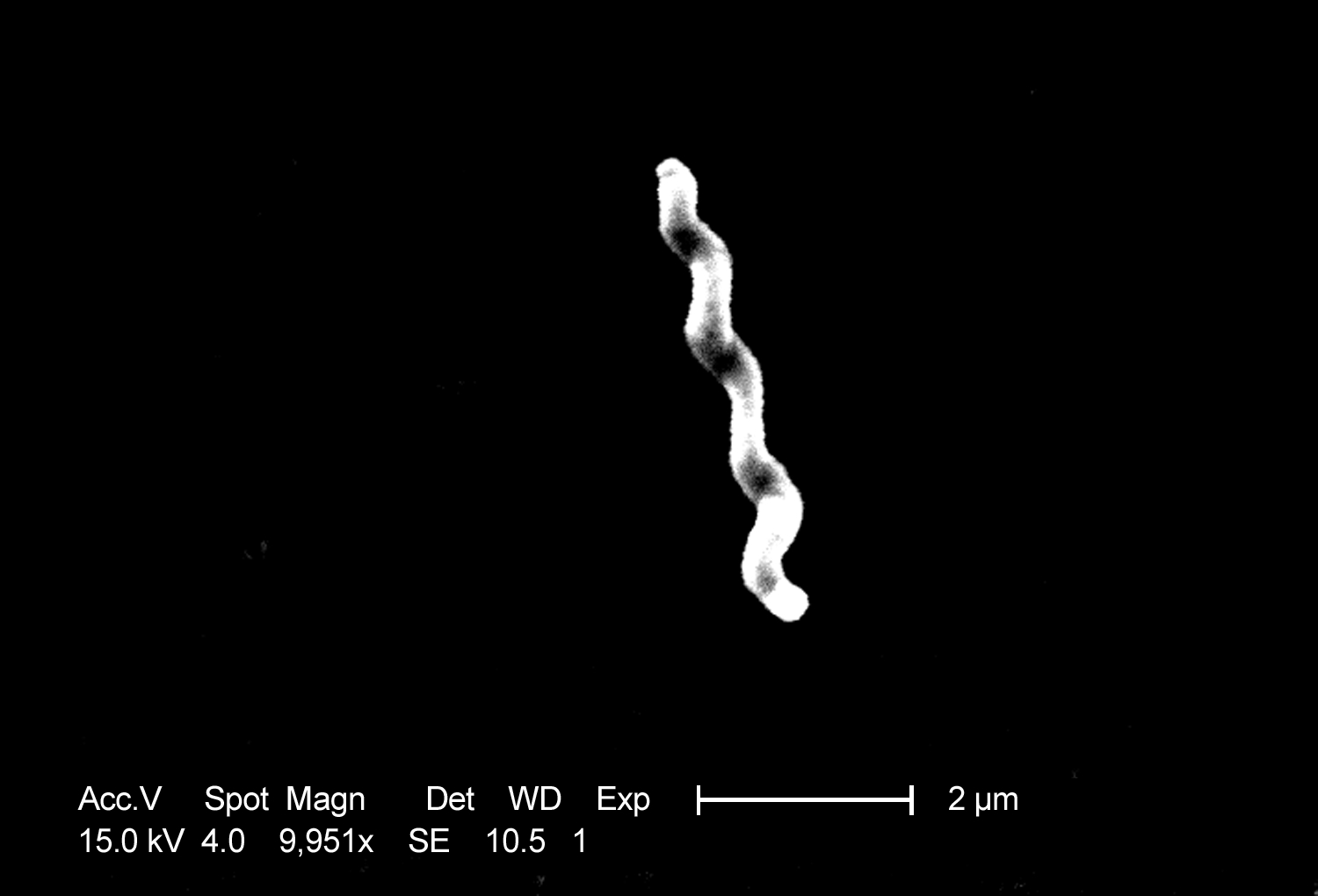
Photographie au microscopie électronique de
Campylobacter jejuniClassification classique Règne Bacteria Embranchement Proteobacteria Classe Epsilon Proteobacteria Ordre Campylobacterales Famille Campylobacteraceae Genre Campylobacter Nom binominal Campylobacter jejuni
(?F.S.Jones M.Orcutt & R.B.Little 1931)
Véron & ?R.Chatelain 1973Parcourez la biologie sur Wikipédia : 





Les bactéries de l'espèce Campylobacter jejuni sont des organismes connus depuis plus d’un siècle comme agents responsables de diarrhées et d'avortements (campylobactérioses) au sein de troupeaux de bovins et de moutons, mais ils ne sont considérés comme ayant une incidence dans la santé humaine que depuis quelques décennies (13). Décrits réellement comme pathogènes alimentaires depuis 1972 (8) ils font l’objet depuis 1982 d’une surveillance par les services de santé publique (13).
Aujourd’hui on sait que C. jejuni est la première cause de pathologie entérique d’origine bactérienne dans les pays industrialisés. Pour exemple, les Campylobacter sont isolés sur 5 % des patients américains et anglais atteints de diarrhées, et l’incidence annuelle de ces infections est évaluée à 50 cas pour 100 000 habitants aux États-Unis, dépassant celles estimées pour les Salmonella et les Shigella (35). On a estimé que plus de 1 % de la population de l’Amérique du Nord est touchée par une campylobactériose chaque année.
Parmi les 16 espèces que contient le genre Campylobacter, 12 ont été retrouvées associées à des entérites bactériennes, avec une nette implication de deux espèces : Campylobacter jejuni ainsi que Campylobacter coli qui sont retrouvées dans environ 95 % des campylobactérioses recensées chez l’homme.
Une infection à Campylobacter jejuni peut engendrer dans de rare cas, des neuropathies auto-immunes sévères (syndromes de Guillain-Barré et de Miller-Fisher) provoquées par un mimétisme moléculaire entre les gangliosides (ou glycosphingolipides) exprimés chez la cellule nerveuse et les lipooligosaccharides présents au niveau de la membrane externe de C. jejuni (59). Le syndrome de Guillain-Barré est considéré comme assez répandu à travers le monde (1 à 2 cas/100 000 habitants/an) avec des incidences plus importantes dans le sud-est asiatique (33).
Les campylobactérioses souvent sporadiques et rarement épidémiques (environ 10% des cas totaux de cette maladie) sont communément acquises par ingestion de viande de volaille insuffisamment cuite pour les cas dispersés, de lait non pasteurisé ou d’eau contaminée suite à des défauts de traitement pour les cas endémiques. C. jejuni colonise le tractus gastro-intestinal des oiseaux préférentiellement, où il s’installe au niveau du cæcum, du gros intestin et du cloaque, et celui des animaux et de l’homme où il envahit l’iléon et le colon. En 2007, un peu plus de 200 000 personnes ont été atteintes en Europe, avec une nette augmentation en Allemagne et un peu moindre en grande-Bretagne[1].
Le réservoir est principalement le tube digestif des volailles et dans une moindre mesure celui des animaux domestiques. La principale voie de contamination vers l’homme s’effectue via les pièces de volailles destinées à la consommation. En effet la grande majorité des carcasses de poulets (entre 50 et 90 %) sont contaminées suite aux processus d’abattages (29).
Campylobacter jejuni présenté comme un micro-organisme exigeant est capable de surmonter plusieurs stress environnementaux durant sa période de transmission à l’homme tel qu’un environnement aérobie. De facto il est toujours difficile à l’heure actuelle d’expliquer l’importante incidence des campylobactérioses aux regards de ses exigences de croissance mais également aux regards des autres entéro-pathogènes gastriques. Cependant, depuis février 2000, la séquence annotée de la souche de C.jejuni NCTC11168 a été publiée (36) et ouvre de nouvelles possibilités pour caractériser ce germe pathogène alimentaire qui dispose d’un mode de régulation qui s’orchestre différemment des modèles d’E. coli et de B. subtilis.
Sommaire
Taxonomie et phylogénie
Campylobacter a d'abord été classé dans la famille des Spirillaceae de par sa morphologie (forme spiralée et hélicoïdale): c'est un genre qui contient 16 espèces recensées à ce jour. Ce genre bactérien a été proposé en 1963 par Sebald et Véron (44) pour une bactérie préalablement connue sous le nom de Vibrio fetus. Des études de phylogénie par hybridation ADN-ADN puis ADN-ARNr (40, 52) et par comparaison des séquences ARN16S, véritable chronomètre de l’évolution ont montré que le genre Campylobacter appartient à la superfamille VI ou branche ε de la division des Proteobacteriae. Le genre Campylobacter constitue, avec les genres Arcobacter et Sulfurospirillum, la famille des Campylobacteraceae.
Morphologie et structure de Campylobacter jejuni
C. jejuni est un bacille à Gram négatif, fin, incurvé et de forme spiralée, de 0,2 à 0,5 µm de diamètre sur 0,5 à 8 µm de longueur (Figure 1). Cette bactérie présente généralement une ondulation qui lui donne un aspect en virgule ou en « S » et quelquefois en hélice pour les plus longues. Elle est asporulée et possède un ou deux flagelles polaires de taille variable qui peuvent atteindre trois fois la longueur du bacille. Ils lui confèrent une grande mobilité dite « en vol de mouette », qui est importante dans le phénomène de colonisation du tractus intestinal. La présence d’une capsule a été démontrée (20) et elle aurait des répercussions dans la virulence et la variabilité antigénique de C. jejuni .
Caractères culturaux
Campylobacter est cultivé sur un milieu sélectif particulier "CAMP" à 42 °C, la température normale du corps des oiseaux plutôt qu'à 37 °C, la température utilisée pour la plupart des autres bactéries pathogènes. Puisque les colonies sont capables d'utiliser l'oxygène de l'air, elles pousseront en surface des milieux de cultures mais elles préfèrent des conditions microaérophiles (elle privilégie les atmosphères pauvres en dioxygène) et capnophile, nécessitant une atmosphère de croissance aux proportions suivantes : 5% O2 (dioxygène), 10% CO2 (dioxyde de carbone) et 85% N2 (diazote). Elle se multiplie entre 30°C et 47°C avec une température optimale de croissance à 42°C. Elle est dite thermotolérante (comme C. lari, C. coli et C. upsaliensis) par comparaison avec d’autres espèces ne se cultivant pas à 42°C mais capables de se développer à 25°C (23). La zone optimale de pH pour obtenir une croissance convenable se situe entre 6,5 et 7,5 (9).
Caractéristiques Résultat Croissance à 25 °C - Croissance à 35-37 °C - Croissance à 42 °C + Réduction des nitrates + Test à la catalase + Test d'oxydase + Croissance sur agar de MacConkey + Mobilité + Consommation de glucose - Hydrolyse de l'hippurate + Résistance à l'acide nalidixique - Resistance à la céphalothine + Génome de Campylobacter jejuni NCTC11168
Actuellement, deux souches de C. jejuni ont été séquencées la souche NCTC11168 (36) et la souche RM 1221 (12). Nous nous intéresserons uniquement ici à la souche NCTC11168.
Le génome de la souche NCTC11168 affiche un total de 1 641 481 paires de bases (pb) et est constitué d’une unique molécule d’ADN circulaire au pourcentage en G+C de l’ordre de 30,6%. 1654 séquences codantes ont été prédites, parmi lesquelles 20 représentent probablement des pseudogènes et 54 des ARN stables. La taille du génome est très en deçà de celle d’E. coli (4, 4-5 Mb) et de L. monocytogenes (2,9Mb) ce qui expliquerait l’exigence nutritive de C. jejuni.
La longueur moyenne d’un gène est estimée à 948 pb et 94,3% du génome code des protéines, contre 87% à 87,9% pour les différentes souches de l’espèce E. coli, faisant de lui l'un des génomes bactériens les plus denses. Une étude des régions inter-géniques a permis d’établir que celles-ci représentent 4,3% du génome et n'était pas distribuée de manière homogène (36).
L’organisation des gènes est atypique. Effectivement, mis à part les deux opérons codant les protéines ribosomales et les groupes de gènes impliqués dans la biosynthèse des lipooligosaccharides, des polysaccharides extracellulaires et dans la modification du flagelle, peu de gènes paraissent agencés en opérons ou clusters, y compris les gènes de biosynthèse des acides aminés. On dénombre qu’une seule séquence d’insertion similaire en partie à l’IS605 tnpB d’H. pylori mais aucune autre séquence d’insertion, transposon, rétron ou prophage.
Le séquençage a mis en exergue la présence de 25 régions polymorphiques hypervariables riches en G. Ces régions se situent préférentiellement parmi les gènes intercédant dans la biosynthèse ou le changement de structures de surfaces telles que le flagelle, la capsule et les lipooligosaccharides. L’existence de ces séquences singulières provoquerait un taux de mutation plus conséquent dans ces secteurs du génome (36). Ces possibles mutations et réarrangements par glissement permettraient à la bactérie d’éviter une réponse du système immunitaire de l’hôte par la modifications des structures de surfaces immunogènes.
L’expression des gènes chez C. jejuni n’obéit pas aux mêmes promoteurs que chez E. coli (49). L’étude du génome de la souche NCTC11168 a permis de mettre en évidence l’existence de trois facteurs sigma permettant une expression spécifique de certains gènes. Il s’agit de RpoD ou σ70 (facteur σ qui garantit la transcription des gènes assurant les fonctions indispensables à la vie de la bactérie) et des facteurs FliA ou σ28 et RpoN ou σ54 impliqués dans le contrôle des gènes de l’appareil flagellaire.
À l’instar d’H. pylori, le génome de C. jejuni possède peu de facteurs sigma, soulevant ainsi l’hypothèse de l’existence d’une mise en œuvre de l’expression des gènes dissemblable de celle d’E. coli et de B. subtilis.
La survie de Campylobacter jejuni
Dans ses environnements naturels, c'est à dire dans le tractus gastro-intestinal des hôtes aviaires et mammifères, ou bien dans le milieu extérieur durant la transmission à l’humain, C. jejuni doit affronter des conditions de limitation de croissance potentiellement létales telles que : la pression osmotique, les chocs thermiques chaud et froid, les stress oxydatifs, azotés et pH ; ainsi que la limitation en fer ou en nutriments comme le carbone, le phosphate ou le soufre.
À l’heure actuelle, les connaissances acquises sur l’adaptation de C. jejuni à différentes conditions environnementales sont basées sur les analyses des séquences des génomes des souches NCTC11168 et RM 1221 et des expressions différentielles observées en réponse à des changements de température ou changements de milieu de culture (45, 34, 51). Ainsi, C. jejuni possédant trois facteurs σ, montre des systèmes de régulation différents de ceux d'E. coli et de B. subtilis avec environ 37 régulateurs répertoriés comme fonctionnels ou potentiels, et plusieurs systèmes à deux composantes (Tableau 1). C. jejuni ne possède ni RpoS (36) ou facteur σ de la réponse de la phase stationnaire ou de la carence, ni RpoH ou facteur σ de la réponse à un stress thermique chaud chez E. coli ou σB ou CtsR chez B. subtilus, ; pourtant la bactérie synthétise un groupe de protéines hautement résistantes appelées "protéines de choc thermiques" (HSP) suite à une augmentation de température.
Vingt quatre protéines HSP ont été identifiées chez C.jejuni. Parmi ces protéines, il fut identifié des homologues des gènes dnaJ, dnaK, groEL, groES, grpE, hrcA, hspR, clpB et lon. HrcA pourrait servir de régulateur négatif des gènes grpE et dnaK comme chez B. subtilis car il a été découvert dans la région promotrice de l’opéronhrcA-grpE-dnaK de C. jejuni un élément CIRCE (pour Controlling Inverted Repeat of Chaperone Expression) qui correspond à la séquence d’ADN que fixe la protéine HrcA. Cette séquence est également présente dans l’opéron groESL. C. jejuni contient aussi un homologue d’un autre régulateur, HspR retrouvé chez Streptomyces albus. Dernièrement des études ont révélé qu’HspR régulait l’expression de dnaK, groELS et cbpA chez C. jejuni (23, 45).
Concernant la réponse au choc froid C. jejuni ne possède pas d’homologue de la protéine majeure de réponse au choc froid d’E. coli qui est l’ARN chaperonne CspA (Cold shock protein A). Néanmoins, il a été montré qu’à 4°C, C. jejuni était mobile, consommait du dioxygène, synthétisait des protéines et était capable de survivre (17, 45). Cependant si aucun régulateur de la réponse du choc au froid est à ce jour connu, il semblerait que la bactérie ressente le choc au froid comme un choc oxydatif car plusieurs gènes dont les protéines sont impliquées dans la réponse au stress oxydatif voient leurs transcriptions augmentées à basses températures (45).
De plus, C. jejuni semble également s’adapter au froid en acquérant ou biosynthétisant des cryoprotecteurs et/ou des protéines stabilisatrices, et en maintenant une traduction efficace (45).
Campylobacter jejuni de par son cycle de vie évoqué précédemment, doit pouvoir survivre dans un milieu hostile tel que le milieu extérieur qui peut, au vu des ressources nutritives accessibles, être assimilé à la phase stationnaire du cycle physiologique classique d’une population bactérienne. On observe que des cultures âgées ou ayant été exposées à des conditions de stress, présentent des modifications dans la morphologie des cellules. Deux formes prédominent dans la phase exponentielle de croissance : "bacillaire et vibrioïde" tandis que la forme "coccoïde" est retrouvée dans une culture âgée ou stressée (21).
Une telle réduction de la taille de la cellule se déroule également chez d’autres bactéries que sont Vibrio vulnificus et Helicobacter pylori où cette transformation est associée à un état viable mais non cultivable (21). Dans la majorité des espèces bactériennes caractérisées à ce jour, l’entrée en phase stationnaire ou en carence, est accompagnée par de profonds changements structuraux et physiologiques qui se traduisent par une augmentation de la résistance aux chocs chaud, osmotique, aux stress oxydatif, acide.
Chez E. coli lors de l’entrée en phase stationnaire ou en carence plus d’une trentaine de gènes sont induits dont le facteur de la réponse générale, le facteur RpoS ou σS. L’analyse in silico de la séquence génomique de la souche NCTC11168 indique que le gène codant la protéine RpoS est absent. Cependant C. jejuni est capable de produire des facteurs de résistance lorsqu’il est mis en condition de carence. En effet, bien que la réponse de C. jejuni en condition de phase stationnaire et/ou carence soit peu connue, il a été observé dans les premières heures de ces conditions une synthèse protéique. Mais à l’heure actuelle cette synthèse protéique n’a pas encore été élucidée, cependant deux hypothèses s’affrontent soit cette production de protéines est le résultat d’une réponse spécifique induite par le stress, soit il s’agit d’une synthèse continue de protéines non adaptatives. Il faut noter également que C. jejuni est capable de mettre en place une réponse d’austérité face à une carence en acides aminés et/ou carbone. Car il est doté dans son génome d’homologues des enzymes du métabolisme de la guanosine polyphosphate («(p)ppGpp») que sont Ndk (Nucléoside diphosphate kinases), Gpp (Guanosine 5'-triphosphate,3'-diphosphate (pppGpp) pyrophosphatase) et SpoT (ppGpp synthetase) (14).
Il est également connu que lors de l’adaptation à des changements environnementaux, les régulations post-transcriptionnelles à savoir : contrôle de la stabilité des ARNm, transcrits antisens, inhibition de la traduction, sont des caractéristiques décisives pour l’expression des gènes. Des études d’intégration de C. jejuni en microcosme aqueux froid (18) sur deux souches : Bf et NCTC11168, ont révélé après analyse des protéomes, l’existence d’une synthèse de protéines en réponse aux stress et une diminution de certaines voies métaboliques.
Les données recueillies en transcriptomique sur la souche NCTC11168 ont permis de déceler la surexpression d’une protéine de ce stress aqueux froid (45). Il s’agit de la protéine CstA (gène Cj0917c chez NCTC11168). Chez E. coli CstA (carbon storage starvation) est un transporteur de peptide (10) et est impliqué dans le métabolisme des peptides (43). Son expression est induite par la carence en carbone et son induction nécessite l’AMPc et CRP (cAMP Receptor Protein) (43). Sa traduction est sujette à une régulation par CsrA (Carbon storage regulator) (10). Considéré comme un régulateur chez E. coli.
Notes et références
- ↑ The Community Summary Report on Trends and Sources of Zoonoses and Zoonotic Agents in the European Union in 2007, European Food Safety Authority
Liens externes
- Référence Catalogue of Life : Campylobacter jejuni (en)
- Référence NCBI : Campylobacter jejuni (en)
Catégorie : Bactérie
Wikimedia Foundation. 2010.