- Virus de la rougeole
-
 Virus de la rougeole
Virus de la rougeole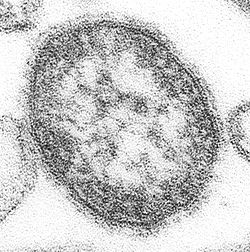
Measles virus Classification des virus Type Virus Groupe Groupe V Ordre Mononegavirales Famille Paramyxoviridae Sous-famille Paramyxovirinae Sous-famille Morbillivirus Espèce Virus de la rougeole 
D'autres documents multimédia
sont disponibles sur CommonsLe virus de la rougeole (aussi MV pour measles virus) est un virus appartenant au genre morbillivirus de la famille des Paramyxovirus. C’est un virus très contagieux pouvant causer des problèmes graves, voir mortels, comme des encéphalites. Tout n’est pas connu à son sujet, principalement les récepteurs cellulaires qu’il cible. Il est important de poursuivre les recherches afin de trouver de meilleurs vaccins contre ce virus.
Sommaire
Description
C’est un virus à ARN. Ce virus est rapidement inactivé par la chaleur (30 minutes à 56 °C) ou la lumière (ultra-violets). De plus il est sensible à de nombreux désinfectants (l’éthanol à 70 % par exemple).
Le virion mesure de 100 à 300 nm et est entouré d'une enveloppe lipidique.
Gènes
Le virus comporte six protéines[1] dénommées : F, H, L, M, N et P :
- M matrice
- N nucléocapside
- P phospholipide
- L large
- H haemagglutinine
- F fusion
Liste des sous-taxons
- Measles virus Calgary/CAN/40.01
- Measles virus genotypes and isolates
- Measles virus strain AIK-C
- Measles virus strain Biken
- Measles virus strain Edmonston
- Measles virus strain Halle
- Measles virus strain HU2
- Measles virus strain IP-3-CA
- Measles virus strain Leningrad-16
- Measles virus strain Moraten
- Measles virus strain Philadelphia-26
- Measles virus strain Rubeovax
- Measles virus strain Schwarz
- Measles virus strain Shanghai-191 vaccine
- Measles virus strain Yamagata-1
- Measles virus type N
Composantes du virus
Le MV est un virus possédant une enveloppe faite d’une double couche de lipides qui entoure une nucléocapside à symétrie hélicoïdale. Son diamètre varie entre 120 et 300 nm.
Génome
Son génome est un brin d’ARN négatif non segmenté composé de 15 894 nucléotides formant 6 gènes qui codent pour 8 protéines différentes. Un seul promoteur est présent dans le génome et se trouve à l’extrémité 3’. Il a une longueur de 56 nucléotides. À l’autre extrémité se trouve le terminateur d’une longueur de 40 nucléotides. Chaque gène est séparé du suivant par 3 nucléotides qui ne sont pas transcrits lors de la transcription. L’ordre des 6 gènes est, sur le brin positif, N-P/V/C-M-F-H-L, chaque lettre représentant une protéine. Dans le deuxième gène, il y a changement de cadre de lecture lors de la traduction, ce qui permet de former les 3 protéines P, V et C.
Protéines de la nucléocapside
Nucléoprotéine N : Ce sont les protéines internes les plus nombreuses dans le virus. Elles se lient au génome d’ARN, l’entourent et le protègent. Elles s’associent également avec les protéines L et P, formant ainsi le complexe de transcription RNP (ribonucléoprotéine). Le domaine N-terminal de la protéine N sert à lier l’ARN et son domaine C-terminal sert à lier la protéine P. Elle lie également la protéine L, formant un complexe de transcription RNP contenant P, L, N et l’ARN.
Protéine large L : Cette protéine forme avec la protéine P l’ARN polymérase ARN-dépendant faisant partie du complexe de transcription. C’est elle qui assure les fonctions d’une polymérase comme la transcription, la réplication, l’ajout de la coiffe et de la queue poly A à l’ARNm.
Phosphoprotéine P : Cette protéine lie la protéine L à la nucléocapside. Dans sa partie C-terminale se trouve une région nommée PMD (P multimerization domain) pouvant lier la protéine L et à l’extrémité complètement se trouve le site de liaison avec N. Elle reconnait le promoteur et guide L vers lui.
Protéines d'enveloppe
Protéine de fusion F : C’est la protéine qui permet de faire fusionner l’enveloppe du virus avec la membrane de la cellule. Son peptide de fusion se trouve dans sa partie N-terminale. C’est cette protéine qui est également responsable de la formation des cellules de Warthin-Finkeldey, cellules géantes retrouvées lors d’un cas de rougeole. En effet, les protéines de fusion sont exprimées à la surface des cellules infectées, permettant à celles-ci de s’associer à leurs cellules voisines pour former des syncitia. La protéine F se retrouve sous forme trimérique à la surface du virus. Elle est d’abord synthétisée sous une forme inactive appelée F0. C’est la furine protéase d’une cellule hôte qui peut l’activer, donnant 2 sous-unités, F1 et F2, liées entre elles par un pont disulfure.
Hémagglutinine H : L’hémagglutinine est une glycoprotéine qui permet d’attacher le virus à la cellule cible en reconnaissant un récepteur particulier. Elle est retrouvée sous forme dimérique pouvant former des tétramères à la surface du virus. Selon certaines études, sa liaison avec un récepteur cellulaire entrainerait une modification de la forme d’une protéine de fusion adjacente qui libèrerait alors son peptide de fusion hydrophobe et pourrait l’insérer dans la membrane cellulaire.
Protéine de matrice M : Cette protéine hydrophobe recouvre la surface interne de l’enveloppe du virus. Elle n’est pas une partie intégrante de l’enveloppe, mais elle y est reliée. Elle est aussi reliée au complexe RNP, permettant ainsi le maintien de la structure du virus.
Protéines non structurales
Protéine V : La principale fonction de cette protéine est de bloquer la réponse immunitaire innée de l’hôte et ainsi permettre la propagation du virus. Elle se lie à l’hélicase MDA-5, une protéine activant la production d’interféron β, et l’inhibe. Elle interagit également avec Jak1 et STAT1, 2 protéines présentes dans la signalisation induites par le récepteur d’interférons de type 1. Elle bloque donc les voies de signalisation cellulaire en lien avec les interférons.
Protéine C : Tout comme V, cette protéine inhibe la voie des interférons une fois le virus entré dans une cellule hôte. En plus de cela, elle joue un rôle dans l’infectiosité du virus.
Tropisme
Le MV pénètre dans l’organisme par le système respiratoire. Il se réplique d’abord dans les cellules épithéliales de ce système, puis il se répand en infectants les cellules dendritiques résidant dans les muqueuses respiratoires. Ces cellules infectées se rendent ensuite vers les ganglions lymphatiques, où elles transmettent le virus aux lymphocytes et les monocytes présents. L’infection se répand ensuite dans les organes lymphoïdes secondaires. À partir de là, le virus peut se répandre partout dans l’organisme.
Contrairement aux autres paramyxovirus, les morbillivirus n’utilisent pas l’acide sialique comme récepteur. C’est pourquoi ils n’ont pas besoin de neuraminidase à leur surface, une protéine qui sert à briser les acides sialiques. Les deux récepteurs ciblés par le virus de la rougeole connus à ce jour sont le CD46 (ou MCP) et le CD150 (ou SLAM).- CD46 : Ce récepteur se retrouve sur toutes les cellules nucléées. Cette molécule joue un rôle dans la régulation du complément. En fait, elle inhibe l’action du complément en inactivant les composantes C3b et C4b. Elle empêche donc la lyse de la cellule par le système du complément. La liaison du récepteur CD46 avec l’hémagglutinine trouvée à la surface du virus ou à la surface d’une cellule infectée entraine l’internalisation du récepteur. S’en suit alors une fusion des membranes soit du virus et de la cellule cible, soit d’une cellule infectée et d’une cellule avoisinante. Il arrive aussi que les particules virales entrent dans la cellule par endocytose médiée par CD46. Une fois la protéine CD46 à l’intérieur de la cellule, elle perd sa fonction, rendant la cellule vulnérable à la lyse par le complément. Cependant, selon des études, ce récepteur serait très peu utilisé in vivo par le virus.
- CD150 : Cette protéine se retrouve à la surface des cellules NK et des lymphocytes ainsi que sur les cellules dendritiques et les monocytes activés. Une SAP (SLAM-associated protein) peut se lier à la portion cytoplasmique de ce récepteur lorsque celui-ci est activé. Cela déclenche une cascade de signalisation menant à la production d’IL-4 et d’IL-13 par les lymphocytes Th et la production d’IL-12 par les macrophages. C’est le principal récepteur cellulaire du virus de la rougeole.
Cycle de réplication
La réplication du virus de la rougeole se fait comme celle de tous les virus à génome ARN négatif. Cependant, des études ont montré que le noyau aurait peut-être un rôle à jouer, mais des recherches restent encore à faire à ce sujet. Les méthodes utilisées par le virus pour détourner les composants cellulaires à leur profit et leur permettre de s’assembler et de sortir de la cellule ne sont pas encore connus.
L’entrée du virus de la rougeole peut se faire soit par fusion (la plupart du temps), soit par endocytose. Si c’est par endocytose, une étape de plus s’ajoute au cycle. Une protéine présente dans la membrane de l’endosome doit d’abord faire entrer des ions H+ dans l’endosome pour que le milieu devienne acide. Cela induit un changement dans la forme des protéines transmembranaires virales, permettant la fusion de l’enveloppe avec la membrane cellulaire. La nucléocapside est alors libérée dans le cytoplasme.
Après l’entrée du virus dans la cellule, le complexe RNP commence la transcription primaire, c'est-à-dire la formation des ARNm (+) de chacune des protéines à partir du génome ARN ‒. Il utilise les ribonucléosides présents dans le cytosol de la cellule hôte. La polymérase se fixe à l’extrémité 3’ du génome négatif et débute la transcription du premier gène, soit N. À la fin, elle ajoute une queue poly A au gène et commence la transcription du second gène, soit P/V/C. Or, le passage au gène suivant ne se fait pas à chaque fois. La polymérase peut arrêter sa transcription à n’importe quel gène. C’est pourquoi la protéine N est la plus nombreuse et la protéine L est la moins nombreuse dans le virus. Les protéines sont par la suite synthétisées par les ribosomes cellulaires. Les glycoprotéines de l’enveloppe sont synthétisées à partir des ribosomes du réticulum endoplasmique et sont transportées jusqu’à la membrane cellulaire en passant par l’appareil de Golgi. Les autres protéines sont produites dans le cytosol. Le génome peut ensuite être répliqué en antigénome positif. C’est cet antigénome qui sert de matrice pour la synthèse de plusieurs copies du génome du virus.
Les nouveaux génomes s’assemblent avec les protéines N, L et P pour former les nucléocapsides. Celles-ci vont ensuite vers la membrane cellulaire pour se lier sur le domaine cytosolique des protéines d’enveloppe transmembranaires du virus. La membrane se replie et la particule virale est libérée par bourgeonnement.
Bilbiographie
- R. Cattaneo et M. McChesney, Measles Virus, dans B. W. J. Mahy (Éd.), Encyclopedia of virology (3rd éd., Vol. 3). Amsterdam, Boston, Academic Press, 2008, p. 285-288
- Erling Norrby , Measles, dans B. N. Fields, Virology, New York, Raven Press, 1985, p. 1305-1309
- Thierry Borrel, Les virus, Paris, Nathan, coll. Sciences 128, 1996
- Hélène Valentin, Branka Horvat, Pierre Rivailler et Chantai Rabourdin-Combe, Rôle de la molécule CD46 dans l'infection par le virus de la rougeole, Annales de l'Institut Pasteur/Actualités, Volume 8, Issue 2, Juillet-Septembre 1997, Pages 181-184
- D. Gerlier et H. Valentin, Measles virus interaction with host cells and impact on innate immunity, Curr Top Microbiol Immunol. 2009;329:163-91
- Yusuke Yanagi, Makoto Takeda and Shinji Ohno, Measles virus: cellular receptors, tropism and pathogenesis, J Gen Virol 87, 2006
- Peng Zhang, Lingyun Li, Chunlin Hu, Qin Xu, Xin Liu and Yipeng Qi, Interactions among Measles Virus Hemagglutinin, Fusion Protein and Cell Receptor Signaling Lymphocyte Activation Molecule (SLAM)Indicating a New Fusion-trimer Model, Journal of Biochemistry and Molecular Biology, Vol. 38, No. 4, Juillet 2005, pp. 373-380
- C. Speziani, D. Laine, C. Servet-Delprat, H. Valentin et C. Rabourdin-Combe, Virus de la rougeole et immunosuppression, Médecine et maladies infectieuses 34, 2004, S2-S6
- Cedric Bernard , Stéphane Gely, Jean-Marie Bourhis , Xavier Morelli, Sonia Longhi et Hervé Darbon, Interaction between the C-terminal domains of N and P proteins of measles virus investigated by NMR, FEBS Letters 583, 2009, 1084–1089
- «Measles virus, complete genome» (Page consultée le 18 avril 2011) [En ligne], adresse URL: http://www.ncbi.nlm.nih.gov/sites/entrez?Db=genome&Cmd=ShowDShowDetai&TermToSearch=10295
- Laboratoire CERBA, Guide des analyses spécialisées, Elsevier Masson SAS, 5e édition, 2007, p 832-835
- Moselio Schaechter, Gerald Medoff etBarry I. Eisensteinp, Microbiologie et pathologie infectieuse, Paris, De Boeck University, 2e édition, 1999, 427-433
- François Denis, Les virus transmissibles de la mère à l’enfant, Paris, John Libbey Eurotext, 1999, p. 337-343
- Harvey Lodish, Arnold Berk, Paul Matsudaira, Chris A. Kaiser et James Darnell, Biologie moléculaire de la cellule, Paris, De Boeck Université, 3e édition, 2005, p. 140-141
Notes et références
- *Dowling & al., 1986 : Transcriptional Map of the Measles Virus Genome. J. gen. Virol., vol. 67, p. 1987-1992
- NCBI, consulté le 25 mai 2010
Voir aussi
Liens externes
- Référence NCBI : Measles virus (en)
Wikimedia Foundation. 2010.