- Bilharziose
-
 données inconnues <50 50-75 75-100 100-150 150-200 200-250 250-300 300-350 350-400 400-450 450-500 >500
données inconnues <50 50-75 75-100 100-150 150-200 200-250 250-300 300-350 350-400 400-450 450-500 >500Seconde endémie parasitaire mondiale après le paludisme, la bilharziose ou schistosomiase est une maladie chronique et débilitante dont la prévalence atteint les 180 millions d’individus[1]. Un des parasites responsable, Schistosoma haematobium, a été identifié en 1851 par le parasitologiste allemand Théodore Bilharz, d'où le nom de la maladie.
Cette parasitose, retrouvée en zones tropicales et subtropicales en Afrique, en Amérique du Sud et en Asie, est responsable d’environ 280 000 décès chaque année[1]. La morbidité observée chez les populations humaines infectées est essentiellement liée à l’étonnante fécondité du parasite femelle dont les œufs, pondus par centaines chaque jour, sont piégés dans de nombreuses muqueuses et tissus, ce qui est à l’origine de la pathologie.
Sommaire
Taxonomie
Les bilharzioses humaines décrites en Égypte par T.Bilharz, sont dues à un ver hématophage, le schistosome, appartenant à l’embranchement des Plathelminthes (vers plats non segmentés), à la classe des Trématodes (appareil digestif avec cæcum), à l’ordre des Strigeatida (ventouses ventrale et buccale), ) la famille des Schistosomatidés (cercaires libres) et enfin au genre Schistosoma, car l’hôte définitif est un mammifère.
Le genre Schistosoma comporte 18 espèces dont 5 sont pathogènes pour l’homme :
- S.mansoni : bilharziose intestinale aux Antilles et en Amérique centrale.
- S.haematobium : bilharziose urogénitale en Afrique, Inde et Péninsule arabique
- S.intercalatum : bilharziose rectale et génitale en Afrique centrale
- S.japonicum : bilharziose intestinale avec complications artério-veineuses en Chine, Japon et Thaïlande.
- S.mekongi : bilharziose intestinale avec complications artério-veineuses en Chine, Japon et Thaïlande.
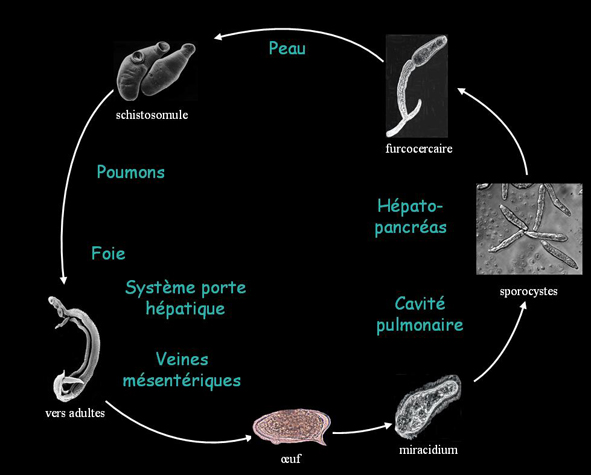
Cycle de vie du schistosome
Le schistosome se développe au cours d’un cycle dixène gonochorique où l’hôte intermédiaire est un mollusque d’eau douce, spécifique de chaque espèce de schistosome (Biomphalaria glabrata dans le cas de S.mansoni), et l’hôte définitif un vertébré (homme, souris, hamster…).
 Schéma du cycle de vie de Schistosoma
Schéma du cycle de vie de SchistosomaLa pénétration de la forme infestante, la furcocercaire (cercaire à queue fourchue) issue du mollusque, chez l’hôte définitif, se fait par voie transcutanée, lors de périodes de baignades en eau douce et stagnante. Au cours de la pénétration, la furcocercaire subit des modifications morphologiques et biochimiques - notamment la perte de la queue - aboutissant à sa métamorphose en schistosomule. Deux à quatre jours après l’infection, le schistosomule quitte le derme, gagne la lumière d’un vaisseau lymphatique ou sanguin, puis est entraîné jusqu’au cœur et aux poumons où il séjourne quelques jours, bloqué au niveau des capillaires sanguins. Le schistosomule subit une maturation, essentiellement morphologique, qui lui permet de quitter les poumons et de gagner le système circulatoire péri-hépatique où il se différencie en schistosome adulte mâle ou femelle sexuellement mûr. La femelle atteint sa maturité sexuelle dès lors qu’elle se loge dans le canal gynécophore du mâle. Le couple migre alors vers le lieu de ponte (plexus mésentériques pour S.mansoni) où il se localise définitivement. A ce stade, la femelle commence à pondre des œufs en continu (jusqu’à 300 par jour pour S.mansoni), dont une partie est piégée dans les tissus où ils formeront des granulomes, à l’origine de la pathologie. L’autre partie des œufs traverse la paroi intestinale ou vésicale pour finalement être excrétés dans les selles ou les urines. L’œuf en eau douce va éclore et libérer une forme larvaire, le miracidium, extrêmement mobile, qui est la forme infestante de l’hôte intermédiaire. Chez les mollusques, le miracidium se transforme en sporocystes primaires puis, par bourgeonnement, en sporocystes secondaires, chacun donnant naissance à une furcocercaire qui sera libérée à des instants précis de la journée, coïncidant généralement avec les périodes de contact aquatique des populations.
Pathologie
Les manifestations pathologiques observées au cours de l’infection par S.mansoni sont essentiellement liées à la formation de granulomes autour des œufs piégés dans les tissus. Le granulome est la conséquence d’une réaction inflammatoire de type hypersensibilité retardée, provoquée par le dépôt des œufs dans le tissu hépatique (1), se traduisant par un afflux de cellules inflammatoires telles que les éosinophiles, les macrophages et les lymphocytes. Ces cellules sont progressivement remplacées par des fibroblastes producteurs de protéines matricielles (collagène) dont l’accumulation va constituer un nodule fibreux. L’évolution en fibrose cicatricielle génère une hypertension portale responsable de l’hépatomégalie. Une circulation collatérale peut se développer, avec formation de varices œsophagiennes, dont la rupture peut provoquer des hémorragies mortelles.
Réponse immunitaire au cours de l’infection
Des études ont révélé l’existence d’une dichotomie de la réponse immunitaire, essentiellement basée sur la production de cytokines par les lymphocytes T (CD4+ et CD8+). Des travaux initiaux, réalisés chez la souris ont mis en évidence une réponse de type 1 (Th1/Tc1), caractérisée par la production d’IL-2 et d’IFN-γ, et de type 2 (Th2/Tc2), caractérisée par la production d’IL-4, -5, -10 et -13 (2). La réponse immune apparaît alors comme une balance dont l’équilibre peut être rompu par la sécrétion de cytokines immunorégulatrices (IL-12 et IFN-γ pour un profil de type 1, IL-4 et IL-10 pour un profil de type 2).
Dans le modèle de la schistosomiase murine, il a été montré qu’une réponse de type Th1 se développe, dirigée contre les formes larvaire et adulte du parasite, et caractérisée par la production d’IFN-γ et d’IL-2 par les lymphocytes T CD4+. La réponse humorale est également polarisée vers un profil Th1 puisque sont détectées majoritairement des immunoglobulines d’isotypes IgG2a et IgG2b. Cependant, cette réponse immune reste faible. Ce phénomène pourrait être lié au fait que les stades cutanés interfèrent avec les fonctions immunostimulantes des DCs. A titre d’exemple, une étude a montré que le schistosomule était capable d’inhiber la migration des cellules de Langerhans vers les ganglions lymphatiques par la production d’une prostaglandine, la PGD2 (3, 4). Par la suite, le dépôt des œufs dans les tissus par les vers femelles induit une réorientation de la réponse immune, spécifique et non-spécifique vers une forte réponse Th2. Cette réponse est caractérisée par la production d’IL-4, IL-5, IL-10 et IL-13. Des immunoglobulines d’isotypes IgG1 et IgE, marqueurs d’un profil Th2, sont également détectées. Les mécanismes précoces responsables de l’induction de la réponse Th2 par les oeufs du schistosome sont cependant encore mal connus. Il est possible que des cellules dendritiques régulatrices (CD11c low) puissent avoir un rôle important.
Traitement
L’antimoine a été utilisé dans le passé pour traiter cette maladie. A faible dose, il se fixe aux atomes de soufre des enzymes du parasite et le tue sans affecter l’hôte. Depuis quelques années, les sujets atteints de la bilharziose sont traités par une prise annuelle d’une dose de praziquantel[2],[3]. Comme c’est le cas pour la majorité des maladies parasitaires, la communauté scientifique s’attèle au développement d’un vaccin qui bloquerait le parasite dans son cycle de développement chez l’homme.
Après plus de 20 ans de recherches internationales, le Bilhvax, un vaccin contre la bilharziose, est entré dans la phase III de son développement. Ce vaccin a été mis au point par les chercheurs de l’Inserm (Institut national français de la santé et de la recherche médicale) et de l’Institut Pasteur de Lille et est produit par Eurogentec, la société de biotechnologie située dans le Liège Science Park au Sart Tilman[4],[5].
Notes et références
- (en)Partnership for Parasite Control, Second Meeting Rome, avril 2002
- Schistosomiasis Control Program
- 20 April 2009 - BILHVAX - A VACCINE AGAINST BILHARZIOSE, A world first!
- 20 April 2009 - 300.000 morts évitées grâce au vaccin liégeois!
- 22 April 2009 - Eurogentec, la société de biotechnologie située au Sart Tilman, produit et produira le vaccin contre la bilharziose.
Annexes
Articles connexes
Bibliographie
- 1. Warren, K. S., E. O. Domingo, and R. B. Cowan. 1967. Granuloma formation around schistosome eggs as a manifestation of delayed hypersensitivity. Am J Pathol 51:735.
- 2. Mosmann, T. R., H. Cherwinski, M. W. Bond, M. A. Giedlin, and R. L. Coffman. 1986. Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J Immunol 136:2348.
- 3. Angeli, V., C. Faveeuw, O. Roye, J. Fontaine, E. Teissier, A. Capron, I. Wolowczuk, M. Capron, and F. Trottein. 2001. Role of the parasite-derived prostaglandin D2 in the inhibition of epidermal Langerhans cell migration during schistosomiasis infection. J Exp Med 193:1135.
- 4. Herve, M., V. Angeli, E. Pinzar, R. Wintjens, C. Faveeuw, S. Narumiya, A. Capron, Y. Urade, M. Capron, G. Riveau, and F. Trottein. 2003. Pivotal roles of the parasite PGD2 synthase and of the host D prostanoid receptor 1 in schistosome immune evasion. Eur J Immunol 33:2764.


Wikimedia Foundation. 2010.