- Autophagie
-
Autophagie, du grec αυτο (soi-même) et φαγειν (manger), dégradation d'une partie du cytoplasme de la cellule par ses propres lysosomes. Synonyme : autolyse. (λυσις = dissolution) ou autophagocytose.
L'histolyse (ἱστος = tissu) est le même processus, mais intervenant lors d'une métamorphose, il est utile à l'organisme. Par exemple, les amphibiens. L'histolyse est également présente au cours du développement embryonnaire ou en fin de lactation.
Le terme « autophagie » (se manger soi-même) regroupe plusieurs voies de dégradation lysosomale des constituants cellulaires, essentielles à l’homéostasie cellulaire. Il existe trois types différents d’autophagies :
- la microautophagie
- l’autophagie réalisée par des protéines chaperonnes
- la macroautophagie : c'est la forme principale.
La macroautophagie, appelée couramment autophagie, est un mécanisme permettant à la cellule de digérer une partie de son contenu, que ce soit du cytoplasme, des protéines ou des organites cellulaires. C’est la seule voie qui puisse dégrader massivement des macromolécules et des organites, c’est une voie de dégradation alternative à celle du protéasome.
Sommaire
Historique
La découverte des gènes Atg (autophagy related genes), au milieu des années 1990 chez la levure a été importante, non seulement pour la dissection en termes moléculaires de la macroautophagie, mais aussi pour comprendre son importance en physiologie et physiopathologie. On distingue plus de 30 gènes Atg dont plus de la moitié sont impliqués dans l’autophagie. Ces gènes sont présents aussi bien chez la levure que chez les cellules de mammifères.
Mode de fonctionnement
Sur le plan cellulaire, la macroautophagie débute par la formation d’une vacuole qui s’allonge et séquestre de façon non sélective des constituants du cytoplasme. L’autophagosome ainsi formé est délimité par une double membrane phospholipidique. Plusieurs compartiments cellulaires (réticulum endoplasmique, appareil de Golgi et réseau trans-golgien) et la membrane plasmique participeraient probablement à la formation de l’autophagosome. Une quinzaine de protéines Atg sont nécessaires à sa biogenèse. À l’exception d’Atg9, ces protéines ne possèdent pas de domaine transmembranaire. Les protéines Atg, recrutées dans le cytoplasme, se complexent les unes aux autres et s’associent de façon transitoire avec la membrane pré-autophagosomale et à celle de l’autophagosome.
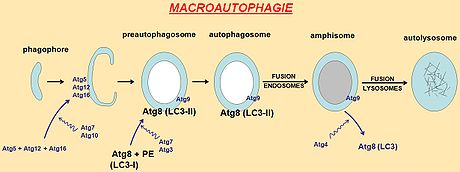 Mécanisme de l'autophagie
Mécanisme de l'autophagie
Déclenchement de l'autophagie
L’autophagie peut être stimulée en conditions de stress, telles que la carence en nutriments, l’absence de facteurs de croissance ou l’hypoxie.
La protéine mTOR (mammalian target of rapamycin) joue un rôle-clé dans l’intégration de ces signaux et la régulation de l’autophagie. Lorsqu’elle est activée, mTor inhibe la voie de l’autophagie, mais son inactivation (suite à une carence en nutriments, par exemple) permet de lever l'inhibition et donc de lancer l’autophagie. Cette enzyme est la cible de la rapamycine (une drogue classiquement utilisé pour induire l’autophagie).
Une autre voie déclenchant l'autophagie fait intervenir le complexe Atg6 (aussi appelé Beclin 1)/PI3 kinase de classe III. Ce complexe participe à l’induction de l’autophagie, à l’incurvation du pré-autophagosome et à la formation de l’autophagosome.
Le déroulement ultérieur du processus autophagique repose essentiellement sur deux systèmes de conjugaison analogues aux systèmes d'ubiquitination des protéines :
- le premier conjugué, formé des protéines Atg5-Atg12, permet le recrutement du deuxième complexe sur l'autophagosome en formation, puis est recyclé vers le cytosol.
- le deuxième conjugué résulte de la conjugaison de la protéine Atg8 (MAP-LC3 chez les mammifères) à la phosphatidyléthanolamine (PE) et s’incorpore dans la membrane du pré-autophagosome. LC3 existe donc sous forme cytosolique (LC3-I) ou associé au phagosome (LC3-II = Atg8-PE). LC3-II pourrait jouer une fonction structurale permettant l’élongation et la formation de l’autophagosome mature, et constitue un marqueur cellulaire d'identification des autophagosomes.
Autres rôles physiologiques de l'autophagie
L’autophagie joue un rôle important dans :
- le maintien de l’homéostasie car elle permet l’élimination et le remplacement continuel des protéines et des organites non fonctionnels.
- l’adaptation et la survie des cellules soumises à des conditions de stress.
- l’immunité innée à l’échelle de la cellule car elle permet d’éliminer des pathogènes intracellulaires.
- l’immunité adaptative car la dégradation des protéines par autophagie génère des peptides qui seront ensuite présentés sur le complexe majeur d'histocompatibilité (CMH).
Rôle anti-tumoral
L’identification de molécules impliquées dans le contrôle et l’exécution de l’autophagie a mis en lumière une étroite relation entre l’autophagie et la progression tumorale. On a remarqué que les cellules cancéreuses présentaient généralement une capacité autophagique plus réduite que les cellules normales. Le rôle anti-tumoral de l’autophagie est suggéré par son implication dans la réduction de l’instabilité chromosomique, de la prolifération et de l’inflammation des cellules tumorales. L’autophagie peut aussi être un mécanisme pro-tumoral en permettant la survie des cellules tumorales exposées à des variations de leur microenvironnement (hypoxie, carence nutritionnelle…). Le processus autophagique est induit en réponse à divers traitements anti cancéreux. Cette réponse peut s'avérer être un mécanisme permettant la survie des cellules, ou favorisant et amplifiant la mort induite par ces traitements.
Rôle protecteur dans les maladies neurodégénératives
Une caractéristique commune à de nombreuses maladies neurodégénératives est l’accumulation de protéines mutées ou toxiques et leur agrégation dans le cytoplasme. Ces maladies sont désignées sous le terme de « protéinopathies » et comprennent notamment la maladie d’Huntington, la maladie d’Alzheimer, la maladie de Machado-Joseph (SCA-3)…
Ces agrégats de protéines anormales se formeraient, par exemple, suite à une diminution de l’activité du protéasome. Leur présence entraîne une augmentation de cette autophagie induite qui, dans les premiers stades de la maladie, est assez efficace pour assurer leur élimination. Cependant, à un stade plus avancé, l’autophagie induite n’est plus suffisante, et des inclusions protéiques se forment dans les cellules ; la stimulation de l’autophagie (par un agent pharmacologique) pourrait s’avérer efficace pour éliminer ces inclusions et ainsi lutter contre la neurodégénérescence.
Dans le cas de la maladie de Machado-Joseph (SCA-3, Spino Cerebellar Ataxia - 3), cette technique de stimulation de l'autophagie est actuellement étudiée par les équipes de Luis Pereira de Almeida, Center for Neurosciences and Cell Biology, University of Coimbra, Largo Marquês de Pombal, 3004-517 Coimbra, Portugal[1]Notes et références
- (en)Overexpression of the autophagic beclin-1 protein clears mutant ataxin-3 and alleviates Machado–Joseph disease, 2011. Mis en ligne le February 1, 2011
Voir aussi
Liens externes
- HADb, une base de donnée dédiée à l'autophagie
- L'autophagie : un nouveau concept en cancérologie, magazine Bull Cancer, volume 95, n°1, p43-50
- L'autophagie remplaçant de luxe du protéasome, magazine Médécine Sciences, volume 24, n°1, p 19-21
- Les deux visages de l'autophagie dans le système nerveux, magazine Médecine Sciences, volume 25, n°4, pp. 383-388
 Portail de la biologie cellulaire et moléculaire
Portail de la biologie cellulaire et moléculaire
Wikimedia Foundation. 2010.