- Pteridospermatophyte
-
Pteridospermatophyta
Pteridospermatophyta †

Fossile de ptéridosperme Classification classique Règne Plantae Division Pteridospermatophyta †
— auteur incomplet —, date à préciserParcourez la biologie sur Wikipédia : 





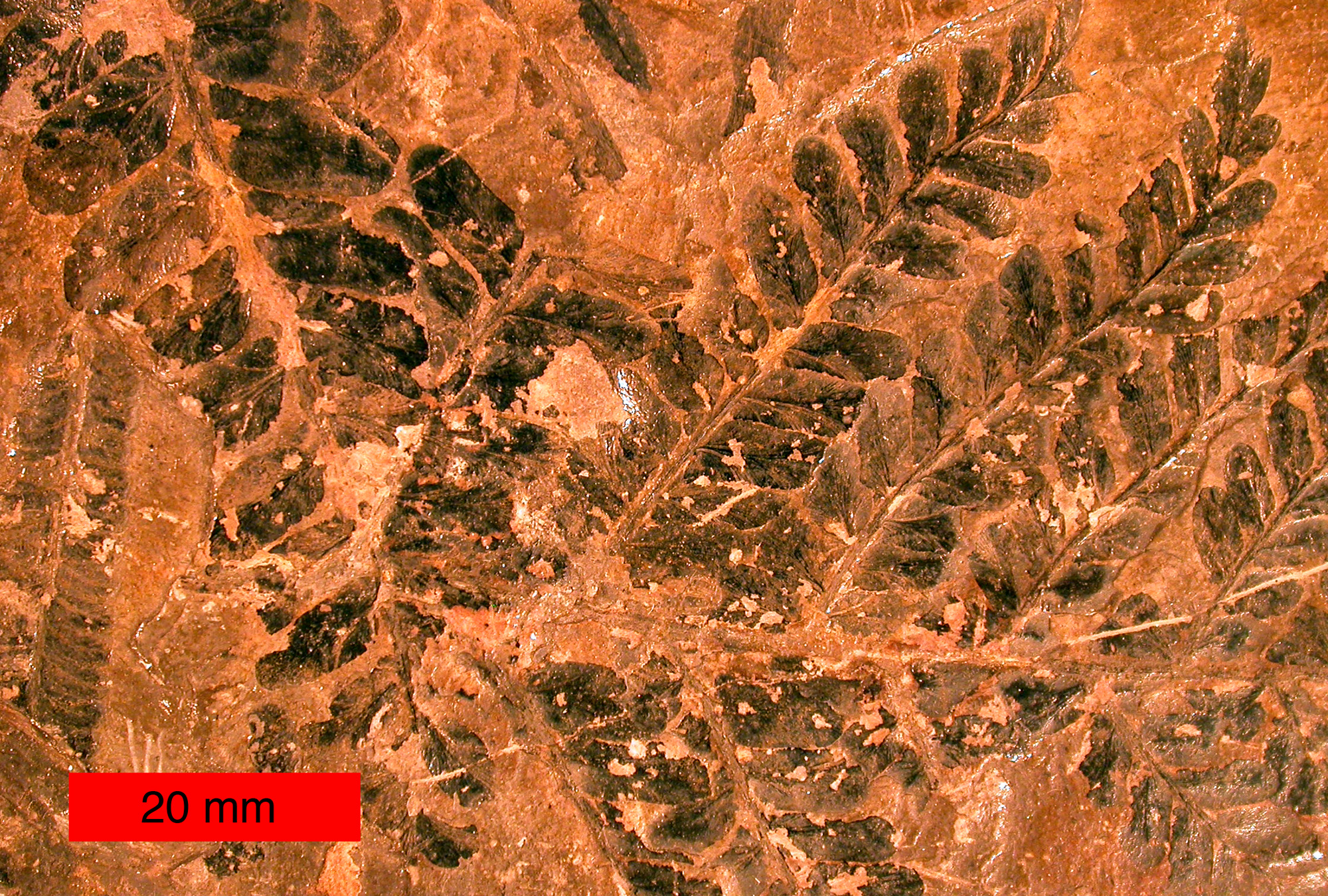
Les Pteridospermatophyta, aussi appelées ptéridospermes ou fougères à graines, sont un groupe de spermaphytes. Les membres de cette division apparaissent au cours du Dévonien tardif, puis déclinèrent il y a 200 million d'années, et disparurent au Crétacé. Les ptéridospermatophytes n'ont pas encore acquis de position précise dans la systématique et semblent être un taxon paraphylétique. Certaines semblent être affiliées aux gymnospermes, d'autres ont été proposées comme des ancêtres possibles des angiospermes.
Les différents groupes de ptéridospermes [1] ordre âge distribution Buteoxylales Carbonifère inférieur Europe Calamopityales Carbonifère inférieur Amérique du Nord, Europe Lyginoptéridales Carbonifère Amérique du Nord, Europe Médullosales Carbonifère-Permien Amérique du Nord, Europe Callistophytales Carbonifère supérieur Amérique du Nord, Europe Glossopteridales Permien-Trias Australie, Antarctique, Inde, Amérique du Sud, Afrique Gigantopteridales Permien Amérique du Nord, Asie Peltaspermales Permien-Trias Afrique du Sud, Europe, Groenland, Russie Corystospermales Trias-Crétacé Amérique du Nord, Asie Caytoniales Trias-Crétacé Amérique du Nord, Asie, Europe Sommaire
Les premières ptéridospermes
Les plus anciennes ptéridospermes connues sont datées du Dévonien supérieur. La plupart sont représentées seulement par des pré-ovules, des ovules, des cupules. La plus ancienne ptéridosperme reconstruite à ce jour est Elkinsia[2], du Famennien des États-Unis. Elkinsia était une plante non-ramifiée de petite stature qui portait de larges frondes dichotomes. Sa tige comprenait une protostèle centrale, un peu de bois et un cortex fibreux. La base des pétioles a une anatomie en C ou en W. Les frondes fertiles portaient soit des cupules contenant chacune plusieurs ovules (structure 'femelle') soit des organes polliniques (structure 'mâles').
Si on assiste au Dévonien supérieur à une diversification des ptéridospermes, la découverte récente du proto-ovule Runcaria [3]dans le Givétien (Dévonien moyen) de Belgique indique que l'origine du groupe est vraisemblablement plus ancienne.
Butéoxylales
Ce groupe a été établi par Barnard et Long en 1973. Il ne comprend que deux genres, basés sur des tiges du Carbonifère inférieur d’Europe: Buteoxylon et Triradioxylon. Une des caractéristiques du groupe est la forme en T de la base du pétiole. Le feuillage serait peut-être de type Triphyllopteris. Les parties fertiles sont inconnues.
Calamopityales
Les Calamopityales sont connues au Carbonifère inférieur en Europe et aux États-Unis. Leur système vasculaire primaire varie d'une protostèle (pas de moelle) à une eustèle. Leur bois est manoxylique (trachéides larges et grands rayons). Elles portaient de grandes frondes et devaient ressembler à des lianes. Cinq genres de tiges sont décrits dont Calamopitys et Stenomyelon.Les parties fertiles sont inconnues mais il a été suggéré que Stenomyelon portaient des graines de type Lyrasperma.
Bien que les affinités systématiques de ce groupe soient incertaines, il est important car il montre différents stades de l'évolution d'une eustèle. De plus il est possible que les médullosales aient évolué à partir de ce groupe.
Lyginoptéridales
Les Lyginoptéridales étaient de petites plantes graciles dont les genres les plus connus sont Heterangium et Lyginopteris. Ce groupe du Carbonifère a une importance historique particulière. En 1904, F. Oliver et D.H. Scott ont montré la connexion entre plusieurs organes connus séparément (tige, feuillage et graines) ce qui a permis de reconstruire Lyginopteris et a mené au concept de ptéridospermes.
Tige. Les tiges peuvent atteindre 4 cm de diamètre. Leur partie centrale est occupée par une eustèle avec une moelle parenchymateuse contenant des cellules sclérifiées. le bois est composé de trachéides et de grands rayons. Il y a un peu de périderme. La partie la plus externe de la tige est composée d'une alternance de bandes de parenchyme et de fibres sclérifiées. L'épiderme des jeunes tiges est couvert de glandes multicellulaires. Ce sont ces glandes présentes sur tous les organes de la plantes sauf les racines qui ont permis la reconstruction de la plante entière par Oliver et Scott.
Racines. Les racines sont adventives et atteignent 7 mm de diamètre.
Feuillage. La base des pétioles de Lyginopteris est en forme de V ou de W. Les frondes sont disposées en hélice sur la tige. Leur pinules correspondent au genre de forme Sphenopteris.
Reproduction. Lyginopteris portait des graines de type Lagenostoma (appelées Lagenospermum en compression). Lagenostoma a une forme en ellipse et mesure quelques millimètres de diamètre. Chaque ovule était porté dans une cupule en forme de tulipe. La partie apicale du nucelle est allongée en une structure particulière (lagénostome). Les organes polliniques de Lyginopteris ne sont pas connus. Le genre Crossotheca a longtemps été un candidat mais il semble qu'il corresponde en fait à des organes polliniques de fougères.
Certains genres moins bien connus du début du Carbonifère sont aussi placés dans les Lyginoptéridales, principalement en raison de la forme de leur pétiole. Certains de ces genres correspondent à de petites plantes (ex.: Tristichia) , d'autres à des arbres (ex.: Pitus).
Médullosales
Les Médullosales poussaient dans les plaines tropicales ou subtropicales. Certaines sont reconstruites comme de petits arbres qui pouvaient atteindre plusieurs mètres de haut. Le groupe atteint son maximum de diversité au Carbonifère supérieur.
Tige. La tige des Médullosales est caractérisée par une eustèle particulière où le cambium entoure individuellement chaque faisceau de xylème primaire ce qui donne l’impression qu’il y a plusieurs stèles.
Racines. Les racines sont adventives. Les plus gros spécimens ont 2.5 cm de diamètre.
Feuillage. les feuilles des Médullosales étaient de grandes frondes, généralement bifurquées et portées en spirale. Le feuillage retrouvé en compression est décrit notamment sous les noms Neuropteris et Alethopteris.
Reproduction. les Médullosales avaient de grosses graines charnues de plusieurs cm de diamètre et des grains de pollen très gros (jusqu'à 600µm). Ceci suggère une interaction avec des animaux disperseurs et pollinisateurs. Les ovules sont appelés Trigonocarpus ou Pachytesta selon qu’ils sont retrouvés en compression ou anatomiquement préservés. il n'étaient pas inclus dans une cupule. les organes polliniques pouvaient atteindre plusieurs cm de diamètres.
Callistophytales
Les Callistophytales sont représentées par le genre Callistophyton, découvert dans les années 1970. Callistophyton était un petit buisson ou une liane qui portait de grandes frondes.
Tige. Les plus grosses tiges ont 3 cm de diamètre. La partie centrale est une eustèle avec une moelle parenchymateuse. Le bois comprend des trachéides et des rayons bisériés. Un peu de périderme se met en place dans les tiges âgées.
Feuillage. Les frondes sont portées en spirale et atteignent 30 cm de long. Leurs pinnules peuvent porter des poils et ressemblent au genre de forme Sphenopteris.
Reproduction. Les graines de Callistophyton sont appelées Callospermarion. Elles pouvaient atteindre 5 mm de long. les organes polliniques sont connus ainsi que la structure du pollen qui est d'apparence relativement moderne.
Glossoptéridales
La présence de feuilles de Glossoptéridales (Glossopteris) dans des roches d'âge Carbonifère à Trias d'Australie, Afrique, Amérique du Sud, Antarctique et Inde a joué un rôle important dans la théorie de la dérive des continents en montrant que ces différentes zones avaient autrefois constitué une seule masse continentale: le Gondwana.
Les Glossoptéridales sont reconstruites comme des buissons ou des arbres. Elles avaient un rôle écologique important et dominaient certains écosystèmes du Permien. Plus de 200 espèces ont été décrites à travers le monde mais ce sont des espèces de forme et on ne sait pas à ce jour quelle est la part de variation intraspécifique dans ce groupe.
Tiges/troncs. les Glossoptéridales avaient des tiges avec une eustèle et un bois picnoxylique, c'est à dire avec des trachéides étroites et de petits rayons.
Racines. Les racines de Glossoptéridales sont incluses dans le genre Vertebraria, nom donné en raison de leur ressemblance avec des vertèbres quand elles sont conservées en compression. Le bois des racines est séparé en secteurs par des zones vides qui pourraient avoir un rôle aérifère.
Feuillage. Les feuilles de Glossoptéridales sont simples, très longues et ont un aspect rubanné. Elles étaient portées en hélice. On considère que ces feuilles étaient caduques en raison de l'alternance annuelle de sédiments contenant beaucoup de feuilles et de sédiments sans feuilles.
Reproduction. Plus de 40 espèces d'organes reproducteurs attribués aux Glossoptéridales ont été décrits. La plupart sont conservés en compression et le détail de leur anatomie est mal connu.
Les paléobotanistes on suggéré plusieurs affinités pour les Glossoptéridales. En particulier, certains comme E. Plumstead (1956) ou plus récemment G. Retallack et D. Dilcher (1981) ont défendu une relation avec les angiospermes. Les analyses cladistiques les plus récentes les placent soient dans le clade des anthophytes, qui contient aussi les angiospermes [4], soit dans les ptéridospermes[5].
Gigantoptéridales
Cet ordre comprend des plantes connues en Asie et en Amérique du Nord. L'ordre est essentiellement basé sur des feuilles en compression. Ces feuilles peuvent atteindre plus de 30 cm de long. S. Meyen, qui a crée le groupe sous le nom "gigantonomiales" en 1987, considérait que ces plantes étaient affiliées notamment aux Callistophytales, Caytoniales et Ginkgoales.
Peltaspermales
Ce groupe est basé sur des feuillage en compression et des éléments fertiles associés pollen et graines). Le principal genre de feuillage est Lepidopteris, qui comprend des formes généralement bi-pennées. Le genre Permien Callipteris a aussi été rangé dans cet ordre. Callipteris est un marqueur bio-stratigraphique de la limite Carbonifère-Permien en Europe. Les ovules étaient portés en cercle sur la face inférieure d'une mégasporophylle en forme d'ombrelle.
Corystospermales
Les Corystospermales forment un petit groupe de plantes ligneuses reconstruites à partir d'organes toujours trouvés en association.
Tige/Tronc. Les plus gros spécimens inclus dans le groupe sont des troncs de 80 cm de large (Rhexoxylon) retrouvés en Amérique du Sud, Afrique du Sud et Antarctique. La partie centrale des tiges contient une moelle avec des groupes de cellules sclérifiées. Le xylème secondaire (bois) se développe de part et d'autre du xylème primaire (=centrifuge et centripète).
Feuillage. Le feuillage le plus courant associé aux Corystospermales est Dicroidium. C'est une fronde dont le rachis est bifurqué. les différentes espèces sont pennées à tripennées.
Reproduction. Les branches fertiles portent des cupules recourbées. Chaque cupule contient un ou plusieurs ovules. Les organes polliniques sont des microsporophylles portée sur des axes courts.
Caytoniales
Les Caytoniales sont reconstruites comme de petits arbres. Elles ont été initialement décrites comme un nouveau groupe d'angiospermes par H. Thomas (1925) et ont souvent été par la suite considérées comme des ancêtres possibles des angiospermes.
Feuillage. Les feuilles des Caytoniales appartiennent au genre Sagenopteris. Ce sont des feuilles composées palmées ayant 3 à 6 folioles.
Reproduction. La structure qui porte les ovules est appelée Caytonia. C'est un axe qui porte sur des pédoncules des cupules recourbées contenant plusieurs ovules (8 à 30). Ces cupules multiovulées ont été initialement décrites comme des fruits d'Angiospermes. Les organes polliniques atteignaient 1 cm de long et contenaient de petits grains de pollen (25-30µm).
Liens externes
Pteridosperms or seed ferns,UNIVERSITY MÜNSTER
Virtual paleobotany lab: Medullosa
Virtual paleobotany lab: Callistophytales, Caytoniales, Glossopteridales Corystospermales et Peltaspermales Pteridospermopsida Links
Notes et références
- ↑ TAYLOR, T. N., et TAYLOR, E. L. 1993. The biology and evolution of fossil plants. Prentice Hall, Englewood Cliffs, New Jersey, USA.
- ↑ SERBET, R., et ROTHWELL, G. W. 1992. Characterizing the most primitive seed ferns. I. A reconstruction of Elkinsia polymorpha. Int. J. Plant. Sci. 153: 602-621.
- ↑ GERRIENNE, P., MEYER-BERTHAUD, B., FAIRON-DEMARET, M., STREEL, M., et STEEMANS, P. 2004. Runcaria, a Middle Devonian Seed Plant Precursor. Science 306: 856-858.
- ↑ DOYLE, J. A., et DONOGHUE, M. J. 1986. Seed Plant Phylogeny and the Origin of Angiosperms: An Experimental Cladistic Approach. Bot. Rev. 52: 321-431.
- ↑ ROTHWELL, G. W., et SERBET, R. 1994. Lygnophyte Phylogeny and the Evolution of Spermatophytes: A Numerical Cladistic Analysis. Syst. Bot. 19: 443-482.
Autres références générales sur les ptéridospermes utilisées:
GALTIER, J. 1988. Morphology and Phylogenetic Relationships of Early Pteridosperms. In C. B. Beck [ed.], Origin and Evolution of Gymnosperms. Columbia University Press, New-York. 135-176.
DUNN, M. T. 2006. A review of permineralised seed ferns stems of the Upper Paleozoic. Journal of the Torrey Botanical Society 133: 20-32.
 Portail de la botanique
Portail de la botanique
Catégories : Plante préhistorique | Plante
Wikimedia Foundation. 2010.