- Adenoviridae
-
 Adénovirus
Adénovirus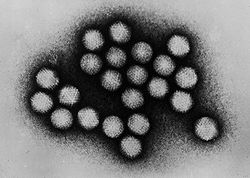
Des adénovirus au
microscope électronique en transmissionClassification des virus Type Virus Groupe Groupe I Famille Adenoviridae Adenoviridae (ou Adénovirus) est une famille de virus qui regroupe une centaine de variétés, dont une quarantaine environ peut infecter l'Homme. C'est en 1953 que ceux-ci ont été mis en évidence par Wallace P. Rowe à partir de fragments d'amygdale.
Sommaire
Morphologie
Les adénovirus sont des virus possédant de l'ADN double brin linéaire (30 000 à 38 000 pb), les deux brins d'ADN possèdent à leurs extrémités des séquences répétées inversées qui permettent aux molécules monocaténaires (simple brin) de se circulariser (la protéine p55 permet la circularisation). Les adénovirus sont des particules d'un diamètre de 60 à 90 nm, sans enveloppe, à capside icosaédrique formée de 252 capsomères (240 hexons et 12 pentons). Les capsomères situés aux sommets de l'icosaèdre sont des pentons prolongés par une fibre de longueur variable et terminée par l'antigène Y, responsable de la propriété d'hémagglutination.
Protéines de la capside
L’hexon est constitué de 3 chaînes de polypeptide II. Les polypeptides VI, VIII et IX forment les liaisons entre les capsomères. La base du penton est constituée de 5 polypeptides III et la fibre de 3 polypeptides IV.
Protéines de core
Les polypeptides VII sont des analogues d’histones, ils se complexent avec l'ADN. Les polypeptides V entourent le complexe ADN-VII et le relie à la capside.
Cycle Viral
Le cycle viral complet dure de 30 à 36 heures et se termine par la libération d'environ 10 000 particules virales après lyse de la cellule infectée. Ce rendement, très impressionnant, varie cependant d'un sérotype à un autre et du type cellulaire infecté. Le cycle se divise en trois étapes, une phase précoce correspondant à l'attachement et à l'entrée du virus, suivi d'une phase de réplication de l'ADN viral et de production des protéines virales et d'une phase tardive d'assemblage et de libération des virions.
Phases précoces
Le cycle viral débute par l'attachement de la particule virale à la cellule hôte et à son internalisation dans cette dernière.
La phase précoce peut être décrite par les différentes étapes suivantes:
- Adsorption : la fibre du virion est responsable de l’attachement cellulaire.
- Pénétration par endocytose.
- Décapsidation
- perte des pentons près de la membrane plasmique
- perte des hexons près de la membrane nucléaire
- l'ADN pénètre dans le noyau
- Expression des gènes précoces
L'entrée du virus
L'entrée de l'adénovirus a beaucoup été étudiée pour les sérotypes 2 et 5 (appartement à la famille C des adénovirus humains). Cette entrée est séquentielle et repose sur l'interaction avec des protéines cellulaires. Pour les adénovirus de sérotype 2 et 5, la fibre de la particule virale est responsable de l'attachement à la cellule hôte via le récepteur CAR (Coxsackie and Adenovirus Receptor). Ce récepteur est commun aux virus Coxsackie B3 de la famille des Picornaviridae et aux adénovirus de la famille A, C, E et éventuellement D. CAR est une glycoprotéine transmembranaire de 46kDA appartenant à la superfamille des immunoglobulines. Cette protéine est exprimée sur un large spectre de type cellulaire expliquant le large tropisme des adénovirus[1]. La protéine CAR est impliquée dans l'adhésion intercellulaire au niveau basolatéral des jonctions serrées des épithéliums[2].
Tous les sérotypes, à l'exception de ceux du sous-groupe B, ont la capacité de se lier à CAR, mais tous n'utilisent pas préférentiellement ce récepteur. Par exemple, l'adénovirus 37, appartenant au sous-groupe D, peut se lier à CAR mais utilise le récepteur CD46 pour infecter les cellules[3]
Même si l'expression de CAR participe au tropisme de l'Adénovirus in vitro [4], il n'y a cependant pas de corrélation directe entre le tropisme du virus pour des tissus exprimant fortement CAR in vivo chez la souris[5]. Ceci sous-entend la présence de récepteurs alternatifs qui seraient préférentiellement utilisés in vivo.
L'attachement de la tête de la fibre au récepteur CAR conduit à un changement de conformation permettant d'exposer le motif RGD (Arginine - Glycine - Acide Aspartique)porté sur une boucle à la surface de chaque monomère de la base de penton. Ce motif va ensuite interagir avec les intégrines cellulaires AlphaV Béta1, AlphaV Béta3, AlphaV Béta5, Alpha5 Béta1 et Alpha3 Béta1.
L'exposition des 5 motifs RGD portés par la base de penton (molécule pentamérique) permet l'association simultanée de plusieurs molécules d'intégrines. ce recrutement va initier l'entrée par endocytose dans la cellule.
Transport au noyau
Le virus est internalisé par la voie des vésicules à clathrine puis passe dans les endosomes. Le mécanisme d'échappement de l'endosome n'est pas encore clairement élucidé. Néanmoins, suite à l'acidification des endosomes précoces, les protéines IIIa et VIII se dissocient.
Activation des gènes viraux
Les gènes précoces sont divisés en 4 groupes : E1 (A et B), E2 (A et B), E3 et E4.
Expression de E1A. E1A code des protéines transactivatrices de la transcription qui agissent au niveau des promoteurs viraux et promoteurs de gènes cellulaires endogènes ainsi que pour des protéines d’autorégulation. Il active l’entrée en phase S. Une protéine de E1A active également le gène qui code la protéine p53 qui induit les mécanismes d’apoptose (p53 est un facteur de transcription qui va activer l'expression du gène de Bax).
Expression de E4. Cette région code 7 protéines (minimum) toxiques pour la cellule qui vont induire l’apoptose indépendante de p53. La protéine E4orf6 bloque l’accumulation de p53. E4orf4 se lie à une protéine phosphatase p2a, entraînant la déphosphorylation d’activateurs de E1A et autres facteurs de transcription, aboutissant à la diminution de l’expression de E4 (autorégulation).
Expression de E3. Son expression dépend d’activateurs synthétisés lors de l’expression de E1A et E4. La région E3 permet à la cellule infectée d’échapper au système immunitaire, elle contrecarre les défenses innées. E3 code une protéine qui s’associe aux molécules CMH de classe I. Cette protéine empêche que ces molécules soient exprimées à la surface de la cellules. E3 code aussi plusieurs protéines qui interfèrent avec l’inflammation (Tumor Necrosis Factor). Ces protéines de E3 viennent empêcher la fixation du TNF sur son récepteur. Une protéine de la région E3 produite très tardivement jouerait un rôle dans la destruction de la cellule.
Expression de E1B. E1B a une fonction anti-apoptotique. E1B s’exprime quasi simultanément avec E3 et code la protéine p55k qui se lie a p53 et l’inactive, et pour la protéine p19k, un équivalent fonctionnel de Bcl-2 (la protéine Bcl2 empêche l’homodimérisation de Bax, et l’homodimérisation de Bax permet l’activation de la caspase qui active à son tour l’endocaspase qui digère l’ADN). E1B bloque également l’apoptose induite par les produits d’E4 (nécessité d’un délai : le virus doit avoir du temps pour synthétiser ses particules virales). E1B19k serait inhibée tardivement par la protéine d’E3 qui joue un rôle dans la destruction de la cellule, la protéine p19k étant inhibée, Bax est actif et induit l'apoptose.
Expression de E2. Son expression, comme celle de E3, dépend d’activateurs produits par E1A et E4. Cette région E2 est impliquée dans la réplication de l’ADN viral. E2A code une DNA binding protein (DBP) p72 qui recouvre l’ADN monocaténaire néosynthétisé et le protège de l'activité des nucléases cellulaires. E2B code une protéine terminale p55 liée de façon covalente aux extrémités 5’ de l’ADN bicaténaire et sert d’amorce à la réplication.
Réplication
Un des brins va être copié de façon continue de 5’vers 3’. L’autre brin est déplacé et recouvert par une protéine : la DBP (DNA Binding Protein) p72. Plusieurs fourches de réplication peuvent se suivre. On obtient des brins bicaténaires et monocaténaires. Ces derniers vont se circulariser grâce à des séquences répétées inversées a et a'.
Phases tardives
Les gènes tardifs sont regroupés sous un seul opéron et se décomposent en 3 familles : L1, L2 et L3. La transcription s’effectue à partir de ce promoteur unique. Le regroupement de plusieurs gènes sous la régulation d’un seul promoteur permet une expression simultanée des protéines tardives. Les ARNm tardifs vont être traduits en protéines du core, de la capside et protéines des fibres.
L’assemblage se fait dans le noyau, donc les protéines de structure doivent migrer dans le noyau. Les produits des régions d'E3 et d'E4 induisent l'apoptose, la cellule est lysée et les virions libérés.
Pouvoir pathogène
Ils sont capables d'infecter des cellules à division lente et se multiplient dans l'œil, l'appareil respiratoire et l'appareil digestif. Le pouvoir pathogène des adénovirus s'exerce principalement sur l'appareil respiratoire. La transmission peut être :
- directe : par voie aérienne
- indirecte : conjonctivite des piscines
Ils sont excrétés par les selles de façon prolongée. Ils sont responsables de pharyngites et de pneumonies, mais aussi de conjonctivites. Certains sérotypes (40, 41), lorsqu'ils sont déglutis, peuvent causer des gastro-entérites (1 à 9% des gastro-entérites chez les enfants).
Les adénovirus peuvent aussi provoquer des dermatoses erythémateuses (Acrodermatite papuleuse infantile de Gianotti-Crosti) Les corps apoptotiques contenant des particules virales sont transmis aux cellules adjacentes par endocytose. On estime que 50% des infections à adenovirus sont inapparentes.
Les anticorps apparaissent dès l'enfance, cependant, la multitude des types (plus de 42 chez l'Homme) fait que l'on peut avoir des gastro-entérites à répétition. A l'age de 4 ans, 85% des enfants ont développé une immunité contre l'avénovirus de type entérique[6].
Liste des genres
- Aviadenovirus
- Atadenovirus
- Mastadenovirus
- Siadenovirus
Notes et références
- Howitt J, Bewley MC, Graziano V, Flanagan JM and Freimuth P. J Biol Chem, 2003, 278: 26208-15
- Coyne CB, Bergelson JM. [[]] 2005 Apr 25;57(6):869-82
- Iacobelli-Martinez M, Nepomuceno RR, Connolly J, Nemerow GR. J Virol.2005 Sep;79(17):11259-68.
- Freimuth P. J Virol.1996 Jun;70(6):4081-5.
- Fechner H, Haack A, Wang H, Wang X, Eizema K, Pauschinger M, Schoemaker R, Veghel R, Houtsmuller A, Schultheiss HP, Lamers J, Poller W.Gene Ther. 1999 Sep;6(9):1520-35
- FDA, bad bug book, 2011
Bibliographie
- P. Rowe, Occurrence of Adenovirus-SV40 Hybrids Among Monkey Kidney Cell Adapted Strains of Adenovirus, Proceedings of the Society for Experimental Biology and Medicine, 1966, Vol. 122, pp. 214-218.
- Marc Vasseur, Les Virus Oncogènes, Hermann, 1997.
Liens externes
- Réplication des virus a ADN Université de Caroline du Sud
Wikimedia Foundation. 2010.