- Sauropode
-
Sauropoda
Sauropodes

Squelette remonté de Diplodocus carnegiei Classification classique Règne Animalia Embranchement Chordata Sous-embr. Vertebrata Classe Reptilia Super-ordre Dinosauria Ordre Saurischia Sous-ordre Sauropodomorpha Infra-ordre Sauropoda
Marsh, 1878Familles de rang inférieur - Antarctosauridae
- Argyrosauridae
- Brachiosauridae
- Cetiosauridae
- Camarasauridae
- Dicraeosauridae
- Diplodocidae
- Euhelopodidae
- Nemegtosauridae
- Rebbachisauridae
- Saltasauridae
- Vulcanodontidae
 Retrouvez ce taxon sur Wikispecies
Retrouvez ce taxon sur Wikispecies
D'autres documents multimédia
sont disponibles sur CommonsParcourez la biologie sur Wikipédia : 





Les sauropodes sont un infra-ordre de dinosaures, quadrupèdes herbivores, appartenant au sous-ordre des sauropodomorphes. Ils vécurent entre le Jurassique moyen et le Crétacé supérieur et remplacèrent les prosauropodes sur l'ensemble de la planète. On compte parmi les sauropodes les plus longs et les plus imposants dinosaures (brachiosaure, diplodocus sauroposeidon), et par conséquent les plus grands animaux qui aient vécu sur terre. Voici une liste partielle des familles :
- cétiosauridés : 18 m de long, tête plate et cou relativement court (tous les continents)
- brachiosauridés : voir brachiosaurus
- camarasauridés : 12 à 18 m de long, crânes massifs et vertèbres creuses (Amérique du Nord, Europe et Asie)
- diplodocidés : voir diplodocus.
- titanosauridés : 9 à 21 m de long (continents du sud)
C'est O.C. Marsh, en 1878, qui forgea le mot sauropode, du grec « pied de lézard »[1].
Les découvertes de fossiles complets de sauropodes sont rares. De nombreuses espèces, et plus spécialement les plus grandes, ne sont connues qu'à partir d'ossements isolés et désarticulés. Beaucoup de spécimens quasi complets n'ont pas de tête, d'appendices caudaux ou de membres.
Sommaire
Description
Les sauropodes étaient des quadrupèdes herbivores, avec habituellement un long cou et des dents en forme de spatule : base large, col étroit. Ils avaient de petites têtes, des corps énormes et, en général, de longues queues. Leurs pattes épaisses se terminaient par de gros pieds à cinq orteils, bien que trois seulement fussent pourvu de griffes.
Dimensions
 Squelette de Brachiosaurus brancai, le plus grand sauropode connu à partir de spécimens quasi complets.
Squelette de Brachiosaurus brancai, le plus grand sauropode connu à partir de spécimens quasi complets.
La caractéristique la plus évidente des sauropodes était leur taille. Même les sauropodes nains comme l'Europasaurus (peut-être de 5 à 6 mètres) faisaient partie des plus gros animaux de leur écosystème. Leurs seuls véritables concurrents en termes de taille sont les rorquals. Mais, à la différence des baleines, les sauropodes vivaient tous sur la terre ferme. Quelques uns, tels les diplodocidés avaient un port de tête bas, tandis que d'autres, comme le Camarasaurus, avaient un port de tête haut.
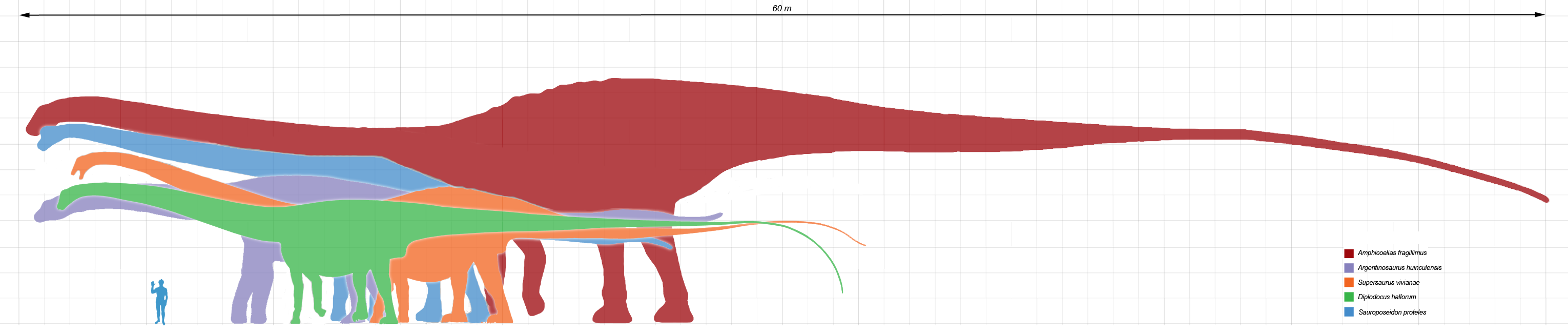
 Comparaison de la taille de quelques dinosaures sauropodes géants.
Comparaison de la taille de quelques dinosaures sauropodes géants.Leur schéma corporel ne variait pas autant que celui des autres dinosaures, peut-être en raison de contraintes dimensionnelles, mais ils présentaient tout de même une grande variété. Certains, comme les diplodocidés, étaient extrêmement longs et possédaient une queue démesurément longue, qu'ils pouvaient peut-être faire claquer comme un fouet[2]. Le Supersaurus, avec 40 mètres, est probablement le plus long, mais d'autres, tels le Diplodocus (l'ancien détenteur du record), restent extrêmement longs. L'Amphicoelias fragillimus, dont on ne possède plus que l'esquisse d'une seule vertèbre, aurait eu une colonne vertébrale encore plus longue que celle de la baleine bleue. Le plus long des animaux terrestres en vie aujourd'hui, le python réticulé, n'atteint qu'une longueur de 10 mètres.
D'autres, comme les brachiosauridés étaient de haute taille, avec de hautes épaules et un cou extrêmement long. Le Sauroposeidon avait probablement la plus haut stature, atteignant 18 mètres, le précédent record de hauteur étant détenu par le Mamenchisaurus. En comparaison, la girafe, l'animal le plus grand de nos jours, n'atteint que 4,80 m à 5,5 m de haut.
Certains étaient incroyablement massifs : l’Argentinosaurus est probablement le plus lourd avec 80 à 100 tonnes, bien que le Paralititan, l’Andesaurus, l’Antarctosaurus, et l'Argyrosaurus soient de dimensions comparables. Il existe un indice très mince d'un titanosaure encore plus massif, le Bruhathkayosaurus, qui pourrait avoir pesé entre 175 et 220 tonnes ! Le gros plus animal terrestre, l'éléphant d'Afrique, ne pèse pas plus de 10 tonnes.
Parmi les plus petits des sauropodes, se trouvaient l’Ohmdenosaurus (4 m de long), le titanosaure nain Magyarosaurus (5,30 m) et le brachiosauridé nain Europasaurus, qui mesurait 6,20 mètres à l'âge adulte[3]. Sa petite stature résultait vraisemblablement du nanisme insulaire d'une harde de sauropodes isolés, sur une île qui correspond à l’Allemagne actuelle. Remarquable également est le sauropode diplodocoïdé Brachytrachelopan, qui était le plus petit membre du groupe en raison de son cou inhabituellement court. Contrairement aux autres sauropodes, dont les cous pouvaient atteindre jusqu'à quatre fois la longueur de leur tronc, le cou des Brachytrachelopans était plus court que leur colonne vertébrale.
Membres et extrémités
Comme quadrupèdes massifs, les sauropodes développèrent des membres spécialement adaptés pour supporter un poids très élevé. Les pieds arrières étaient larges, et possédaient trois griffes chez la plupart des espèces. Particulièrement inhabituels en comparaison des autres animaux, étaient les pieds avants grandement modifiés (manus). Les pieds avants des sauropodes étaient très différents de ceux des gros quadrupèdes modernes, comme l’éléphant. Au lieu d'être évasés pour créer un pied à large appui comme chez ces derniers, les os de la manus étaient arrangés en colonnes complètement verticales, avec des phalanges extrêmement diminuées (bien que les sauropodes les plus primitifs, comme le Vulcanodon et le Barapasaurus, eussent des pieds avants évasés comportant des doigts). Les pieds avant étaient tellement modifiés chez les eusauropodes que les doigts individuels ne devaient pas être discernables, avec une manus en forme de sabot[4].
La disposition des colonnes (metacarpales) du pied avant chez les eusauropodes était semi-circulaire, de sorte que les empreintes de pas des sauropodes sont en forme de fer à cheval. Contrairement aux éléphants, l'examen des empreintes montre que les sauropodes n'avaient pas de coussinets pour soutenir les pieds avants, ce qui les rendait concaves[5]. La seule griffe visible chez la plupart des sauropodes était la griffe du pouce, nettement distincte (associée avec le doigt 1). Presque tous les sauropodes possédaient une telle griffe, bien que son utilité soit inconnue. La griffe était très grande (mais également longue et latéralement plate) chez les diplodocidés, et très petite chez les brachiosauridés, dont certains paraissent avoir complètement perdue cette griffe, d'après l'examen des traces de pas[6].
Les titanosaures perdirent de même complètement la griffe du pouce (à l'exception de formes primitives telles que les Janenschia). Les titanosaures étaient presque insolites chez les sauropodes, car en plus de la griffe externe, ils perdirent complètement les doigts des pieds avants. Les titanosaures évolués n'avaient ni doigts ni phalanges, et marchaient seulement sur des « moignons » en forme de fer à cheval, composés des os en forme de colonne du métacarpe[7].
Des empreintes au Portugal montrent que, au moins chez quelques sauropodes (probablement des brachiosauridés), le bas et les côtés de la colonne du pied avant étaient vraisemblablement recouverts de petites écailles épineuses, qui ont laissé des entailles sur les empreintes[8]. Chez les titanosaures, les extrémités des métacarpes qui faisait contact avec le sol était inhabituellement larges et carrées. Quelques spécimens conservent des tissus mous couvrant cette surface, ce qui donne à penser que les pieds avants était entourés avec une sorte de coussinet chez ces espèces[7].
Sacs aériens
Comme d'autres dinosaures saurischiens (tels que les oiseaux et autres théropodes), les sauropodes possédaient un système de sacs aériens, dont l'existence est prouvée d'après les indentations et les cavités creuses de la plupart de leur vertèbres. On appelle de telles ouvertures dans le dos, le cou et l'appendice caudal, pneumaticité, et des os creux, pneumatiques sont caractéristiques des sauropodes[9].
Le caractère creux des os des sauropodes, similaire à celui des oiseaux, fut mis en évidence tôt dans l'étude de ces animaux, et en fait, un spécimen de sauropode découvert au XIXe siècle, l’Ornithopsis, fut d'abord confondu avec un ptérosaure volant, précisément à cause de ce fait[10].
Armure
Certains sauropodes avaient une armure. Il existait des genres pourvus de dos couverts d'épines, tels que l’Agustinia, certains d'entre eux possédaient une petite massue, comme le Saltasaurus, et l’Ampelosaurus avait de minuscules ostéodermes recouvrant des portions de son corps.
Paléobiologie
Troupeaux et soin des petits
 Quelques sauropodes, tels que les Alamosaurus sanjuanensis, formaient des troupeaux regroupées selon l'âge.
Quelques sauropodes, tels que les Alamosaurus sanjuanensis, formaient des troupeaux regroupées selon l'âge.De nombreux indices fossilisés, que ce soient les gisements d'ossements ou les empreintes de pas, indiquent que les sauropodes étaient des animaux grégaires qui formaient des troupeaux. Cependant, la composition des troupeaux variait selon les espèces. Quelques gisements d'os, par exemple dans un site du Jurassique moyen en Argentine, semblent montrer des troupeaux composés d'individus de divers âges, mélangeant les jeunes et les adultes. Toutefois, quantité d'autres sites de fossiles et d'empreintes indiquent que de nombreuses espèces de sauropodes se déplaçaient en troupeaux formés selon l'âge, les jeunes formant des troupeaux séparés des adultes. De telles stratégies de regroupement ont été mises en évidence chez l’Alamosaurus, le Bellusaurus, et quelques diplodocidés[11].
Devant l'évidence de divers type de troupeaux, Myers et Fiorillo essayèrent d'expliquer pourquoi les sauropodes paraissent avoir souvent formé des troupeaux séparés par l'âge. Des études de l'usure des dents au microscope montrent que les sauropodes juvéniles avaient un régime alimentaire différent de celui des adultes. Cette différence d'usure dentaire donne à penser que les sauropodes juvéniles avaient une stratégie alimentaire autre que celle des adultes ; par conséquent, se regrouper indistinctement n'aurait pas été aussi productif que le regroupement par âge, où des membres individuels pouvaient fourrager de manière coordonnée. L'énorme différence de taille entre les petits et les adultes a pu également jouer son rôle dans les différentes stratégies alimentaires et de regroupement[11].
Puisque la ségrégation entre petits et adultes devait se mettre en place peu après l'éclosion, Myers et Fiorillo conclurent que les espèces aux troupeaux séparés par l'âge ne devaient guère prendre soin de leurs petits, si ce n'est même pas du tout[11]. D'autre part, les scientifiques qui ont étudié les sauropodes aux troupeaux d'âge indistinct suggèrent que ces espèces devaient prendre soin de leurs petits sur une période étendue avant que les petits atteignissent l'âge adulte[12].
Savoir comment la répartition entre troupeaux séparés par l'âge et troupeaux indistincts variait selon les sauropodes est impossible. D'autres exemples de comportement grégaire devront être découverts à partir d'autres espèces de sauropodes pour parvenir à déterminer un éventuel schéma de répartition[11].
Position cabrée
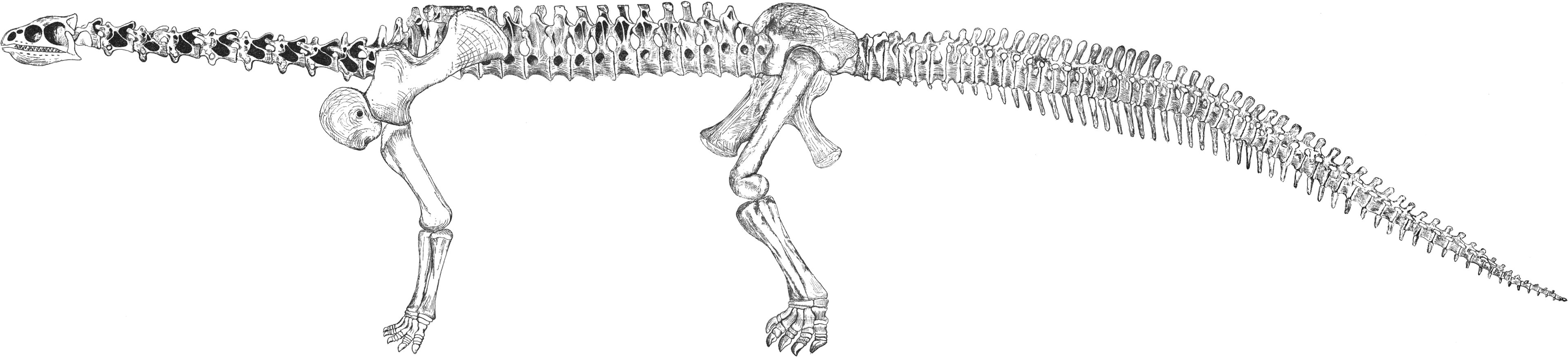
 Squelette monté d'un Barosaurus lentus, illustrant la station redressée tripodale.
Squelette monté d'un Barosaurus lentus, illustrant la station redressée tripodale.Tôt dans l'histoire de leurs études, des scientifiques comme Osborn ont supposé que les sauropodes étaient capables de se redresser sur leurs pattes arrières, utilisant la queue comme la troisième « patte » d'un trépied[13]. Un montage du squelette montrant le diplodocidé Barosaurus lentus se redressant sur ses pattes arrières à l’American Museum of Natural History est une illustration de cette hypothèse. Dans un article de 2005, Carpenter et Tidswell ont objecté que si les sauropodes avaient adopté occasionnellement une posture bipède, il y aurait des traces de fractures de fatigue sur les « mains » des membres supérieurs. Néanmoins, aucune ne fut découverte après qu'ils eurent examiné un grand nombre de squelette de sauropodes[14].
Heinrich Mallison a été le premier, en 2009, à étudier la possibilité physique que divers sauropodes aient pu se cabrer dans une posture tripodale. Mallison a découvert que certaines caractéristiques reliées à l'adaptation au cabrage étaient, en fait, sans relation (comme des os larges de la hanche, chez les titanosaures) ou auraient rendu le cabrage très difficile. Par exemple, les titanosaures avaient une colonne vertébrale particulièrement souple, ce qui aurait réduit la stabilité de la posture tripodale et aurait exercé plus de tension sur les muscles. De même, il est improbable que les brachiosauridés aient pu se redresser sur leurs membres postérieurs, étant donné que leur centre de gravité se situait beaucoup plus en avant que chez les autres sauropodes, ce qui aurait rendu une telle posture instable[15].
À l'inverse, les diplodocidés semblent avoir été bien adaptés pour se cabrer en posture tripodale. Ceux-ci possédaient un centre de gravité directement au-dessus des hanches, ce qui leur procuraient un plus grand équilibre sur deux pattes. Les diplodocidés avaient également le cou le plus mobile des sauropodes, une ceinture pelvienne bien musclée, et des vertèbres caudales à forme spécialisée qui permettait à la queue de supporter leur poids au point de contact avec le sol. Mallison en conclut que les diplodocidés étaient mieux adaptés pour se cabrer que les éléphants, qui le font occasionnellement dans la nature. Les diplodocidés semblent, toutefois, être les seuls sauropodes qui fussent aptes à cette posture[15].
Position du cou et de la tête
Il y a controverse pour savoir si les sauropodes portaient leur tête verticalement ou horizontalement. L'affirmation selon laquelle leur long cou permettait aux sauropodes de brouter dans les hauts arbres a été remise en cause sur la base d'un calcul de l'énergie nécessaire pour créer la pression artérielle nécessaire à la tête, si elle était maintenue verticalement. Ces calculs suggèrent que cela aurait exigé la moitié de sa ration alimentaire[16]. Qui plus est, pour irriguer la tête tenue verticalement, une pression sanguine de 700 mmHg aurait été nécessaire au niveau du cœur. Cela aurait requis que leur cœur fût 15 fois plus gros que celui des baleines de même taille[17]. Ce qui donne à penser qu'il est plus probable que le long cou était tenu horizontalement pour leur permettre de se nourrir de plantes sur une large étendue sans besoin de mouvoir leur corps — une économie d'énergie importante pour des animaux de trente à quarante tonnes. À l'appui de cette théorie, des reconstructions du cou du Diplodocus et de l’Apatosaurus montrent qu'il est droit à la base avec une légère inclinaison qui oriente leur tête dans une « position neutre et non-fléchie » quand elle est proche du sol[18].
Traces de pas et locomotion
 A narrow-gauge sauropod trackway.
A narrow-gauge sauropod trackway.On sait que les sauropodes ont laissé de très nombreuses traces de pas avec des empreintes de pieds (qu'on appelle ichnites[19]) sur la plupart des continents. Les ichnites ont permis d'élaborer d'autres hypothèses biologiques concernant les sauropodes, y compris les pieds avant et arrières (cf. supra « Membres et extrémités »). Les empreintes des pieds de devant sont beaucoup plus petites que celles des pieds de derrière, et sont souvent en forme de croissant. Parfois, les ichnites conservent des traces de griffes et permettent de déterminer quels groupes de sauropodes ont perdu des griffes ou même des orteils de leurs pieds avants[20].
Les traces de pas des sauropodes se divisent généralement en trois catégories fondées sur la distance entre membres opposés : petit écartement, écartement moyen et grand écartement. L'écartement des traces de pas permet de déterminer l'écart entre membres de divers sauropodes, et comment cet écart influençait leur manière de marcher[20]. Une étude menée en 2004 par Day et ses collègues a établit qu'un schéma général se dégageait parmi les groupes de sauropodes évolués, dont chaque famille de sauropodes se caractérisait par un certain écartement visible sur les traces de pas. Ils ont découvert que la plupart des sauropodes autres que les titanosaures présentaient un écartement étroit, avec des empreintes profondes de la griffe du gros orteil des pieds de devant. Les traces à écartement moyen avec empreinte du pouce provenaient probablement de brachiosauridés et d'autres titanosauriformes primitifs, qui développaient des membres à écartement plus large mais retenaient leurs griffes. Les membres à écartement large étaient detenus par des titanosaures évolués, dont les traces de pas montrent un écartement large et l'abscence de griffes ou d'orteils sur les pieds avants[21].
Évolution de la taille
Plusieurs scientifiques ont essayé d'aborder la question pourquoi les sauropodes atteignaient des tailles aussi énormes. Des tailles gigantesques furent atteintes tôt dans l'évolution des sauropodes, qui remonte aux premiers véritables sauropodes, au triassique supérieur. Selon Kenneth Carpenter, quelle que soit la pression évolutive qui ait entraîné une grosse taille, elle doit avoir été présente depuis les origines du groupe[22].
Des études sur les mammifères herbivores qui atteignent une grande taille, tels que les éléphants, ont établit qu'une taille plus grande chez les herbivores entraine une plus grande efficacité pour digérer la nourriture. Comme les animaux plus gros ont des systèmes digestifs plus longs, la nourriture est gardée sur des périodes nettement plus longues, ce qui permet aux gros animaux de survivre de sources de nourriture de moins bonne qualité. Cela est particulièrement vrai des animaux qui ont un grand nombre de chambres de fermentation le long des intestins, ce qui permet la prolifération microbienne et la fermentation de la matière végétale, favorisant ainsi la digestion. À travers leur évolution, les dinosaures sauropodes vivaient principalement en milieux semi-arides avec une saison sèche et une baisse correspondante de la qualité de la nourriture durant cette saison sèche. L'environnement de la plupart des sauropodes géants du Jurassique supérieur, tels que l’Amphicoelias, était essentiellement la savane, semblable aux milieux arides où vivent les herbivores géants modernes, ce qui appuie l'idée qu'une nourriture de faible qualité en milieu aride favorise l'évolution des herbivores géants. Carpenter a soutenu que les autres bénéfices d'une grande taille, tels qu'une relative immunité vis à vis des prédateurs, une plus faible dépense d'énergie et une plus grande longévité, était probablement des avantages secondaires et que les sauropodes atteignaient une grande taille principalement pour une digestion plus efficace[22].
Histoire de la découverte
Les premiers restes fossiles fragmentaires, reconnus maintenant comme sauropodes, venaient tous d’Angleterre, et furent interprétés, au départ, en diverses manières. Leur lien avec les autres dinosaures ne fut reconnu que bien plus tard après leur découverte initiale.
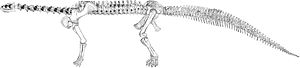
 Squelette reconstitué moderne d'un Camarasaurus.
Squelette reconstitué moderne d'un Camarasaurus.Le premier fossile de sauropode décrit scientifiquement était une unique dent, nommée par son descripteur (non-linnéen) Rutellum implicatum[23]. Ce fossile fut décrit par Edward Lhuyd en 1699, mais ne fut pas reconnu comme celui d'un reptile préhistorique géant, sur le moment[24]. Les dinosaures ne seraient reconnus comme groupe que plus d'un siècle plus tard.
Richard Owen publia la première description scientifique moderne des sauropodes, en son rapport nommant le Cetiosaurus et le Cardiodon. Le Cardiodon n'était connu qu'à partir de deux dents inhabituelles en forme de cœur (desquelles il tira son nom), qui ne pouvaient être identifiées au-delà du fait qu'elle provenaient d'un reptile inconnu jusqu'alors. Le Cetiosaurus était un peu mieux connu, mais avec des restes encore fragmentaires. Owen pensa, à l'époque, que le Cetiosaurus était un reptile marin géant apparenté aux crocodiles modernes, d'où son nom, qui signifie « lézard-baleine ». Un an plus tard, quand Owen forgea le mot Dinosauria, il n'inclut pas Cetiosaurus et Cardiodon dans le groupe[25].
En 1850, Gideon Mantell reconnut la nature « dinosaurienne » de plusieurs os attribués au Cetiosaurus par Owen. Mantell remarqua que les os cruraux contenaient une cavité médullaire propre aux animaux terrestres. Il classa les échantillons dans un nouveau genre, Pelorosaurus, qu'il rattacha avec les dinosaures. Cependant, Mantell ne reconnaissait toujours pas le lien de parenté avec le Cetiosaurus[10].
La prochaine découverte de sauropode à être décrite et erronément rattachée à quelque chose d'autre qu'un dinosaure, est un ensemble de vertèbres de la hanche, décrites par Harry Seeley en 1870. Seeley trouva que les vertèbres étaient de construction très légère pour leur taille, et qu'elles contenaient des ouvertures pour des sacs aériens (pneumatisation). Des sacs aériens semblables n'étaient connu, à l'époque, que sur les oiseaux et les ptérosaures, et Seeley considéra les vertèbres comme appartenant à un ptérosaure. Il nomma le nouveau genre Ornithopsis, ou « face d'oiseau », pour cette raison[10].
Quand Phillips décrit des échantillons plus précis du Cetiosaurus en 1871, il reconnut finalement l'animal comme un dinosaure apparenté au Pelorosaurus[26]. Néanmoins, ce n'est pas avant la description de nouveaux squelettes presque complets trouvés aux États-Unis (representant l’Apatosaurus et le Camarasaurus), plus tard en cette même année, qu'une nouvelle image des sauropodes émergea. Une reconstitution approximative d'un squelette complet de sauropode fut effectuée par John A. Ryder, fondée d'après les restes d'un Camarasaurus, bien que de nombreuses caractéristiques fussent encore inexactes ou incomplètes, selon les découvertes et études biomécaniques ultérieures[27]. De même, en 1877, Richard Lydekker nomma un autre parent du Cetiosaurus, le Titanosaurus, d'après une vertèbre isolée[10].
En 1878, le sauropode le plus complet fut découvert et décrit par Othniel Charles Marsh, qui le baptisa Diplodocus. Avec cette découverte, Marsh créa aussi un nouveau groupe pour accueillir les Diplodocus, les Cetiosaurus, afin de les différencier des autres grands groupes de dinosaures, car leurs listes de parents était en augmentation. Marsh nomma ce groupe Sauropoda, ou « pieds de lézard »[10].
Classification
La Classification des sauropodes s'est nettement stabilisée ces dernières années, bien qu'il y ait toujours des incertitudes, comme la position de l’Euhelopus, de l’Haplocanthosaurus, du Jobaria et des Nemegtosauridae. Ce qui suit sont deux récentes classifications alternatives (indiquant seulement les clades supra-génériques, dans le second exemple). Il ne s'y trouve en aucune manière de liste exhaustive des schémas de classification des sauropodes récents. En certains cas, des familles telles que les Vulcanodontidae, les Cetiosauridae et les Omeisauridae n'y sont pas incluses parce qu'elles sont considérées comme paraphyletiques, ou même (dans le cas des Camarasauridae), comme polyphyletiques.
Taxonomie
Cette taxonomie ci-après suit Wilson & Sereno 1998, Yates 2003, Galton 2001 [2] et Wilson 2002[28], avec rangs d'après Benton, 2004[29].
- Infra-ordre Sauropoda
- Ammosaurus
- Anchisaurus
- ?Isanosaurus
- Kotasaurus
- Lessemsaurus
- Famille Blikanasauridae
- Famille Melanorosauridae
- Famille Vulcanodontidae
- Famille Cetiosauridae
- Famille Omeisauridae
- ?Famille Tendaguridae
- Clade Turiasauria
- Division Neosauropoda
- Haplocanthosaurus
- ?Jobaria
- Super-famille Diplodocoidea
- Famille Rebbachisauridae
- Famille Dicraeosauridae
- Famille Diplodocidae
- Subdivision Macronaria
- Famille Brachiosauridae
- Famille Camarasauridae
- Famille Euhelopodidae
- Super-famille Titanosauroidea
Phylogénie
Cladogramme simplifié d'après Wilson, 2002[28].
Sauropoda Eusauropoda Shunosaurus
unnamed unnamed unnamed Omeisaurus
unnamed Neosauropoda Haplocanthosaurus
Macronaria Titanosauriformes Somphospondyli Euhelopus
Titanosauria
Références
- ↑ Marsh, O.C. (1878). "Principal characters of American Jurassic dinosaurs. Part I". American Journal of Science and Arts 16: 411-416
- ↑ Whips and Dinosaur Tails
- ↑ Sander, P.M., Mateus, O., Laven, T. & Knötschke, N. (2006). "Bone histology indicates insular dwarfism in a new Late Jurassic sauropod dinosaur." Nature, 441: 739-741.
- ↑ Paul, G.S. (1987). "The science and art of restoring the life appearance of dinosaurs and their relatives - a rigorous how-to guide." Pp. 4-49 in Czerkas, S.J. and Olson, E.C. (eds.), Dinosaurs Past and Present Vol. II. Seattle: University of Washington Press.
- ↑ Bonnan, M.F. (2003). "The evolution of manus shape in sauropod dinosaurs: implications for functional morphology, forelimb orientation, and phylogeny." Journal of Vertebrate Paleontology, 23: 595-613.
- ↑ Upchurch, P. (1994). "Manus claw function in sauropod dinosaurs." Gaia, 10: 161-171.
- ↑ a et b Apesteguía, S. (2005). "Evolution of the titanosaur metacarpus." Pp. 321-345 in Tidwell, V. and Carpenter, K. (eds.) Thunder-Lizards: The Sauropodomorph Dinosaurs. Indianapolis: Indiana University Press.
- ↑ Milàn, J., Christiansen, P. and Mateus, O. (2005). "A three-dimensionally preserved sauropod manus impression from the Upper Jurassic of Portugal: implications for sauropod manus shape and locomotor mechanics." Kaupia, 14: 47-52.
- ↑ Wedel, M.J. (2009). "Evidence for bird-like air sacs in Saurischian dinosaurs." Journal of Experimental Zoology, 311A: 18pp.
- ↑ a , b , c , d et e Taylor, M.P. (In press). "Sauropod dinosaur research: a historical review." In Richard Moody, Eric Buffetaut, David M. Martill and Darren Naish (eds.), Dinosaurs (and other extinct saurians): a historical perspective. HTML abstract.
- ↑ a , b , c et d Myers, T.S. and Fiorillo, A.R. (2009). "Evidence for gregarious behavior and age segregation in sauropod dinosaurs." Palaeogeography, Palaeoclimatology, Palaeoecology, 274: 96-104.
- ↑ Coria, R.A. (1994). "On a monospecific assemblage of sauropod dinosaurs from Patagonia: implications for gregarious behavior." GAIA, 10: 209–213.
- ↑ Osborn, H.F. (1899). "A skeleton of Diplodocus, recently mounted in the American Museum". Science, new series, vol. 10: 870-874.
- ↑ (en) Rothschild, B.M. and Molnar, R.E., Thunder Lizards: The Sauropodomorph Dinosaurs, Indiana University Press, 2005 (ISBN 0-253-34542-1), « Sauropod Stress Fractures as Clues to Activity », p. 381–391
- ↑ a et b Mallison, H. (2009). "Rearing for food? Kinetic/dynamic modeling of bipedal/tripodal poses in sauropod dinosaurs." P. 63 in Godefroit, P. and Lambert, O. (eds), Tribute to Charles Darwin and Bernissart Iguanodons: New Perspectives on Vertebrate Evolution and Early Cretaceous Ecosystems. Brussels.
- ↑ Seymour, R.S. (2009). "Raising the sauropod neck: it costs more to get less". Biol Lett. 23;5(3):317-9. PMID 19364714
- ↑ Seymour, R.S. and Lillywhite, H.B. (2000). "Hearts, neck posture and metabolic intensity of sauropod dinosaurs". Proc Biol Sci. 22;267(1455):1883-7. PMID 11052540
- ↑ Stevens, K.A. and Parrish, J.M. (1999). "Neck posture and feeding habits of two Jurassic sauropod dinosaurs". Science. 284(5415):798-800. PMID 10221910
- ↑ Mot féminin
- ↑ a et b Riga, B.J.G. and Calvo, J.O. (2009). "A new wide-gauge sauropod track site from the Late Cretaceous of Mendoza, Neuquen Basin, Argentina." Paleontology, 52(3): 631-640.
- ↑ Day, J.J., Norman, D.B., Gale, A.S., Upchurch, P. and Powell, H.P. (2004). "A Middle Jurassic dinosaur trackway site from Oxfordshire, UK." Palaeontology, 47: 319–348.
- ↑ a et b Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus." In Foster, J.R. and Lucas, S.G., eds., 2006, Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin 36: 131-138.[1]
- ↑ Delair, J.B., and Sarjeant, W.A.S. (2002). "The earliest discoveries of dinosaurs: the records re-examined." Proceedings of the Geologists' Association, 113: 185-197.
- ↑ Lhuyd, E. (1699). Lithophylacii Britannici Ichnographia, sive lapidium aliorumque fossilium Britannicorum singulari figura insignium. Gleditsch and Weidmann: London.
- ↑ Owen, R. (1842). "Report on British Fossil Reptiles." Part II. Report of the British Association for the Advancement of Science, Plymouth, England.
- ↑ Phillips, J. (1871). Geology of Oxford and the Valley of the Thames. Oxford: Clarendon Press, 523 pp.
- ↑ Osborn, H.F., and Mook, C.C. (1921). "Camarasaurus, Amphicoelias and other sauropods of Cope." Memoirs of the American Museum of Natural History, n.s. 3:247-387 and plates LX-LXXXV.
- ↑ a et b Wilson, J. A. (2002). "Sauropod dinosaur phylogeny: critique and cladistic analysis." Zoological Journal of the Linnean Society, 136: 217-276.
- ↑ Benton, M.J. (2004). Vertebrate Palaeontology, Third Edition. Blackwell Publishing, 472 pp.
- Bob Strauss, 2008, Sauropods: The Biggest Dinosaurs that Ever Lived, The New York Times
- Kristina Curry Rogers and Jeffrey A. Wilson, 2005, The Sauropods: Evolution and Paleobiology, University of California Press, Berkeley, ISBN 0-520-24623-3
- Upchurch, P., Barrett, P.M. and Dodson, P. 2004. Sauropoda. In The Dinosauria, 2nd édition. D. Weishampel, P. Dodson, and H. Osmólska (eds.). University of California Press, Berkeley. Pp. 259–322.
Liens externes
- Référence Tree of Life Web Project : Sauropoda (en)
- Référence The Paleobiology database : Sauropoda Marsh 1878 (en)
Sources
- (en) Cet article est partiellement ou en totalité issu d’une traduction de l’article de Wikipédia en anglais intitulé « Sauropoda ».
 Portail de la paléontologie
Portail de la paléontologie Portail des dinosaures
Portail des dinosaures Portail de la zoologie
Portail de la zoologie
Catégorie : Sauropoda





Wikimedia Foundation. 2010.