- Macaca thibetana
-
Macaque du Tibet
Macaque du Tibet

Macaca thibetana. Classification classique Règne Animalia Embranchement Chordata Sous-embr. Vertebrata Classe Mammalia Sous-classe Theria Infra-classe Eutheria Ordre Primates Famille Cercopithecidae Sous-famille Cercopithecinae Genre Macaca Nom binominal Macaca thibetana
Milne-Edwards, 1870Répartition géographique 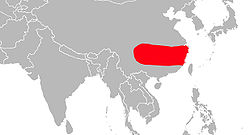
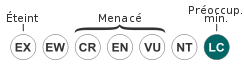
Statut de conservation IUCN :
LC : Préoccupation mineure
 Retrouvez ce taxon sur Wikispecies
Retrouvez ce taxon sur Wikispecies
D'autres documents multimédia
sont disponibles sur CommonsParcourez la biologie sur Wikipédia : 





Sommaire
Caractéristiques générales
 Macaque du Tibet Macaca thibetana. Photo Noel Rowe.
Macaque du Tibet Macaca thibetana. Photo Noel Rowe.
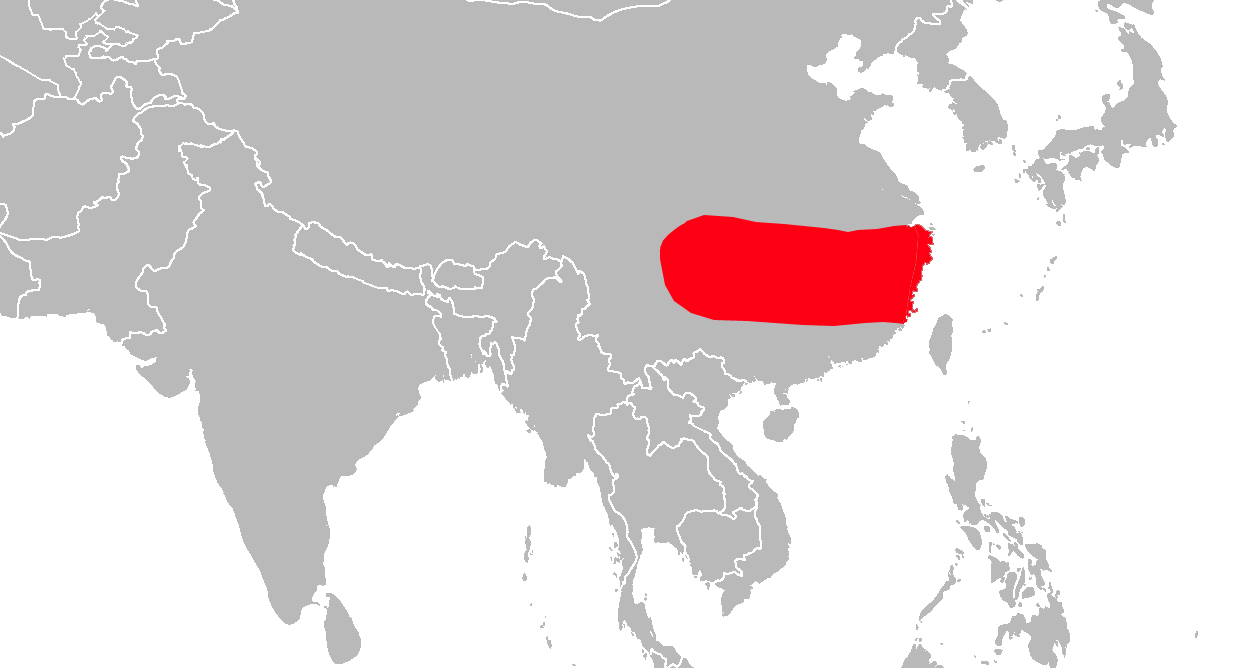
Le macaque du Tibet vit dans le sud-est de la Chine, de l'Himalaya à la mer de Chine. C'est un macaque très massif dont la queue est très réduite et qui vit surtout au sol. Il appartient au groupe sinica-arctoides des macaques dont il est le représentant le plus septentrional. Son pelage est brun foncé à noir sur la face dorsale et beige presque blanc sur la face ventrale. Une mue du pelage a lieu à la fin de l'été à l'approche de la clôture de la saison des pluies. Une petite éminence de pelage plus long est perceptible sur le haut du crâne et le faciès est cerné d'une barbe et de favoris importants. La peau faciale est relativement claire chez les jeunes mais peut s'orner de taches rougeâtres et noires ou considérablement foncer chez certains individus adultes.
Reproduction
La reproduction est saisonnière, avec une période d'accouplement faisant un pic de fin septembre à début novembre, couplée à une période de naissance en mars-avril. Les femelles en œstrus présentent un fort gonflement de la zonne ano-génitale et initient les copulations par des approches et des présentations devant les mâles.
Le comportement sexuel du macaque du Tibet est, par ailleurs, assez complexe. Vingt catégories de comportements sexuels ont été rapportés démontrant la grande complexité du répertoire des interactions socio-sexuelles de cette espèce comparativement à d'autres macaques. La diversité du mode de regroupement est importante durant la saison de reproduction. Les montes sont plus fréquentes au centre du groupe qu'à la périphérie, bien que la compétition y soit plus importante. Les jeunes mâles adultes immigrants sont les plus actifs sexuellement. Les individus subordonnés ( mâles et femelles ) semblent avoir une stratégie d'appariement moins risquée à la périphérie du groupe.
 Femelle adulte avec un juvénile. Photo Noel Rowe.
Femelle adulte avec un juvénile. Photo Noel Rowe.Les jeunes mâles adultes, classe d'âge la plus active sexuellement et dans les migrations inter-groupes, reçoivent le plus de blessures. Les mâles adultes plus âgés sont les plus agressifs et infligent de nombreuses blessures aux mâles sub-adultes et jeunes adultes. Pendant la saison de reproduction, une très forte proportion de jeunes mâles adultes et quelques mâles plus âgés changent de groupe. Bien que les jeunes mâles adultes encourent plus de risques de blessures, ils acquièrent généralement un rang hiérarchique plus élevé suite à ce transfert. Les mâles adultes plus âgés émigrent moins fréquemment mais chutent dans la hiérarchie de dominance. Les mâles sub-adultes périphériques, qui émigrent rarement, jouent un rôle actif et déterminant dans le taux de blessures ainsi que dans le taux et la direction des migrations au sein du groupe. Certaines blessures infligées par les mâles résidents conduisent à la mort des mâles immigrants.
Les copulations non-reproductives, qui ont lieu en dehors de la saison de reproduction, sont moins fréquentes, moins longues et ne sont pas entrecoupées de pauses avec des vocalisations comme c'est le cas pour les copulations ayant lieu lors de la saison de reproduction. Ces montes ont souvent lieu lorsqu'une femelle qui n'allaite pas est impliquée dans un conflit ou qu'elle approche le mâle pour susciter l'accouplement. Aucune femelle gestante ou allaitante n'est généralement concernée par ces montes non-reproductives. Les paires d'individus ayant effectué ces montes non-reproductives passent plus de temps ensemble lors de la recherche alimentaire, ce qui reflète une plus grande tolérance sociale de la part du mâle. Les mâles juvéniles, qui copulent rarement à la saison de reproduction, sont impliqués dans ces montes, et bien qu'elles n'aient pas de fonctions reproductives, elles semblent remplir une fonction sociale pour les femelles, comme le soutien ou l'apaisement des mâles lors de conflits ou un meilleur accès aux ressources alimentaires. Ceci est à mettre en parallèle avec les interventions pacifiques dans les conflits que l'on rencontre fréquemment chez les macaques de Sulawesi, réputés pour la grande tolérance sociale de leur organisation.
Organisation sociale
Les groupes de macaques du Tibet comprennent en moyenne une vingtaine de membres ( 10 à 70 individus ). Des mâles peuvent demeurer solitaires. Les groupes comprennent à la fois des mâles et des femelles mâtures accompagnées de leur jeunes comme chez les autres espèces de macaques. Dans la composition moyenne d'une troupe les adultes ( femelles de plus de 5 ans et mâles de plus de 7 ans )représentent 40 % des individus, les sub-adultes ( femelles de 3 à 5 ans et mâles de 3 à 7 ans ) 25 %, les juvéniles ( de 1 à 3 ans ) 20 % et les jeunes de l'année 15 %. Dans un groupe étudié à Yulingkeng, la mortalité infantile n'est pas très élévée pendant la première année de vie, mais seuls 50 % des individus atteignent l'âge de 4 ans. Cette valeur est cependant supérieure à ce que l'on observe en milieu naturel chez le macaque japonais ou le macaque à toque.
 Un groupe de macaque du Tibet en milieu naturel. Photo Noel Rowe.
Un groupe de macaque du Tibet en milieu naturel. Photo Noel Rowe.Chez le macaque du Tibet il existe une hiérarchie de dominance linéaire entre les différents mâles adultes du groupe, mais celle-ci est peu stable au cours du temps et change généralement chaque année avant la saison d'accouplement ou durant la première moitié de cette période. Les mâles adultes les plus jeunes occupent la position la plus dominante et réciproquement de façon graduelle en fonction de l'âge. Les jeunes mâles adultes ou les mâles subadultes émigrent généralement à l'approche de la saison des accouplements. Cette migration n'est pas dépendante du rang hiérarchique dans le groupe de naissance. Les mâles ayant récemment migré dans un nouveau groupe ont plus d'interactions affiliatives et sexuelles avec les femelles adultes que les membres originaires du groupe. Les macaques du Tibet se blottissent en sous-groupes pendant la nuit sur leur site de sommeil, généralement, au bord d'un rocher abrupt. Les femelles forment moins fréquemment des triades avec deux mâles pendant la saison des accouplements, qu'elles ne le font à la saison des naissances. Le choix des partenaires de sommeil semble cependant fortement lié au type de relations qu'entretiennent les différents protagonistes sur de longues périodes plus qu'à des changements ponctuels comme ceux qui interviennent lors des accouplements et des naissances.
Comme beaucoup des primates étudiés à ce jour, les macaques du Tibet se reconcilient après un conflit, c'est-à-dire que dans l'ensemble, les opposants s'engagent dans une interaction affiliative immédiatement après un conflit à une fréquence plus importante qu'à n'importe quel autre moment. Les probabilités d'interaction pacifique peuvent augmenter jusqu'à 30 fois après la fin des hostilités. Cependant, la tendance conciliatoire est faible ( 6,4 % ) chez cette espèce, comparée à d'autres qualifiées de plus tolérantes. De plus, les femelles privilégient fortement les membres du groupe qui leur sont apparentés dans les intéractions affiliatives et tolérantes. Ces deux traits sont typiques des systèmes sociaux de type despotique qu'on peut rencontrer chez le macaque rhésus ou le macaque japonais, par exemple. C'est le seul macaque du groupe sinica-arctoides à posséder un système social aussi despotique.La fission de groupes a été rapportée plusieurs fois dans des groupes comprenant de nombreux individus et possédant le plus haut rang parmi les groupes environnants. La fission a lieu quand le nombre de membres a atteint une certaine limite, préférentiellement durant la saison de reproduction à la période des accouplements, là où le niveau de compétition est à son paroxysme. La fission peut être initiée par un mâle et une femelle adultes qui entretiennent une longue relation. La ligne de fission suit globalement les lignées maternelles mais un facteur social, comme le fait d'avoir de bonnes relations avec un individu, peut aussi jouer un rôle important dans le choix du groupe d'appartenance. Des mâles adultes ayant occupé autrefois un haut rang hiérarchique peuvent être à l'origine des fissions. Le groupe initial, malgré la fission d'une partie de ses membres, conserve une taille et une position dominante vis-à-vis des groupes voisins susceptibles de rentrer en compétition avec celui-ci. Ceci suggère que, dans ces deux cas, la fission semble écarter l'excédant du groupe afin de réduire la compétition pour la nourriture et les partenaires sexuels, sans amoindrir le bénéfice de la coopération pour la défense des ressource face aux groupes rivaux.
 Une séance de toilettage d'un juvénile sur un mâle adulte. Photo Noel Rowe.
Une séance de toilettage d'un juvénile sur un mâle adulte. Photo Noel Rowe.Les groupes ayant fissionné dans un récent passé partage le même domaine vital et présente le plus fort taux d'hostilité de la part des femelles lors des rencontres inter-groupes. Par opposition à la forte implication des femelles dans la compétition inter-groupes pour les ressources alimentaires, les mâles adultes sont moins impliqués dans ces conflits. Les femelles forment une sorte de harem autour des mâles de haut rang dans plus de 80 % des rencontres étroites observées entre plusieurs groupes. La formation de harems est une structure sociale rare chez les macaques mais bien documentée chez les babouins, notamment le babouin hamadryas, Papio hamadryas qui vit en Éthiopie. La sélection sexuelle semble jouer un rôle important dans la formation de telles cellules sociales.
Les interactions inter-groupes peuvent prendre la forme d'avertissements de la part des mâles de haut rang ( secouement de branches et/ou cris puissants ), d'interactions à longue distance ( caractérisées par des changements de leurs trajets habituels de recherche alimentaire ) ou de contacts étroits ( avec comportement affiliatif, femelles se regroupant autour d'un mâle, interaction sexuelle, conflit de groupe, conflit dyadique entre deux mâles adultes, avancée ou retraite opportuniste, etc. de la part de différentes classes d'âge et de sexe ). Le schéma des interactions entre groupes semble suivre un profil saisonnier, plus coopératif à la saison des naissances ( pouvant parfois conduire à la fusion temporaire de plusieurs groupes en un même grand groupe ) et plus compétitif à l'approche de la saison des accouplements.
Mensurations
Poids
-
- Mâle : 14 à 17,5 kg
- Femelle : 9,5 à 11 kg
Taille
Longueur tête + corps
-
- Mâle : 580 à 660 mm
- Femelle : 490 à 555 mm
Longueur de la queue
-
- Mâle : 55 à 80 mm
- Femelle : 55 à 80 mm
Habitat
 Macaques du Tibet en milieu naturel. Photo Bernard Thierry.
Macaques du Tibet en milieu naturel. Photo Bernard Thierry.Le macaque du Tibet est essentiellement terrestre et dépose des marques au sol sur ses trajets habituels de déplacement et de recherche alimentaire. Dans son aire de distribution ( 25 à 33 ° N et 1000 à 2500 m d'altitude ), le climat varie du sub-tropical humide au modérément continental humide. L'habitat de prédilection de ces macaques est la forêt pluviale persistente de moyenne montagne. Cette espèce peut toutefois se déplacer vers les zones adjacentes de clairières naturelles, de forêts persistentes de bambou et les champs cultivés. Dans certaines aires, une migration saisonnière peut avoir lieu, entre les hautes altitudes en été ( 1300 à 1700 m ) et les basses altitudes en hiver ( 400 à 600 m ). En pareil cas, les animaux exploitent les versant nords l'été et sud en hiver, vraisemblablement pour s'adpater aux variations extrêmes de température aussi bien qu'à la disponibilité des ressources végétales. Dans d'autres zones, le macaque du Tibet demeure à haute altitude en automne et en hiver.
Les macaques du Tibet sont parmi les rares macaques à ne pas dormir dans des arbres mais plutôt sur des rochers ou dans des grottes.
Nutrition
 Femelle avec un nouveau-né en recherche alimentaire. Photo Noel Rowe.
Femelle avec un nouveau-né en recherche alimentaire. Photo Noel Rowe.Le domaine vital d'un groupe de macaques du Tibet peut être inférieur à 500 ha ( 2,5 km de diamètre ) lorsque la nourriture est abondante ( notamment en période estivale ) et, au contraire, quand la nourriture est disséminée en hiver, les animaux peuvent couvrir jusqu'à 30 km par jour, recherchant des graines en chemin à un rythme de 6 à 7 km/h. Le domaine vital demeure relativement inchangé d'une année sur l'autre.
Le macaque du Tibet est principalement végétarien, la consommation d'animaux avoisinant seulement les 5 % du régime alimentaire. Il peut consommer, en une même localité, au moins 19 espèces différentes de plantes. Les pousses de bambou constituent une proportion importante de sa nourriture. La consommation de feuilles est plus importante chez ce macaque que chez les espèces proches. Sur 17 plantes bien déterminées qu'ingère le macaque du Tibet, 11 le sont pour les feuilles contre 5 seulement pour les fruits. Lorsqu'il s'attaque aux cultures, il consomme souvent du maïs. La nourriture animale peut prendre la forme de consommation d'invertébrés, essentiellement des insectes, ou de petits vertébrés comme les lézards.
La pénurie alimentaire durant l'hiver est une cause fréquente de mortalité et les rivalités intra-groupe pour la nourriture sont communes chez cette espèce.
Population
L'état des populations de macaques du Tibet n'est pas rigoureusement documenté mais l'IUCN ne place pas cette espèce parmi les plus menacées. On ne connait pas de prédateur clairement avéré chez ce macaque bien que des comportements d'attaque aient été observés vis-à-vis du milan noir, Milvus migrans.
Voir aussi
Articles connexes
Bibliographie
- Li J, Yin H & Zhou L, 2007. Non-reproductive copulation behavior among Tibetan macaques (Macaca thibetana) at Huangshan, China. Primates 48 : 64-72.
- Berman CM, Ionica CS, Dorner M & Li J. 2006. Postconflict Affiliation Between Former Opponents in Macaca thibetana on Mt. Huangshan, China. International Journal of Primatology 27 : 827-854.
- Berman CM, Ionica CS & Li J, 2004. Dominance Style Among Macaca thibetana on Mt. Huangshan, China. International Journal of Primatology 25 : 1283-1312.
- Ogawa H & Takahashi H, 2003. Triadic Positions of Tibetan Macaques Huddling at a Sleeping Site. International Journal of Primatology 24 : 591-606.
- Berman CM & Li J-H, 2002. Impact of Translocation, Provisioning and Range Restriction on a Group of Macaca thibetana. International Journal of Primatology 23 : 383-397.
- Zhao Q-K, 1999. Responses to Seasonal Changes in Nutrient Quality and Patchiness of Food in a Multigroup Community of Tibetan Macaques at Mt. Emei. International Journal of Primatology 20 : 511-524.
- Zhao Q-K, 1997. Intergroup interactions in Tibetan macaques at Mt. Emei, China. American Journal of Physical Anthropology 104 : 459-470.
- Li J, Wang Q & Han D, 1996. Fission in a free-ranging Tibetan macaque troop at Huangshan Mountain, China. Chinese Science Bulletin 41 : 1377-1381.
- Li J, Wang Q & Li M, 1996. Migration of male Tibetan monkeys (Macaca thibetana) at Mt. Huangshan, Anhui Province, China. Acta Theriologica Sinica 16 : 1-6.
- Li J & Wang Q-S, 1996. Dominance hierarchy and its chronic changes in adult male Tibetan macaques (Macaca thibetana). Acta Zoologica Sinica 42 : 330-333.
- Rowe N, 1996. The Pictorial Guide to the Living Primates, Pogonias Press, East Hampton, New-York, p 134.
- Li J, Wang Q & Li M, 1995. Studies on the population ecology of Tibetan monkeys (Macaca thibetana) III. Age structure and life table. Acta Theriologica Sinica 15 : 31-35.
- Zhao Q-K, 1994. Mating competition and intergroup transfer of males in Tibetan macaques (Macaca thibetana) at Mt. Emei, China. Primates 35 : 57-68.
- Zhao Q-K, 1993. Sexual behavior of tibetan macaques at Mt. Emei, China. Primates 34 : 431-444.
- Fooden J, 1988. Taxonomy and Evolution of the Sinica Group of Macaques : 6. Interspecific Comparisons and Synthesis. Fieldiana : Zoology 45.
- Fooden J, 1986. Taxonomy and Evolution of the Sinica Group of Macaques : 5. Overview of Natural History. Fieldiana : Zoology 29.
- Fooden J, 1983. Taxonomy and Evolution of the Sinica Group of Macaques : 4. Species Account of Macaca Thibetana. Fieldiana : Zoology 17.
 Portail de la zoologie
Portail de la zoologie
Catégories : Statut IUCN Préoccupation mineure | Primate | Cercopithecidae | Faune endémique de Chine -








Wikimedia Foundation. 2010.