- GroEL
-
La protéine GroEL appartient à la famille des chaperonines des molécules chaperonnes, et se trouve chez un grand nombre de bactéries[1]. Elle est nécessaire pour le repliement efficace de nombreuses protéines. Afin de fonctionner efficacement, GroEL requiert le complexe protéique couvercle associé GroES. Chez les eukaryotes, les protéines Hsp60 et Hsp10 sont structurellement et fonctionnellement presque identiques à GroEL et GroES, respectivement.
Sommaire
Mécanisme d'action
Dans une cellule, le processus de repliement de protéine par médiation GroEL/ES implique de nombreuses séquences de liaisons, encapsulations et libération de protéine substrat. Les protéines substrats non repliées se lient à une aire hydrophobe sur le bord intérieur de la cavité ouverte du GroEL, formant ainsi un complexe binaire avec la chaperonine. La liaison de protéine substrat de cette façon, en plus de la liaison d'ATP, conduit à une modification conformationnelle qui permet l'association du complexe binaire avec une structure couvercle distincte, GroES. La liaison de GroES avec la cavité ouverte de la chaperonine provoque la rotation des sous-unités individuelles de la chaperonine de manière à ce que le substrat hydrophobe liant soit soustrait à l'intérieur de la cavité, ce qui induit l'éjection de la protéine substrat du bord vers la chambre maintenant hydrophile. L'environnement hydrophile de la chambre favorise l'enfouissement des résidus hydrophobes du substrat, ce qui induit le repliement du substrat. L'hydrolyse de l'ATP et la liaison d'une nouvelle protéine substrat dans la cavité opposée envoie un signal allostérique qui provoque la libération de GroES et d'une protéine encapsulée dans le cytosol. Une protéine donnée subira plusieurs cycles de repliement, retournant à chaque fois à son état non-replié original, jusqu'à ce que la conformation native ou une structure intermédiaire destinée à atteindre l'état natif soit produite. De manière alternative, le substrat peut être dénaturé lors d'une réaction compétitive, comme un mauvais repliement ou une agrégation avec d'autres protéines mal repliées[2].
Thermodynamique de la réaction
La nature contractée de l'intérieur du complexe moléculaire favorise fortement les conformations moléculaires compactes de la protéine substrat. En solution, les interactions apolaires et à longues portées peuvent seulement se produire dans ce cas avec un coût important en entropie. Dans les espaces fermés du complexe GroEL, la perte relative d'entropie est beaucoup plus faible. La méthode de capture tends aussi à concentrer les sites liants apolaires séparément des sites polaires. Lorsque les surfaces apolaires de GroEL sont ôtées, la chance pour qu'un quelconque groupe apolaire donné puisse rencontrer un site apolaire intramoléculaire sont bien plus importantes qu'en solution. Les sites hydrophobes se situant à l'extérieur sont rassemblés au sommet du domaine cis et liés entre eux. La géométrie de GroEL impose que les structures polaires soient motrices, et elles enveloppent le cœur apolaire lorsqu'il émerge depuis le côté trans.
Structure
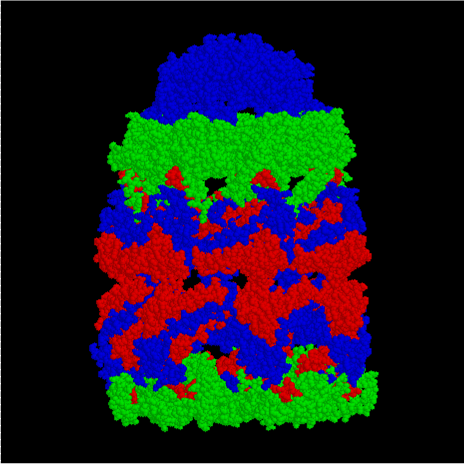
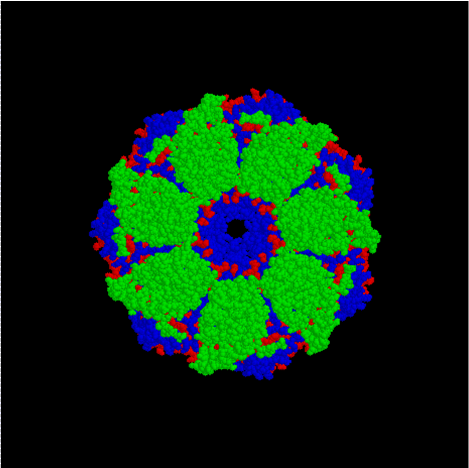
GroEL est, structurellement, un tétradécamère à anneaux doubles, les anneaux cis et trans étant constitués de sept sous-unités chacun. L'intérieur de GroEL est hydrophobe, et constitue l'endroit où le repliement de protéine est censé se faire.
-

GroEL (côté)
-

GroEL (dessus)
-

Complexe GroES/GroEL (côté)
-
Complexe GroES/GroEL (dessus)
La clé de l'activité de GroEL réside dans la structure du monomère. Le monomère Hsp60 possède trois parties distinctes séparées par deux régions charnières. La région apicale contient un grand nombre de sites liants pour les substrats pour protéines non repliées. De nombreuses protéines globulaires ne se lieront pas à cette région apicale car leurs parties hydrophobes sont internes, loin du milieu aqueux, dans une conformation favorisée par la thermodynamique. En conséquence, ces « sites substrats » ne se lieront qu'avec des protéines qui ne sont pas repliées de manière optimale. La région apicale possède aussi des sites liants pour les monomères Hsp10 de GroES.
Le domaine équatorial possède un emplacement près du point charnière pour lier l'ATP, ainsi que deux points de rattachement pour l'autre moitié de la molécule GroEL. Le reste de la région équatoriale est modérément hydrophile.
L'ajout de l'ATP et de GroES induit un effet drastique sur la conformation de la région cis. Il est causé par le pliement et la rotation des monomères Hsp60 aux points charnières. La région intermédiaire se replie vers l'intérieur, d'environ 25° par rapport à la charnière la plus basse. Cet effet, multiplié par le pliement coopératif des monomères, accroît le diamètre équatorial de la cage GroEL. Mais le domaine apical pivote de 60° vers l'extérieur par rapport à la charnière suppérieure, et aussi de 90 ° autour de l'axe charnière. Ce mouvement ouvre très largement la cage au dessus de la région cis, mais supprime l'ensemble des sites liant de l'intérieur de la cage.
Interactions
Il a été montré que GroEL interagit avec GroES[3],[4], l'ALDH2[4], la caspase 3[3],[5] et la reductase dihydrofolate[6].
Voir aussi
Références
- (en) Zeilstra-Ryalls J, Fayet O, Georgopoulos C, « The universally conserved GroE (Hsp60) chaperonins », dans Annu Rev Microbiol., vol. 45, 1991, p. 301 [lien PMID, lien DOI]
- (en) Horwich AL, Fenton WA, Chapman E, Farr GW, « Two Families of Chaperonin: Physiology and Mechanism », dans Annu Rev Cell Dev Biol., vol. 23, 2007, p. 115 [lien PMID, lien DOI]
- (en) Samali A, Cai J, Zhivotovsky B, Jones D P, Orrenius S, « Presence of a pre-apoptotic complex of pro-caspase-3, Hsp60 and Hsp10 in the mitochondrial fraction of jurkat cells », dans EMBO J., Angleterre, vol. 18, no 8, 1999, p. 2040–8 (ISSN 0261-4189) [lien PMID, lien DOI]
- (en) Lee Ki-Hwan, Kim Ho-Seung, Jeong Han-Seung, Lee Yong-Sung, « Chaperonin GroESL mediates the protein folding of human liver mitochondrial aldehyde dehydrogenase in Escherichia coli », dans Biochem. Biophys. Res. Commun., États-Unis d'Amérique, vol. 298, no 2, 2002, p. 216–24 (ISSN 0006-291X) [lien PMID]
- (en) Xanthoudakis S, Roy S, Rasper D, Hennessey T, Aubin Y, Cassady R, Tawa P, Ruel R, Rosen A, Nicholson D W, « Hsp60 accelerates the maturation of pro-caspase-3 by upstream activator proteases during apoptosis », dans EMBO J., Angleterre, vol. 18, no 8, 1999, p. 2049–56 (ISSN 0261-4189) [lien PMID, lien DOI]
- (en) Mayhew M, da Silva A C, Martin J, Erdjument-Bromage H, Tempst P, Hartl F U, « Protein folding in the central cavity of the GroEL-GroES chaperonin complex », dans Nature, Angleterre, vol. 379, no 6564, 1996, p. 420–6 (ISSN 0028-0836) [lien PMID, lien DOI]
A lire également
- (en) Tabibzadeh S, Broome J, « Heat shock proteins in human endometrium throughout the menstrual cycle », dans Infectious diseases in obstetrics and gynecology, vol. 7, no 1-2, 1999, p. 5–9 [lien PMID, lien DOI]
- (en) Schäfer C, Williams JA, « Stress kinases and heat shock proteins in the pancreas: possible roles in normal function and disease », dans J. Gastroenterol., vol. 35, no 1, 2000, p. 1–9 [lien PMID]
- (en) Moseley P, « Stress proteins and the immune response », dans Immunopharmacology, vol. 48, no 3, 2000, p. 299–302 [lien PMID, lien DOI]
- (en) Liu Y, Steinacker JM, « Changes in skeletal muscle heat shock proteins: pathological significance », dans Front. Biosci., vol. 6, 2001, p. D12–25 [lien PMID, lien DOI]
- (en) Van Maele B, Debyser Z, « HIV-1 integration: an interplay between HIV-1 integrase, cellular and viral proteins », dans AIDS reviews, vol. 7, no 1, 2005, p. 26–43 [lien PMID]
- (en) Hochstrasser DF, Frutiger S, Paquet N, et al., « Human liver protein map: a reference database established by microsequencing and gel comparison », dans Electrophoresis, vol. 13, no 12, 1993, p. 992–1001 [lien PMID, lien DOI]
- (en) Ikawa S, Weinberg RA, « An interaction between p21ras and heat shock protein hsp60, a chaperonin », dans Proc. Natl. Acad. Sci. U.S.A., vol. 89, no 6, 1992, p. 2012–6 [lien PMID, lien DOI]
- (en) Brudzynski K, Martinez V, Gupta RS, « Immunocytochemical localization of heat-shock protein 60-related protein in beta-cell secretory granules and its altered distribution in non-obese diabetic mice », dans Diabetologia, vol. 35, no 4, 1992, p. 316–24 [lien PMID, lien DOI]
- (en) Dawson SJ, White LA, « Treatment of Haemophilus aphrophilus endocarditis with ciprofloxacin », dans J. Infect., vol. 24, no 3, 1992, p. 317–20 [lien PMID, lien DOI]
- (en) Singh B, Patel HV, Ridley RG, et al., « Mitochondrial import of the human chaperonin (HSP60) protein », dans Biochem. Biophys. Res. Commun., vol. 169, no 2, 1990, p. 391–6 [lien PMID, lien DOI]
- (en) Venner TJ, Singh B, Gupta RS, « Nucleotide sequences and novel structural features of human and Chinese hamster hsp60 (chaperonin) gene families », dans DNA Cell Biol., vol. 9, no 8, 1991, p. 545–52 [lien PMID, lien DOI]
- (en) Ward LD, Hong J, Whitehead RH, Simpson RJ, « Development of a database of amino acid sequences for human colon carcinoma proteins separated by two-dimensional polyacrylamide gel electrophoresis », dans Electrophoresis, vol. 11, no 10, 1991, p. 883–91 [lien PMID, lien DOI]
- (en) Jindal S, Dudani AK, Singh B, et al., « Primary structure of a human mitochondrial protein homologous to the bacterial and plant chaperonins and to the 65-kilodalton mycobacterial antigen », dans Mol. Cell. Biol., vol. 9, no 5, 1989, p. 2279–83 [lien PMID]
- (en) Waldinger D, Eckerskorn C, Lottspeich F, Cleve H, « Amino-acid sequence homology of a polymorphic cellular protein from human lymphocytes and the chaperonins from Escherichia coli (groEL) and chloroplasts (Rubisco-binding protein) », dans Biol. Chem. Hoppe-Seyler, vol. 369, no 10, 1989, p. 1185–9 [lien PMID]
- (en) Kreisel W, Hildebrandt H, Schiltz E, et al., « Immuno-gold electron microscopical detection of heat shock protein 60 (hsp60) in mitochondria of rat hepatocytes and myocardiocytes », dans Acta Histochem., vol. 96, no 1, 1994, p. 51–62 [lien PMID]
- (en) Corbett JM, Wheeler CH, Baker CS, et al., « The human myocardial two-dimensional gel protein database: update 1994 », dans Electrophoresis, vol. 15, no 11, 1995, p. 1459–65 [lien PMID, lien DOI]
- (en) Baca-Estrada ME, Gupta RS, Stead RH, Croitoru K, « Intestinal expression and cellular immune responses to human heat-shock protein 60 in Crohn's disease », dans Dig. Dis. Sci., vol. 39, no 3, 1994, p. 498–506 [lien PMID, lien DOI]
- (en) Vélez-Granell CS, Arias AE, Torres-Ruíz JA, Bendayan M, « Molecular chaperones in pancreatic tissue: the presence of cpn10, cpn60 and hsp70 in distinct compartments along the secretory pathway of the acinar cells », dans J. Cell. Sci., vol. 107 ( Pt 3), 1994, p. 539–49 [lien PMID]
- (en) Mayhew M, da Silva AC, Martin J, et al., « Protein folding in the central cavity of the GroEL-GroES chaperonin complex », dans Nature, vol. 379, no 6564, 1996, p. 420–6 [lien PMID, lien DOI]
- (en) Tabibzadeh S, Kong QF, Satyaswaroop PG, Babaknia A, « Heat shock proteins in human endometrium throughout the menstrual cycle », dans Hum. Reprod., vol. 11, no 3, 1996, p. 633–40 [lien PMID]
Liens externes
- (es) Palaeos Bacteria: Pieces: GroEL (Pas de droits réservés)
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « GroEL » (voir la liste des auteurs)
-


Wikimedia Foundation. 2010.