- Deinonychus
-
 Deinonychus
Deinonychus
Squelette d'un Deinonychus Classification Règne Animalia Classe Reptilia Ordre Saurischia Sous-ordre Theropoda Super-famille Coelurosauria Famille Dromaeosauridae Genre Deinonychus
Ostrom, 1969Deinonychus (du grec: δεινός: « terrifiant » et ὄνυξ : « griffe ») est un genre de dinosaure carnivore de la famille des dromaeosauridés. Il y en a une espèce de décrite : Deinonychus antirrhopus. Ce dinosaure long d'environ 3,4 m a vécu au début du Crétacé, entre environ 115 et 108 millions d'années (du milieu de l'Aptien au début de l'Albien). Les fossiles ont été découverts dans les États américains du Montana, du Wyoming et de l'Oklahoma, dans les roches des Formations de Cloverly et d'Antlers. Des dents qui peuvent appartenir au genre Deinonychus ont été trouvées beaucoup plus à l'est dans le Maryland.
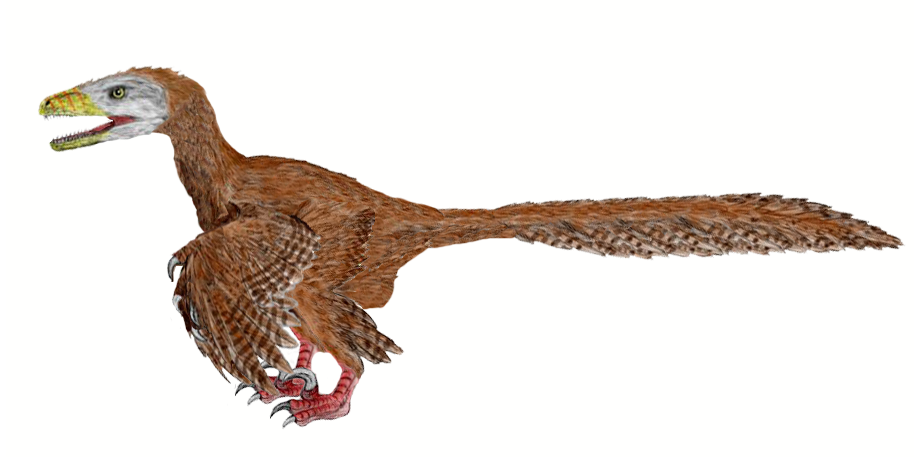
L'étude du Deinonychus par le paléontologue John Ostrom à la fin des années 1960 a révolutionné la façon de penser des scientifiques sur les dinosaures, conduisant à la « renaissance des dinosaures » et rallumant le débat pour savoir si les dinosaures étaient des animaux à sang chaud. Avant cette étude, la conception populaire faisait des dinosaures des sortes de reptiles géants peinant à se déplacer. Ostrom a remarqué que Deinonychus avait un corps, petit, harmonieux, à la posture horizontale, à la colonne vertébrale ressemblant à celle des ratites et surtout ayant aux pattes arrière de grandes griffes comme les rapaces ce qui suggère qu'il s'agissait d'un prédateur actif et agile[1].
Son nom de genre,« Griffe terrifiante », fait référence à l'inhabituelle longue phalange en forme de faucille trouvée sur le deuxième orteil de chaque patte arrière. Le fossile YPM 5205 possède cette grande griffe fortement courbée. Les archosauriens actuels ont un étui corné qui s'étend sur toute la longueur de l'os. Ostrom a étudié les griffes de crocodiles et d'oiseaux et reconstruit de là les griffes de YPM 5205 qui devaient mesurer plus de 12 centimètres de long[1]. Le nom d'espèce antirrhopus qui signifie « contrepoids », renvoie à l'idée que se faisait Ostrom sur le rôle de la queue. Comme chez d'autres dromæosauridés, les vertèbres de la queue ont une série de tendons ossifiés et de processus osseux très allongée. Ces caractéristiques semblaient faire jouer à la queue un rôle de contrepoids raide, mais un fossile de Velociraptor mongoliensis (IGM 100/986) très étroitement apparenté avait une queue courbée' latéralement en forme de S. Ceci suggère que, dans la vie, la queue pouvait se plier latéralement jusqu'à un degré élevé de flexibilité[2].
Dans les deux formations de Cloverly et d'Antlers, les restes de Deinonychus ont été retrouvés étroitement liés à celles de l'ornithopode Tenontosaurus. Des dents de Deinonychus ont été découvertes avec des Tenontosaurus ce qui signifie que ce dernier devait être chassé ou au moins piégé par les Deinonychus.
Sommaire
Description
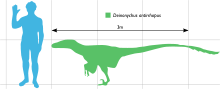 Sa taille comparée à celle d'un humain
Sa taille comparée à celle d'un humain
En se basant sur les plus grands spécimens connus, on peut estimer que Deinonychus devait mesurer au moins 3,4 mètres de long, avoir une tête longue de 41 cm, une taille à la hanche de 0,87 m et peser aux environs de 73 kg[3]. Il avait de puissantes mâchoires munies d'une soixantaine de dents courbes, en forme de lame. Les études de son crâne l'ont beaucoup fait évoluer au fil des décennies. Ostrom avait reconstruit le crâne partiel, imparfaitement conservé, qu'il avait comme triangulaire, large et assez semblable à celui d’Allosaurus. De nouveaux crânes de Deinonychus et d'espèces étroitement apparentées où les trois dimensions du crâne étaient mieux conservées[4] montrent que le palais était plus voûté, la face beaucoup plus étroite qu'Ostrom ne le pensait, tandis que les os jugaux étaient plus évasés, lui permettant d'une meilleure vision stéréoscopique. Le crâne de Deinonychus était toutefois différent de celui du Velociraptor car le sommet de son crâne était plus solide, comparable à celui de Dromaeosaurus et il n'avait pas les os nasaux déprimés de Velociraptor[5]. Le crâne, y compris la mandibule, avait des fenestrae (ouvertures du crâne) qui en réduisait le poids. Chez Deinonychus, la fenêtre antéorbitaire, une ouverture du crâne située entre l'œil et la narine, était particulièrement importante[4].
Deinonychus possédait de grandes mains (manus) avec une griffes sur chacun des trois doigts des patte avant. Le premier doigt était le plus court et la deuxième le plus long. Chaque patte arrière portait une griffe en forme de faucille sur le deuxième doigt, qui était probablement utilisée au cours de la prédation[6].
On n'a trouvé aucune plume en association avec des fossiles de Deinonychus. Néanmoins, des preuves suggèrent que les Dromaeosauridae, dont Deinonychus, avaient des plumes[7]. Le genre Microraptor est à la fois plus ancien géologiquement et plus primitif phylogénétiquement que Deinonychus et est placé dans la même famille[8]. De multiples fossiles de Microraptors ont gardé des plumes pennées comme celles retrouvées chez les oiseaux actuels, au niveau des pattes avant, arrière et de la queue, aussi bien que pour les plumes de couverture[7]. Velociraptor est géologiquement plus jeune que Deinonychus mais y est plus étroitement apparenté (tous deux appartiennent à la sous-famille des velociraptorinae, voir classification ci-dessous). Un spécimen de Velociraptor a été trouvé avec des bourgeons de plume sur ses cubitus. Ces bourgeons sont des excroissances osseuses située le long de l'os où s'inséraient les ligaments folliculaires et sont un indicateur direct de la présence de plumes d'aspect moderne[9].
Classification
 Représentation hypothétique d'un Deinonychus avec des plumes
Représentation hypothétique d'un Deinonychus avec des plumesDeinonychus est l'un des dromæosauridés les mieux connus[10] et est un proche parent du plus petit Velociraptor, trouvé dans les terrains plus jeunes de la fin du Crétacé en Asie centrale[11],[12]. Ils forment tous deux un clade appelé Velociraptorinae. Ce nom de Velociraptorinae a été inventé par Rinchen Barsbold en 1983[13] et contenait à l'origine le seul genre Velociraptor. Plus tard, Phil Currie y a inclus la plupart des dromæosauridés[14]. Deux genres datant du Crétacé supérieur, Tsaagan trouvé en Mongolie[11] et Saurornitholestes trouvé en Amérique du Nord[3], peuvent également être des proches, mais ce dernier est mal connu et difficile à classer[11]. Velociraptor et ses alliés sont considérés comme utilisant plus leurs griffes que leurs dents pour tuer, au contraire des dromæosauridés comme Dromaeosaurus avec leurs têtes trapues[15]. Avec les troodontidés, les dromæosauridés forment le clade des Deinonychosauria qui est un taxon frère des Aves. Phylogénétiquement, les Deinonychosauria forment le groupe de dinosaures non-aviaires les plus étroitement liés aux oiseaux[16].
Découverte et dénomination
Des restes fossilisés de Deinonychus ont été récupérés dans la Formation de Cloverly au Montana et au Wyoming[1] et dans la formation à peu près contemporaine d'Antlers dans l'Oklahoma[17] en Amérique du Nord. La formation de Cloverly a été datée comme allant de l'Aptien supérieur au début de l'Albien au Crétacé inférieur, il y a environ 115 à 108 Ma[18],[19]. En outre, des dents trouvées dans le faciès d'Arundel Clay (mi-Aptien), dans la Formation de Potomac dans les plaines côtières atlantiques du Maryland peuvent être considérées comme appartenant au genre Deinonychus[20].
Les premiers restes ont été découverts en 1931 dans le sud du Montana près de la ville de Bridger. Le responsable de l'équipe, le paléontologue Barnum Brown, avait principalement pour consigne de dégager et de préparer la dépouille d'un dinosaure ornithopode Tenontosaurus mais, dans son rapport sur le site de fouilles fait à l'American Museum of Natural History, il signala la découverte d'un petit dinosaure carnivore à proximité du squelette du Tenontosaurus, mais pris dans le calcaire et difficile à préparer[21]. Il appela l'animal de manière informelle Daptosaurus et se prépara à le décrire lorsqu'il aurait le squelette à disposition, mais il ne termina jamais ce travail[22]. Brown ramena de la Formation de Cloverly le squelette d'un petit théropode aux dents apparemment surdimensionnées qu'il appela officieusement Megadontosaurus. John Ostrom, examina les pièces rapprtées des décennies plus tard et se rendit compte que les dents venaient d'un Deinonychus mais que le reste du squelette venaitt d'un animal complètement différent. Il appela ce squelette Microvenator[22].
Un peu plus de trente ans plus tard, en août 1964, le paléontologue John Ostrom mena une expédition pour le Peabody Museum de l'université Yale qui découvrit de nouveaux matériels squelettiques. Des expéditions au cours des deux étés suivants ont découvert plus de 1 000 nouveaux os, provenant d'au moins trois individus. Comme les possibilités d'association entre les divers os récupérés était faible, rendant impossible de déterminer avec certitude de quel animal les pièces provenaient, le spécimen type (YPM 5205) de Deinonychus se limita à un pied gauche complet et une partie de pied droit qui ont certainement appartenu au même individu[6]. Les autres échantillons ont été catalogués dans cinquante entrées distinctes au Peabody Museum.
Plus tard, Ostrom et Grant E. Meyer ont étudié leur propre matériel ainsi que le Daptosaurus de Brown en détail et ont trouvé qu'ils appartenaient à la même espèce. Ostrom a publié ses conclusions en 1969, donnant aux pièces trouvées le nouveau nom de Deinonychus antirrhopus[6]. « antirrhopus » signifie «contrepoids» et fait réfèrence à la très probable queue raide.
Bien que d'une myriade d'os ait été disponible en 1969, de nombreux os parmi les plus importants étaient manquants ou difficiles à interpréter. Il y avait peu d'éléments du crâne postorbitaire, pas de fémur, pas de sacrum, pas de furcula ou du sternum, il manquait des vertèbres et (du moins Ostrom le pensait) il ne disposait que d'un petit fragment d'une coracoïde. La reconstruction du squelette de Deinonychus par Ostrom incluait un os du pubis très rare, de forme trapézoïdale et plat, contrairement à celui des autres théropodes, mais qui avait la même longueur que l'ischion trouvé juste à côté.
De nouvelles découvertes
 Vue d'artiste
Vue d'artisteEn 1974, Ostrom publia une monographie sur l'épaule de Deinonychus où il expliquait s'être rendu compte que ce qu'il avait décrit comme le pubis était en fait un coracoïde -. un os de l'épaule[23]. La même année, un autre spécimen de Deinonychus était dégagé dans le Montana par une expédition de l'Université Harvard dirigée par Farish Jenkins. Cette découverte apporta plusieurs éléments nouveaux : des fémurs bien préservés, des pubis, un sacrum et des ilions en meilleur état ainsi que des éléments des pieds et du métatarse. Ostrom décrivit le spécimen et révisa sa conception du squelette de Deinonychus. Cette fois, il montra un pubis très long, et Ostrom commença à soupçonner qu'il pouvait même avoir été un peu rétroversé comme celui des oiseaux[24].
Un squelette de Deinonychus avec des os véritables et le plus complet des spécimens peut être vu à l’American Museum of Natural History[25], un autre spécimen étant exposé au Museum of Comparative Zoology de l'Université Harvard. Les spécimens de l’American Museum et de l'Université Harvard sont d'une origine différente de celle des spécimens de Yale. Cependant il manque à ces deux reconstitutions des éléments comme les sternums, des côtes thoraciques, des furcula et gastralia.
Cependant, après tous les travaux d'Ostrom, plusieurs petits blocs de matériau engainés de calcaire étaient restés stockés à l’American Museum. Il s'agissait principalement d'os isolés et de fragments d'os pris dans la matrice rocheuse ainsi que de fragments rocheux avoisinants. L'examen de ces blocs préparés par Gerald Grellet-Tinner et Peter Makovicky en 2000 a révélé des détails intéressants qui avaient été négligés. Plusieurs os longs, minces identifiés sur les blocs comme des tendons ossifiés (structures qui contribuaient à raidir la queue de Deinonychus) se sont révélés être des gastralia (côtes abdominales). Plus important encore, un grand nombre de coquilles d'œufs fossilisés qui étaient auparavant imperceptibles ont été découvertes dans la matrice rocheuse qui avait entouré l'échantillon original de Deinonychus[26].
Dans un rapport plus détaillé paru par la suite sur les coquilles d'œufs, Grellet-Tinner et Makovicky ont conclu que les œufs étaient sûrement ceux de Deinonychus, en faisant les premiers œufs de Dromaeosauridés à être identifiés[21]. En outre, la surface externe d'un œuf a été trouvée en étroit contact avec des gastralia suggérant que Deinonychus pourrait avoir couvé ses œufs. Cela implique que Deinonychus utilisait le transfert de chaleur de son corps comme mécanisme d'incubation, un système analogue à celui des oiseaux actuels[27]. Une étude plus approfondie par Gregory Erickson et ses collègues constate que cet individu avait 13 ou 14 ans au moment de son décès et que sa croissance avait atteint un plateau. Contrairement à ce qui a été observé chez d'autres théropodes, dans des études de spécimens trouvés associés à des œufs ou des nids, celui-ci avait fini de grandir au moment de sa mort[28].
Conséquences
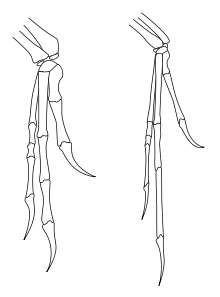 La similitude des membres antérieurs de Deinonychus (à gauche) avec ceux deArchaeopteryx (à droite) a conduit John Ostrom à relancer le lien entre les dinosaures et les oiseaux
La similitude des membres antérieurs de Deinonychus (à gauche) avec ceux deArchaeopteryx (à droite) a conduit John Ostrom à relancer le lien entre les dinosaures et les oiseauxLa description de Deinonychus faite en 1969 par Ostrom a souvent été considérée comme la découverte la plus importante de la paléontologie des dinosaures du milieu du XXe siècle[29]. La découverte de ce prédateur manifestement actif et agile a beaucoup fait pour changer la conception des scientifiques (et populaire) sur les dinosaures et a ouvert la porte à la spéculation que les dinosaures auraient pu être des animaux à sang chaud. Cette évolution a été appelée la renaissance des dinosaures. Plusieurs années plus tard, Ostrom a noté des similitudes entre les pattes antérieures de Deinonychus et des oiseaux, une observation qui l'a amené à relancer l'hypothèse que les oiseaux descendent des dinosaures[30]. Quarante ans plus tard, cette idée est presque universellement acceptée.
En raison de son anatomie très ressemblante à celle des oiseaux et de ses relations étroites avec d'autres dromæosauridés, les paléontologues ont émis l'hypothèse que Deinonychus était probablement couvert de plumes[31],[32],[33]. Alors que l'on a des preuves que plusieurs dromæosauridés comme Velociraptor et Microraptor avaient des plumes analogues à celles des oiseaux actuels, il n'existe à ce jour aucune preuve directe pour Deinonychus lui-même[7],[9]. Quand les paléontologues comme Phil Senter ont étudié des domaines tels que l'amplitude des mouvements dans les membres antérieurs, ils ont pris en compte la présence probable de plumes au niveau des ailes[34].
Paléobiologie et paléoécologie
Les données géologiques indiquent que Deinonychus habitait des plaines inondables ou des zones de marais[10]. Le paléoenvironnement des Formations de Cloverly et d'Antlers où ont été trouvés les restes de Deinonychus était constitué de forêts d'origine tropicale ou sub-tropicale, de deltas et de lagunes et n'était pas sans rappeler la Louisiane d'aujourd'hui[35],[36]. Les Deinonychus partageaient leur milieu avec notamment les dinosaures herbivores tels que les Sauropelta et leurs armures et les ornithopodes Zephyrosaurus et Tenontosaurus. Dans l'Oklahoma, l'écosystème de Deinonychus incluait également le grand théropode Acrocanthosaurus, l'énorme sauropode Sauroposeidon, le crocodilien Goniopholis et le sémionotiforme Lepisosteus[36]. Si les dents trouvées dans le Maryland sont celles de Deinonychus, alors ses voisins comprenaient également Astrodon et un nodosaure (appelé Priconodon) connu seulement à partir de ses dents. La partie centrale de la Formation de Cloverly peut être datée de 115 ± 10 Ma près de la base[18] à 108,5 ± 0,2 Ma près du sommet[19].
Le comportement prédateur
 Représentation d'artiste d'un couple de Deinonychus se nourrissant d'une carcasse.
Représentation d'artiste d'un couple de Deinonychus se nourrissant d'une carcasse.Des dents de Deinonychus ont souvent été trouvées associées à des fossiles du dinosaure ornithopode Tenontosaurus dans la Formation de Cloverly. Deux sites ont été découverts qui possèdent des fossiles assez complets de Deinonychus près de fossiles de Tenontosaurus. Le premier, la carrière de Yale dans la Formation de Cloverly du Montana, comprend de nombreuses dents, quatre squelettes d'adultes et un de juvénile. L'association de plusieurs squelettes de Deinonychus dans un même lieu à proximité d'un Tenontosaurus donne à penser que les premiers ont pu se nourrir du second et l'ont même peut-être chassé. Ostrom et Maxwell ont même utilisé cette information pour spéculer que Deinonychus aurait pu vivre et chasser en meute[37]. La seconde carrière est située dans la Formation d'Antlers en Oklahoma. Le site contient six squelettes partiels de Tenontosaurus de différentes tailles, avec un squelette partiel et de nombreuses dents de Deinonychus. Un humérus de Tenontosaurus porte même ce qui pourrait être des marques de dents de Deinonychus. Brinkman et al. (1998) soulignent qu'un Deinonychus adulte pesait entre 70 et 100 kg tandis que les Tenontosaurus adultes pesaient entre 1 et 4 tonnes. Un Deinonychus solitaire ne pouvait pas tuer un Tenontosaurus adulte, ce qui suggère qu'il savait chasser en groupe[17].
Une étude récente de Roach et Brinkman a remis en question le comportement de chasse en groupe des Deinonychus, en se basant sur ce qui est connu des méthodes de chasse des carnivores modernes et de la taphonomie des sites de Tenontosaurus. Les archosauriens actuels (les oiseaux et les crocodiles) et le dragon de Komodo ont peu tendance à chasser en groupe ; au contraire, ce sont généralement soit des chasseurs solitaires, soit ils sont attirés par des carcasses d'animaux déjà tués ce qui engendre beaucoup de conflits entre individus de la même espèce. Par exemple, dans des situations où des groupes de dragons de Komodo mangent ensemble, les plus gros individus mangent en premier et attaquent les petits dragons qui tentent de se nourrir et, si le petit animal est tué, il est cannibalisé. Lorsque cette observation est appliquée aux sites de fossiles de Tenontosaurus, il semble que ce qui est trouvé est compatible avec les Deinonychus ayant une stratégie alimentaire analogue à celle des dragons de Komodo ou des crocodiles. Les squelettes de Deinonychus découverts sur ces sites sont de jeunes adultes, auxquels il manque des pièces osseuses situation compatible au fait qu'ils ont pu être mangés par d'autres Deinonychus[38]. D'autre part, un article de Li et al .. décrit des sites d'empreintes de pas avec des enjambées régulières et des voies de cheminement parallèles, ce qui implique un comportement grégaire d'attaque au lieu d'un comportement alimentaire coordonné[39].
Force de morsure
 Crâne de Deinonychus.
Crâne de Deinonychus.La première estimation de la force de morsure d'un Deinonychus a été faite en 2005, en se basant sur la musculature d'une mâchoire reconstruite. Cette étude a conclu que Deinonychus avait probablement eu une force maximale de morsure de 15 % de celle d'un alligator moderne[40]. Une étude faite en 2010 par Paul Gignac et ses collègues a tenté d'estimer la force de morsure en se basant directement sur les marques de dents de Deinonychus nouvellement découvertes dans les os d'un Tenontosaurus. Les marques de morsures provenaient d'un adulte de grande taille et ont fourni les premières preuves que Deinonychus pouvait perforer un os. En utilisant ces empreintes, l'équipe de Gignac a été en mesure de déterminer que la force de morsure de Deinonychus était significativement plus élevée que les études antérieures biomécaniques seules ne l'avaient estimé. Ils ont trouvé une force se situant entre 4 100 et 8 200 newtons, plus que les mammifères carnivores actuels, y compris la hyène, et équivalente à celle d'un alligator de taille similaire.
Cependant, Gignac et ses collègues ont également noté que les marques de perforation d'os par Deinonychus sont relativement rares, contrairement aux nombreuses marques de perforation connues dues à de grands théropodes comme Tyrannosaurus. Deinonychus ne devait probablement pas souvent briser ou manger des os. Au lieu de cela, il a probablement utilisé sa force de morsure pour se défendre ou attaquer ses proies mais pas pour se nourrir[41].
Rôle des membres
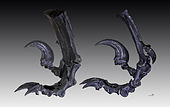 Moulage des os du pied montrant la griffe en faucille du deuxième orteil.
Moulage des os du pied montrant la griffe en faucille du deuxième orteil.Alors qu'elle est l'élément le plus caractéristique de Deinonychus, la forme et la courbure de la griffe du deuxième orteil varie selon les spécimens. Le spécimen type décrit par Ostrom en 1969 a des griffes fortement courbées tandis qu'un individu décrit en 1976 avait des griffes avec une courbure beaucoup plus faible, plus proche de celle des griffes « normales » des autres orteils[24]. Ostrom a suggéré que cette différence de taille et de forme pouvait être due à des variations individuelles, dues au sexe ou liées à l'âge, mais il a admis qu'il ne pouvait pas en être sûr.
On a des preuves anatomiques[1] et par les empreintes de pas[42] que cette griffe n'entrait pas en contact avec le sol tandis qu'ils marchaient sur les troisième et quatrième orteils.
Ostrom a suggéré que Deinonychus pourrait utiliser ses grandes griffes pour couper et dilacérer ses proies[1]. Certains chercheurs ont même suggéré qu'elle était utilisée pour éventrer les grands dinosaures cératopsiens[43]. D'autres études ont suggéré que ces griffes n'étaient pas été utilisés pour dilacérer mais plutôt pour porter de petits coups à la victime[44]. Manning et al. (2005) ont effectué des essais sur une réplique robotique qui correspondait précisément à l'anatomie de Deinonychus et Velociraptor et utilisé un robot muni de vérins hydrauliques pour frapper une carcasse de porc. Au cours de ces essais, les serres n'ont fait que des perforations peu profondes et ne pouvaient pas couper ou dilacérer. Les auteurs suggèrent que les griffes servaient plus à grimper sur les proies qu'à les tuer[45].
Ostrom a comparé Deinonychus à l'autruche et au casoar. Il a noté que ces deux espèces d'oiseaux pouvaient infliger de graves blessures grâce à la grande griffe de leur deuxième orteil[1]. Le casoar a des griffes pouvant atteindre jusqu'à 125 millimètres de long[46]. Ostrom a cité Gilliard (1958) en disant qu'elles peuvent couper un bras ou éventrer un homme[47]. Kofron (1999 et 2003) a étudié 241 attaques documentées de casoar et a constaté qu'un homme et deux chiens ont été tués, mais qu'il n'existait aucune preuve que les casoars pouvaient éventrer ou démembrer les autres animaux[48],[49]. Les casoars utilisent leurs griffes pour se défendre, attaquer des animaux menaçants et dans les combats entre mâles en période de reproduction[46]. Les Cariamas ont aussi une grande griffe au deuxième orteil. Ils l'utilisent pour déchirer leurs proies en petits morceaux avant de les avaler[50].
Des études biomécaniques menées par Ken Carpenter en 2002 ont confirmé que la fonction la plus probable des membres antérieurs était de saisir les proies, leurs grandes longueurs leur permettant d'avoir une plus grande portée que la plupart des autres théropodes. Le coracoïde assez large et allongé, permettant l'insertion de muscles puissants pour les membres antérieurs, vient encore renforcer cette interprétation[51]. Des études biomécaniques menées par Carpenter à l'aide de moulages d'os ont également montré que Deinonychus ne pouvait pas plier ses bras contre son corps comme un oiseau (« pliage aviaire ») contrairement à ce qui avait été déduit de la description précédente en 1985 par Jacques Gauthier[52] et en 1988 par Gregory S. Paul[3].
Des études menées par Phil Senter en 2006 montrent que Deinonychus pouvait utiliser ses membres antérieurs non seulement pour saisir ses proies, mais aussi pour ses œufs ou ses petits vers sa poitrine. Si Deinonychus avait eu des plumes aux membres antérieurs, celles-ci auraient limité l'amplitude de mouvement de ces membres jusqu'à un certain point. Par exemple, lorsque Deinonychus étendait son bras vers l'avant, la « paume » de sa main s'orientait automatiquement vers le haut. Cette situation aurait provoqué un blocage mutuel des deux membres antérieurs s'ils avaient été étendus en même temps, ce qui a conduit Senter à conclure que Deinonychus n'utilisait qu'un bras à la fois pour ramener sa couvée vers sa poitrine. Le rôle des doigts aurait aussi été limité par des plumes alors que, par exemple, le troisième doigt aurait pu être employé à des activités telles que l'exploration de l'intérieur de petites proies, ce seulement dans une position sensiblement perpendiculaire à l'aile[34]. Alan Gishlick, dans une étude de 2001 sur la mécanique des membres antérieurs de Deinonychus, a constaté que, même s'il avait eu de grandes plumes aux ailes, ses capacités de préhension n'auraient pas été significativement entravées, mais qu'il aurait du saisir les objets entre ses deux ailes dans une sorte de bear hug faits qui ont été confirmés dans des études ultérieures de Carpenter et Senter[53].
Parsons et Parsons ont montré que les juvéniles et les spécimens sub-adultes de Deinonychus présentaient quelques différences morphologiques avec les adultes. Par exemple, les bras des spécimens les plus jeunes sont proportionnellement plus longs que ceux des adultes, indiquant peut-être une différence de comportement entre jeunes et adultes[54]. Un autre exemple pourrait être le rôle des griffes des pattes arrière. Parsons et Parsons ont suggéré que la courbure des griffes (pour laquelle Ostrom, en 1976, avait montré l'existence d'une différence entre spécimens[24]) pourrait avoir été plus forte chez les jeunes Deinonychus, ce qui aurait pu les l'aider à grimper dans les arbres, et que les griffes devenaient plus droites lorsque l'animal devenait plus âgé et commençait à vivre uniquement sur le sol[55]. Cela est basé sur l'hypothèse que certains petits dromæosauridés utilisaient leurs griffes pour escalader[45].
Vitesse
Les Dromæosauridés, en particulier Deinonychus, sont souvent dépeints dans les médias comme des animaux exceptionnellement rapides et Ostrom a émis lui-même l'hypothèse dans sa description originale que Deinonychus était un animal véloces[6]. Cependant, lors de la première description, on ne disposait pas d'une jambe complète de Deinonychus et les spéculations d'Ostrom sur la longueur du fémur (os de la cuisse) s'avéreront surestimées. Dans une étude ultérieure, Ostrom a noté que le rapport de longueur du fémur par rapport au tibia (os de la jambe) n'est pas aussi important pour la détermination de la vitesse que la longueur relative du pied par rapport à la jambe. Chez les actuels oiseaux bons coureurs comme l'autruche, le rapport longueur du pied/longueur du tibia est de 0,95. Chez les rares dinosaures véloces comme Struthiomimus ce ratio est de 0,68 mais chez Deinonychus, le ratio n'est que de 0,48. Ostrom en a déduit que « la seule conclusion raisonnable » était que Deinonychus n'était pas particulièrement plus rapide que d'autres dinosaures, et certainement pas aussi rapide que les oiseaux coureurs modernes[24].
Ce faible rapport chez Deinonychus est du en partie à un métatarse (partie supérieure des os du pied) inhabituellement court. Ce rapport est en fait plus grand chez les petits individus que chez les grands. Ostrom a suggéré que le métatarse court pouvait être lié au rôle de la griffe en faucille, et utilisé le fait qu'il semble se raccourcir chez les individus âgés pour soutenir son hypothèse. Il a interprété toutes ces caractéristiques - un deuxième orteil court avec une longue griffe, un métatarse court, etc - pour soutenir qu'il utilisait sa patte arrière comme arme offensive, où la griffe, comme la jambe, orientée en bas et en arrière aurait servi à entailler et à dilacérer sa proie. Ostrom a suggéré qu'un métatarse court réduisait les contraintes sur l'os de la jambe lors d'une telle attaque et interprété la disposition inhabituelle des attaches musculaires sur la jambe de Deinonychus pour appuyer son idée que des groupes de muscles différents étaient utilisés pour courir et pour attraper les proies. Ostrom en a conclu que les jambes de Deinonychus représentaient un juste équilibre entre les adaptations nécessaires à en faire un prédateur rapide et une réduction des contraintes pour utiliser son pied comme seule arme[24].
Dans une étude en 1981 sur des traces de dinosaures canadiens, Richard Kool en est arrivé à estimer approximativement la vitesse de marche en étudiant les empreintes laissées par différentes espèces dans la formation de Gething en Colombie-Britannique. Kool a estimé que sur l'une des séries d'empreintes, l'ichnoespèce Irenichnites gracilis (qui pourrait être en fait Deinonychus), avait une vitesse de marche de 10 km/h[56].
Œufs
 Représentation d'artiste d'un individu en train de couvrer.
Représentation d'artiste d'un individu en train de couvrer.L'identification en 2000 d'un probable œuf de Deinonychus associé à l'un des individus du genre a permis de comparer sa structure, la nidification et la reproduction avec celle d'autres dinosaures théropodes. Lors de leur examen de l'échantillon en 2006, Grellet-Tinner et Makovicky ont examiné la possibilité que l’œuf ait servi à l'alimentation d'un dromaeosauridé ou que les fragments d'œufs aient été associés par hasard avec le squelette de Deinonychus. Ils ont écarté l'idée que l'œuf ait servi à un repas car aucun fragment n'était pris en sandwich entre les côtes abdominales et les os des membres antérieurs, ce qui rendait impossible qu'ils aient pu être contenus dans l'estomac de l'animal. En outre, la manière dont l'œuf avait été brisé indiquait qu'il était intact au moment de son enfouissement, et avait été brisé par un processus de fossilisation. L'idée que l'œuf ait été associé par hasard au dinosaure a été également considérée comme peu probable, les os entourant l'œuf n'ayant pas été dispersés ou désarticulés, mais étant restés dans leur position dans l'animal vivant, ce qui indique que la zone autour de l'œuf et l'œuf n'avaient pas été déplacés lors de leur conservation. Le fait que ces os étaient des côtes abdominales (gastralia), qui sont très rarement retrouvées, venaient à l'appui de cette interprétation. Tous les éléments de preuve, selon Grellet-Tinner et Makovicky, indiquent que l'œuf était probablement intact sous le corps de Deinonychus quand il a été enseveli. Il est possible que cela corresponde à une couvaison ou à un comportement de nidification de Deinonychus similaire à celui rencontré chez d'autres troodontidés et oviraptoridés, ou que l'œuf était en fait à l'intérieur de l'oviducte au moment où l'animal est mort[21].
L'examen de la microstructure de l'œuf de Deinonychus confirme qu'il appartenait bien à un théropode, car il partage des caractéristiques d'autres œufs de théropodes et montre des divergences avec des œufs d'ornithischiens et de sauropodes. Par rapport à d'autres théropodes maniraptoriens, l'œuf de Deinonychus est plus semblable à un œuf d'oviraptoridés qu'à celui d'un troodontidé, en dépit des études qui montrent que ces derniers sont plus étroitement liés aux dromæosauridés comme Deinonychus. Bien que l'œuf ait été trop écrasé pour en déterminer sa taille avec précision, Grellet-Tinner et Makovicky ont estimé son diamètre à environ 7 cm en se basant sur le diamètre du détroit pelvien par lequel l'œuf devait réussir à sortir. Cette taille est semblable aux 7,2 cm de la taille des œufs de Citipatis (des oviraptoridés); Citipati et Deinonychus avaient également environla même taille, ce qui vient appuyer cette estimation. En outre, l'épaisseur des coquilles d'œufs de Citipati et de Deinonychus sont presque identiques, et comme l'épaisseur de la coquille est en corrélation avec le volume de l'œuf, ceci vient encore plus appuyer l'idée que les œufs de ces deux animaux étaient de la même taille.
Galerie d'images
-

Le crâne d'un Deinonychus -

-

Annexes
Articles connexes
Bibliographie
Liens externes
Références taxinomiques
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Deinonychus » (voir la liste des auteurs)
- J. H. Ostrom, « Stratigraphy and paleontology of the Cloverly Formation(Lower Cretaceous) of the Bighorn Basin area, Wyoming and Montana », dans Bulletin of the Peabody Museum of Natural History, vol. 35, 1970, p. 1–234
- Mark A. Norell, « Important features of the dromaeosaurid skeleton II: information from newly collected specimens of Velociraptor mongoliensis », dans American Museum Novitates, vol. 3282, 1999, p. 1–45 [texte intégral]
- (en) G.S. Paul, Predatory Dinosaurs of the World, New York, Simon and Schuster, 1988 (ISBN 978-0-671-61946-6) (LCCN 88023052), p. 366–369
- W. D. Maxwell, « New Material of Deinonychus (Dinosauria, Theropoda) », dans Journal of Vertebrate Paleontology, vol. 16, no 3, 1996, p. 51A
- Lawrence M. Witmer, « The skull of Deinonychus (Dinosauria:Theropoda): New insights and implications », dans Journal of Vertebrate Paleontology, vol. 16, no 3, 1996, p. 73A
- J. H. Ostrom, « Osteology of Deinonychus antirrhopus, an unusual theropod from the Lower Cretaceous of Montana », dans Peabody Museum of Natural History Bulletin, vol. 30, 1969, p. 1–165 [texte intégral]
- X. Xu, « Four-winged dinosaurs from China », dans Nature, vol. 421, no 6921, 2003, p. 335–340 [lien PMID, lien DOI]
- S. H. Hwang, « New Specimens of Microraptor zhaoianus (Theropoda: Dromaeosauridae) from Northeastern China », dans American Museum Novitates, vol. 3381, 2002, p. 44 [al.2002.pdf texte intégral]
- A.H. Turner, « Feather quill knobs in the dinosaur Velociraptor », dans Science, vol. 317, no 5845, 2007, p. 1721 [lien PMID, lien DOI]
- (en) Norell, M.A., Makovicky, P.J., The Dinosauria, Berkeley, University of California Press, 2004 (ISBN 0-520-24209-2), « Dromaeosauridae », p. 196–210
- M.A. Norell, « A new dromaeosaurid theropod from Ukhaa Tolgod (Ömnögov, Mongolia) », dans American Museum Novitates, vol. 3545, 2006, p. 1–51 [texte intégral [PDF], lien DOI (pages consultées le 7 juillet 2007)]
- A.S. Turner, « A small derived theropod from Öösh, Early Cretaceous, Baykhangor Mongolia », dans American Museum Novitates, vol. 3557, 2007, p. 1–27 [texte intégral [PDF], lien DOI (pages consultées le 29 mars 2007)]
- R. Barsbold, « Carnivorous Dinosaurs from the Cretaceous of Mongolia », dans The Joint Soviet–Mongolian Palaeontological Expedition, Transactions, vol. 19, 1983, p. 1–117
- P. J. Currie, « New information on the anatomy and relationships of Dromaeosaurus albertensis (Dinosauria: Theropoda) », dans Journal of Vertebrate Paleontology, vol. 15, no 3, 1995, p. 576–591 [lien DOI] (abstract)
- (en) G.S. Paul, Predatory dinosaurs of the world : a complete illustrated guide, New York, Simon and Schuster, 1988 (ISBN 978-0-671-61946-6) (LCCN 88023052), p. 358
- (en) M.J. Benton, Vertebrate Palaeontology (Third Edition), Oxford, Blackwell Publishing, 2004, 3e éd., poche (ISBN 978-0-632-05637-8) (LCCN 2003028152) [lire en ligne], p. 472
- D. L. Brinkman, « First occurrence of Deinonychus antirrhopus (Dinosauria: Theropoda) from the Antlers Formation (Lower Cretaceous: Aptian–Albian) of Oklahoma », dans Oklahoma Geological Survey Bulletin, vol. 146, 1998, p. 1–27
- Z.-Q. Chen, « A fission track study of the terrigenous sedimentary sequences of the Morrison and Cloverly Formations in northeastern Bighorn Basin, Wyoming », dans The Mountain Geologist, vol. 34, 1997, p. 51–62
- D. Burton, « New radiometric ages from the Cedar Mountain Formation, Utah and the Cloverly Formation, Wyoming: implications for contained dinosaur faunas », dans Geological Society of America Abstracts with Programs, vol. 38, no 7, 2006, p. 52
- (en) T. R. Lipka, Lower and Middle Cretaceous Terrestrial Ecosystems, Albuquerque, New Mexico Museum of Natural History and Science, 1998 (OCLC 40283894), « The Affinities of the Enigmatic Theropods of the Arundel Clay Facies (Aptian), Potomac Formation, Atlantic Coastal Plain of Maryland », p. 229–234
- G. Grellet-Tinner, « A possible egg of the dromaeosaur Deinonychus antirrhopus: phylogenetic and biological implications », dans Canadian Journal of Earth Sciences, vol. 43, no 6, 2006, p. 705–719 [lien DOI]
- (en) M. A. Norell, Gaffney, E. S.; and Dingus, L., Discovering Dinosaurs in the American Museum of Natural History, New York, Knopf, 1995, 1re éd. (ISBN 978-0-679-43386-6) (LCCN 94046225), p. 126–130
- John H. Ostrom, « The Pectoral Girdle and Forelimb Function of Deinonychus (Reptilia: Saurischia) : A Correction », dans Postilla, Peabody Museum of Natural History Bulletin, vol. 165, 1974, p. 1–11
- J.H. Ostrom, « On a new specimen of the Lower Cretaceous theropod dinosaur Deinonychus antirrhopus », dans Breviora, vol. 439, 1976, p. 1–21 [texte intégral]
- American Museum of Natural History, « Deinonychus », http://www.amnh.org, American Museum of Natural History, 2007. Consulté le 13 juillet 2007
- (en) P. J. Makovicky, First international symposium on dinosaur eggs and babies, Isona i Conca Dellà Catalonia, Spain, 23–26 September 1999, 2000, « Association between a specimen of Deinonychus antirrhopus and theropod eggshell », p. 123–128
- Gerard Grellet-Tinner, « Oology And The Evolution Of Thermophysiology In Saurischian Dinosaurs: Homeotherm And Endotherm Deinonychosaurians? », dans Papeis Avulsos de Zoologia, vol. 46, no 1, 2006, p. 1–10 [texte intégral, lien DOI (pages consultées le 2007-07-07)]
- Gregory M. Erickson, « Growth patterns in brooding dinosaurs reveals the timing of sexual maturity in non-avian dinosaurs and genesis of the avian condition », dans Biology Letters, vol. published online, no 5, 2007, p. 558–61 [texte intégral [PDF], lien PMID, lien DOI (pages consultées le 2007-07-26)]
- (en) Fastovsky, D.E., Weishampel, D.B., The Evolution and Extinction of the Dinosaurs, Cambridge, Cambridge University Press, 2005 (ISBN 0-521-81172-4), « Theropoda I: Nature Red in Tooth and Claw », p. 265–299
- J. H. Ostrom, « Archaeopteryx and the origin of birds », dans Biological Journal of the Linnean Society, vol. 8, 1976, p. 91–182 [lien DOI]
- (en) R.T. Bakker, The Dinosaur Heresies, New York, Kensington Publishing, 1986, poche (ISBN 978-0-8065-2260-9), p. 310
- (en) John A. Long et P. Schouten, Feathered Dinosaurs: The Origin of Birds, Oxford, Oxford University Press, 2008 (ISBN 978-0-19-537266-3) (LCCN 2008001232), « Deinonychus », p. 142–143
- (en) Dougal Dixon, The Illustrated Encyclopedia of Dinosaurs, Londres, Lorenz Books, 2007 (ISBN 978-0-7548-1573-0), « Fast Hunters », p. 160–161
- P. Senter, « Comparison of Forelimb Function Between Deinonychus And Bambiraptor (Theropoda: Dromaeosauridae) », dans Journal of Vertebrate Paleontology, vol. 26, no 4, 2006, p. 897–906 [lien DOI]
- C. A. Forster, « The paleoecology of the ornithopod dinosaur Tenontosaurus tilletti from the Cloverly Formation, Big Horn Basin of Wyoming and Montana », dans The Mosasaur, vol. 2, 1984, p. 151–163
- M. J. Wedel, « Sauroposeidon: Oklahoma’s Native Giant », dans Oklahoma Geology Notes, vol. 65, no 2, 2005, p. 40–57 [texte intégral [PDF] (page consultée le 7 juillet 2007)]
- W. D. Maxwell, « Taphonomy and paleobiological implications of Tenontosaurus–Deinonychus associations », dans Journal of Vertebrate Paleontology, vol. 15, no 4, 1995, p. 707–712 [lien DOI] (abstract)
- B. T. Roach, « A reevaluation of cooperative pack hunting and gregariousness in Deinonychus antirrhopus and other nonavian theropod dinosaurs », dans Bulletin of the Peabody Museum of Natural History, vol. 48, no 1, 2007, p. 103–138 [lien DOI]
- Rihui Li, « Behavioral and faunal implications of Early Cretaceous deinonychosaur trackways from China », dans Naturwissenschaften, vol. 95, no 3, 2007, p. 185–91 [lien PMID, lien DOI]
- Therrien, F., Henderson, D.M. et Huff, C.B. (2005). "Bite me: biomechanical models of theropod mandibles and implications for feeding behavior." Pp. 179–237 in Carpenter, K. (ed.), The Carnivorous Dinosaurs. Indianapolis: Indiana University Press.
- P.M. Gignac, « A description of Deinonychus antirrhopus bite marks and estimates of bite force using tooth indentation simulations », dans Journal of Vertebrate Paleontology, vol. 30, no 4, 2010, p. 1169–1177 [lien DOI]
- Rihui Li, « Behavioral and faunal implications of deinonychosaur trackways from China », dans Naturwissenschaften, 2007 [lien DOI]
- Adams, Dawn (1987) "The bigger they are, the harder they fall: Implications of ischial curvature in ceratopsian dinosaurs" p. 1–6 in Currie, Philip J. et Koster, E. (eds) Fourth symposium on mesozoic terrestrial ecosystems. Tyrrell Museum, Drumheller, Canada
- K. Carpenter, « Evidence of predatory behavior by carnivorous dinosaurs », dans Gaia, vol. 15, 1998, p. 135–144
- Phil L. Manning, « Dinosaur killer claws or climbing crampons? », dans Biology Letters, vol. 2, 2006, p. 110–112 [lien DOI]
- (en) S. J. J. F. Davies, Ratites and Tinamous, New York, Oxford University Press, 2002 (ISBN 978-0-19-854996-3) (LCCN 2001133059)
- (en) E. T. Gilliard, Living birds of the world, Garden City, NY, Doubleday, 1958
- Kofron, Chhristopher P. (1999) "Attacks to humans and domestic animals by the southern cassowary (Casuarius casuarius johnsonii) in Queensland, Australia
- Christopher P. Kofron, « Case histories of attacks by the southern cassowary in Queensland », dans Memoirs of the Queensland Museum, vol. 49, no 1, 2003, p. 335–338
- (en) Kent H. Redford et Gustav Peters, « Notes on the biology and song of the red-legged seriema (Cariama cristata) », dans Journal of Field Ornithology, vol. 57, no 4, 1986, p. 261–269 [texte intégral]
- K. Carpenter, « Forelimb biomechanics of nonavian theropod dinosaurs in predation », dans Senckenbergiana Lethaea, vol. 82, 2002, p. 59–76 [lien DOI]
- Gauthier, J.; Padian, K. (1985). "Phylogenetic, Functional, And Aerodynamic Analyses Of The Origin Of Birds And Their Flight" in The Beginnings Of Birds. M.K. Hecht, J.H. Ostrom, G. Viohl, & P. Wellnhofer (eds) proceedings of the International Archaeopteryx Conference, Eichstätt, 1984: 185–197, Eischtatt: Freunde des Jura-Museums Eichstätt. isbn 3-9801178-0-4.
- (en) Gishlick, A.D., New Perspectives on the Origin and Early Evolution of Birds, New Haven, Yale Peabody Museum, 2001, « The function of the manus and forelimb of Deinonychus antirrhopus and its importance for the origin of avian flight », p. 301–318
- W. Parsons, « Morphology and size of an adult specimen of Deinonychus antirrhopus, (Saurischia, Theropoda) », dans Journal of Vertebrate Paleontology, vol. 26, no 3 sup., 2006, p. 109A
- W. L. Parsons, « Further descriptions of the osteology of Deinonychus antirrhopus (Saurischia, Theropoda) », dans Bulletin of the Buffalo Society of Natural Sciences, vol. 38, 2009, p. 43–54 [texte intégral]
- R. Kool, « The walking speed of dinosaurs from the Peace River Canyon, British Columbia, Canada », dans Canadian Journal of Earth Science, vol. 18, 1981, p. 823–825
 Portail de la paléontologie
Portail de la paléontologie Portail de l’origine et de l’évolution du vivant
Portail de l’origine et de l’évolution du vivant Portail des dinosaures
Portail des dinosaures Portail de la zoologie
Portail de la zoologie
Catégories :- Coelurosaure
- Dinosaure du Crétacé
-











Wikimedia Foundation. 2010.