- Virus de la dengue
-
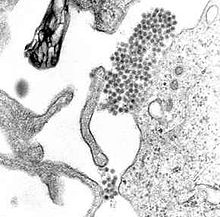
Le virus de la dengue (DENV) fait partie de la famille des Flavivirus et il est classé en quatre sérotypes différents (DENV-1 à DENV-4). Ce virus est l’un des plus répandus à travers le monde et on dénote environ 100 millions de cas chaque année de sa maladie sous sa forme bénigne. Cette grande incidence de cas fait en sorte que les recherches pour mieux comprendre le cycle viral et les moyens de réplication de ce virus sont très importantes afin de créer un vaccin.
Sommaire
Généralités
Le virus de la dengue est la cause de deux maladies qui diffèrent par leur gravité. Il y a la forme bénigne qui est apparue il y a environ 200 ans et qui cause une fièvre bénigne. Il y a également une forme plus grave de la maladie qui cause une fièvre hémorragique et qui a été identifiée après la deuxième Guerre mondiale. Le virus de la dengue est retrouvé en Asie et en Amérique du Sud et est transmis par des moustiques de la famille des Aedes. Ce moustique est répandu en forêt et dans les endroits humides, car ces endroits sont favorables à sa reproduction. Le virus de la dengue est responsable de 100 millions de cas de fièvre et de 500 000 fièvres hémorragiques chaque année. De ce nombre, environ 25 000 personnes en meurent.
Types de virus
Le virus de la dengue fait partie de la famille des Flavivirus et peut être divisé en quatre sérotypes différents DENV-1, DENV-2, DENV-3 et DENV-4. Ces quatre sérotypes ont des différences au niveau de la composition de leur génome et de leurs antigènes de surface. Une personne infectée par un de ces virus est immunisée à vie contre celui-ci mais n’est pas protégée contre les trois autres sérotypes. Des recherches récentes ont démontré que chaque sérotype peut être divisé en génotypes selon l’endroit où le virus est retrouvé dans le monde et que chaque génotype pourrait être subdivisé en intra-génotype selon des différences dans la composition du génome.
Génome du virus
Le génome du virus de la dengue est sous la forme d’un ARN+ simple brin d’environ 11 kb. Il est composé d’un seul cadre de lecture et contient une région non codée à chaque extrémité : la 5' UTR d’environ 100 nucléotides et la 3' UTR d’environ 400 nucléotides. L’ARN code pour une seule polyprotéine qui est clivée pendant et après sa traduction en 7 protéines non structurales (NS1, NS2A, NS2B, NS3, NS4A, NS4B et NS5) et en trois protéines structurales (C, prM et E). La protéine C est présente dans la capside, la protéine prM est un précurseur de la protéine M qui est retrouvée dans la membrane et la protéine E est la composante principale de l’enveloppe. Comme les autres virus à ARN, le génome du virus de la dengue est en constante variation à cause de l'ARN polymérase qui n’a pas de mécanisme de vérification des erreurs lors de la réplication de l’ARN. Les nombreuses variations génétiques sont aussi causées par la réplication rapide du virus et la grande quantité de virus retrouvée dans le monde.
Le cycle viral
L'entrée du virus
Le virus de la dengue est un virus enveloppé. Il pénètre donc dans les cellules cibles par fusion. Le processus de fusion est contrôlé par la protéine E qui est la protéine principale de l’enveloppe. Elle a un rôle important dans les propriétés phénotypiques et immunogéniques du virus. La protéine E a plusieurs fonctions, dont celle de reconnaître les cellules cibles qui contiennent de la clathrine à leur surface. La protéine E contient un peptide de fusion qui est hydrophobe et qui vient s’insérer dans la bicouche lipidique de la cellule cible. Une suite de réarrangements de la protéine E s’ensuivent, dont sa transformation de dimère à trimère. Ces changements permettent la fusion de l’enveloppe virale avec la bicouche cellulaire. Suite à sa fusion, le virus se trouve dans un endosome à l'intérieur de la cellule hôte. Lorsque le pH de l'endosome diminue, le virus peut sortir de celui-ci et commencer sa réplication dans le cytoplasme.
Phase intracellulaire
Après son entrée dans la cellule cible, l’ARN viral est traduit en une polyprotéine par les ribosomes de la cellule hôte. La polyprotéine est clivée en ses composants mentionnés plus haut, pendant et après sa traduction au niveau du réticulum endoplasmique rugueux par une signalase cellulaire et une protéase virale. La protéine structurale C a un rôle important dans la translocation des protéines virales vers le réticulum endoplasmique rugueux. En effet, la protéine de la capside (C) est reliée au précurseur de la protéine M (prM) par un lien hydrophobe qui sert de signal aux protéines virales afin de les diriger vers la membrane du réticulum endoplasmique pour y être internalisé dans sa lumière. Après l’entrée de C-prM dans la lumière du réticulum endoplasmique, le lien hydrophobe est clivé par la protéase virale NS3/2B qui est retrouvée dans le cytoplasme du réticulum endoplasmique. La protéine C, qui est composée de quatre hélices α, devient mature suite à ce clivage. Trois des hélices α se positionnent autour du cœur viral et la quatrième, grâce à des résidus basiques, interagit avec l’ARN viral. Suite à cet assemblage capside-ARN, il se produit une association entre prM et E pour former un hétérodimère. Par la suite, la capside (C) , l’ARN, l’enveloppe (E) et le précurseur de la protéine membranaire (prM) s’assemblent en un complexe pour former les virions.
Sortie du virus
Lors de la sortie du virion, le précurseur prM est clivé par une furine protéase au niveau du trans-Golgi. Le précurseur prM forme donc la protéine M, qui s’insère dans l’enveloppe. Le fragment pr, lui, reste en solution et il est excrété lors de la sortie du virus de la cellule hôte. Après ce clivage, on dit du virion qu’il est mature. Le virion se dirige près de la bicouche lipidique de la cellule hôte et sort par bourgeonnement afin d’utiliser cette bicouche comme enveloppe. Le cycle se répète ensuite.
Vaccin
Le virus de la dengue est le plus meurtrier de tous les virus transmis par des moustiques à travers le monde. Il n’existe pas encore de vaccin contre la maladie. Cependant, des recherches sont présentement en cours pour essayer de trouver une stratégie qui pourrait éliminer le virus. Par exemple, une recherche récente a permis de démontrer qu’en inhibant la formation des vésicules lipidiques, que le virus de la dengue utilise pour sortir de l’appareil de Golgi, on diminue fortement la réplication de DENV. D’autres recherches tentent de modifier la protéine d’enveloppe E afin d’empêcher l’entrée du virus dans la cellule hôte.
Notes et références
Annexes
Articles connexes
Liens externes
Bibliographie
- Marcello M. Samsa et al., 2009, Dengue Virus Capsid Protein Usurps Lipid Droplets for Viral Paricle Formation, PLoS Pathog. 2009 october
- Alberto A Amarilla et al., 2009, Genetic diversity of the E Protein of Dengue Type 3 Virus, Journal of General Virology. 2009;6:113
- Aaron G. Schmidt et al., 2010, Peptide Inhibitors of Dengue-Virus Entry Target a Late-Stage Fusion Intermediate, PLoS Pathog. 2010 April
- Osmali Osman et al., 2009, Genetic characterization of dengue virus type 1 isolated in Brunei in 2005-2006, Journal of General Virology 90. 2009
- Pei-Gang Wang et al., 2009, Efficient Assembly and Secretion of Recombinant Subviral Particles of the Four Dengue Serotypes, PLoS Pathog. 2009 December
Wikimedia Foundation. 2010.