- Concept de continuum fluvial
-
Continuum fluvial
Le concept de continuum fluvial (River Continuum Concept), introduit dans les années 1980 par Robin L. Vannote, est un modèle descriptif longitudinal du fonctionnement écologique des cours d’eau des zones tempérées, qui intègre à la fois les indicateurs biologiques et la géomorphologie.
Sommaire
Contexte
Le concept de continuum fluvial repose sur l’idée que les cours d’eau doivent être considérés comme des écosystèmes ouverts, en interaction avec leur ripisylves, qui se transforment continûment de leur source à leur embouchure. Les modifications progressives de leurs conditions physiques (largeur, profondeur, débit, courant, température...) de l’amont à l’aval engendrent un gradient continu des ressources trophiques disponibles qui détermine la distribution des invertébrés, en fonction des différentes stratégies développées par les espèces pour exploiter ces ressources alimentaires.
Division fonctionnelle des cours d’eau

Suivant le concept de continuum fluvial, on peut toujours diviser les systèmes lotiques en trois domaines majeurs : le ruisseau ou torrent puis les eaux de moyenne vallée et enfin les grands fleuves et rivières.
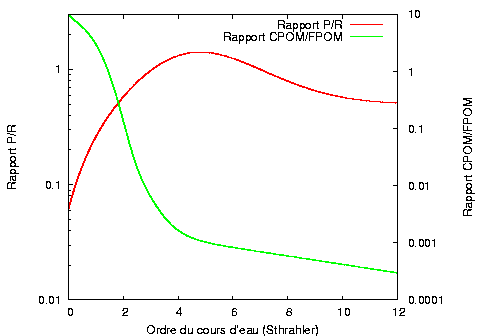
Sur le domaine amont, le cours d’eau est étroit et sa ripisylve est généralement très développée. La faible luminosité et les forts taux de dilution des éléments inorganiques inhibent la production de matière organique par photosynthèse (production primaire) mais de grandes quantités de matière organique allochtone tombent dans le ruisseau. Par conséquent, la respiration (transformation de la matière organique en éléments minéraux) est plus importante que la production primaire (rapport P/R < 1, hétérotrophie). C’est dans ce domaine que la diversité des ressources trophiques disponibles est la plus grande.
 Un fleuve : l’Isar au niveau de München-Thalkirchen
Un fleuve : l’Isar au niveau de München-Thalkirchen
Sur le domaine intermédiaire, l’élargissement de la rivière et la diminution des taux de dilution permet à la production de matériel organique autochtone, notamment sous formes d’algues, d’augmenter sensiblement. On a alors un rapport P/R > 1 (autotrophie).
Sur le domaine aval, le matériel organique particulaire est abondant. L’approfondissement du lit et la turbidité limitent la luminosité et la photosynthèse n’a plus lieu qu’à la surface des eaux. Il y a par conséquent retour à l’hétérotrophie.
Distribution des populations invertébrées
La composition de la biocénose et, plus particulièrement des populations invertébrées, dépend de la position de son biotope le long du continuum fluvial, que l’on mesure par l’ordre du cours d’eau dans la classification de Sthraler.
On classe les invertébrés en quatre grandes catégories : les broyeurs, les collecteurs, les brouteurs et les prédateurs. A l’exception des prédateurs, tous ces organismes se nourrissent de matière organique d’origine végétale : ce sont des saprophytes.
 Larves de Simulie (collecteur)
Larves de Simulie (collecteur)Les broyeurs ont pour rôle de déchiqueter le matériel organique grossier (CPOM = Coarse Particulate Organic Matter), de taille millimétrique, tel que des lambeaux de feuillage. Les amphipodes, les aselles, les larves d’éphéméroptères et de plécoptères comptent parmi les broyeurs les plus courants de cours d’eau d’Europe Centrale.
Les collecteurs sont, eux, capables de récolter la matière organique en suspension dans l’eau courante ou piégée dans les sédiments. Les particules dont ils se nourrissent ont une taille comprise entre 0,5 et 50 micrométres (UPOM = Ultrafine Particulate Organic Matter et FPOM = Fine Particulate Organic Matter). Les larves de simulie, les nématodes et de nombreuses larves de chironomidés appartiennent par exemple au groupe des collecteurs.
 Larve d’Aeschne bleue (prédateur)
Larve d’Aeschne bleue (prédateur)Les brouteurs se nourrissent de ce qui se développe (algues, micro-organismes…) sur les pierres, les morceaux de bois mort et les grandes plantes aquatiques qui structurent le lit des cours d’eau. Parmi eux, on dénombre les gastéropodes, les larves de liponeura et certaines de chironomidés.
Quant aux prédateurs, comme les larves de libellules et différents coléoptères, ils se nourrissent des autres animaux.
Du fait de l’évolution du matériel organique disponible, la représentation de ces différents groupes varient de l’amont vers l’aval du cours d’eau. En amont, c’est-à-dire dans la partie torrentielle, où le matériel organique grossier est abondant, on trouve une forte proportion de broyeurs mais aussi de collecteurs et les brouteurs sont peu représentés. La proportion de brouteurs augmente ensuite, ainsi que celle des collecteurs, au détriment des broyeurs dont les populations régressent significativement, pour finalement disparaître complètement. En aval, où subsiste presque uniquement le matériel organique particulaire, les collecteurs constituent le groupe dominant. Quant aux prédateurs, leur proportion reste toujours à peu près constante ; seule varie la représentation des espèces au sein du groupe, qui suit l’évolution des proies disponibles. Des variations atypiques dans la répartitions des différents groupes invertébrés le long d’un cours d’eau sont le signe d’une perturbation du milieu naturel.
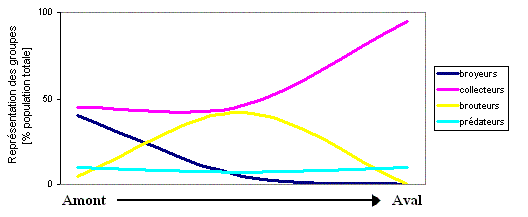 Distributions des groupes d’invertébrés de l’amont vers l’aval d’un cours d’eau (d’après Vannote et al.)
Distributions des groupes d’invertébrés de l’amont vers l’aval d’un cours d’eau (d’après Vannote et al.) Relation du rapport CPOM/FPOM au rapport P/R le long du continuum fluvial
Relation du rapport CPOM/FPOM au rapport P/R le long du continuum fluvialStabilité du système lotique
Le modèle du continuum fluvial permet de mieux comprendre l’allocation des ressources énergétique dans les systèmes lotiques.
A chaque niveau du cours d’eau, l’énergie disponible sous forme de matériel organique peut être consommée, stockée ou transférée en aval. Comme elle constitue le facteur limitant du système, celui-ci doit assurer son utilisation optimale. Des ressources inutilisées permettent en effet à de nouvelles espèces d’intégrer l’écosystème, jusqu’à saturation de son potentiel énergétique. Ce principe est valable pour tout écosystème de manière générale mais il joue ici un rôle particulier en raison de l’évacuation continue du matériel organique non consommé, emporté par les eaux courantes, qui implique un impératif d’efficacité plus important et surtout une utilisation continue des ressources.
Le problème de la continuité temporelle se pose essentiellement aux échelles de la journée et de l’année. Au cours d’une journée, les risques accrus de prédation (les poissons chassent essentiellement grâce à leur vision) et les modifications de facteurs abiotiques tels que la température et la luminosité, induisent une alternance des espèces actives. Les variations journalières étant plus importante sur les cours d’eau intermédiaires, c’est là que la diversité biologique est la plus importante.
L’optimisation constante de l’utilisation des ressources énergétique dans les systèmes lotiques garantit une grande stabilité vis à vis des fluctuations et perturbations de leurs conditions physiques. Après une perturbation, ces systèmes trouvent très rapidement un nouvel équilibre sans succession écologique.
Développement et application du modèle
Le concept de continuum fluvial est le fruit de plusieurs années de recherches financées par la fondation Rockefeller et publiées en 1980 sous le titre „The River Continuum Concept“ dans le Canadian Journal of Fisheries and Aquatic Sciences. Le modèle s’appuyait sur les travaux de Ruth Patrick et de Luna Leopold. L’apport essentiel de l’équipe de Vannote aura été une prise en compte globale et exhaustive de l’écosystème lotique.
Après leur publication, les travaux de Vannote ont très vite emporté une large approbation de la communauté scientifique et le concept de continuum fluvial devint le modèle de référence pour la description de la biocénose des écosystèmes lotiques. Il a notamment le mérite d’expliquer correctement certaines découvertes antérieures, comme celles de Robert Lauterborn (1918) et de Joachim Illies (1961).
Actuellement, on a surtout recours au modèle de Vannote pour diagnostiquer les perturbations des cours d’eau en comparant la composition réelle de sa biocénose à la composition théorique dans le modèle du continuum fluvial.
Limites du concept et propositions alternatives

Malgré ses succès, le concept de continuum fluvial, tel que développé aujourd'hui a un champ d’application limité :
- il a été développé pour les zones biogéographiques tempérées à moyennement froides
- il ne permet pas de décrire les portions stagnantes des cours d’eau (lacs, barrages, bras morts…)
- il ne tient pas compte des phénomènes de crue
- il suppose que la végétation rivulaire est intacte
- il ne permet pas de prédire l’impact de la pollution des eaux
- il ne tient pas compte de l'impact des affluents
Pour tenter d’apporter des réponses à ces problèmes, le concept a été retravaillé par différents chercheurs. J.V. Ward et J.A. Stanford ont par exemple élaboré le Serial Discontinuity Concept (1983) pour tenir compte des ralentissements des eaux courantes puis l’Hyporheic Corridor Concept (1993) pour intégrer les dimensions latérales et verticales (rives et profondeur) à la problématique. Le Flood Pulse Concept de W.J. Junk et al. (1989), modifié par P.B. Bayley (1990) et K. Tockner et al. (2000) vint encore améliorer la prise en compte des phénomène d’inondation. Le cas des très grands fleuves est abordé par le Riverine Productivery Model de J.H. Thorp et M.D. Delong (1994). Le Telescoping Ecosystem Model de Fisher et al. (1998) permet, quant à lui, d’expliquer les flux de matière entre les eaux de surface du cours d’eau proprement dit, les eaux de sub-surface (zones hyporhéique et parafluviale) et la zone riveraine.
Par ailleurs cette notion est complétée par la dynamique fluviale, qui est l'approche physique de l'équilibre et de l'évolution d'un cours d'eau.
Bibliographie
River Continuum Concept
- Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE (1980): The River Continuum Concept, Canadian Journal of Fisheries and Aquatic Sciences 37(1): 130-137. Texte intégral
- Wasson G. (1989): Eléments pour une typologie fonctionnelle des eaux courantes. Revue critique de quelques approches existantes, Bulletin d'écologie 20(2): 109-127.
- Maamri A., Chergui H., Pattee E. (1995): Impact des apports végétaux allochtones sur la distribution de la faune benthique dans un cours d'eau montagnard marocain, l'oued zegzel, Ecologie 26(2): 81-94.
Autres approches
- Junk JW, Bayley PB, Sparks RE (1989): The flood pulse concept in river floodplain systems. Canadian Special Publications of Fisheries and Aquatic Sciences 106: 110-127
- Ward JV, Stanford JA (1983): The serial discontinuity concept of river ecosystems. In: Fontaine TD, Bartell SM (Hrsg): Dynamics of lotic ecosystems, 29-42, Ann Arbor Science Publications, Ann Arbor, Michigan.
- Stanford JA, Ward JV (1993): An ecosystem perpective of alluvial rivers: connectivity and the hyporheic corridor, Journal of the North American Benthological Society 12, 48-60
- Bayley PB (1990): The flood pulse advantage and the restoration of river -floodplain-systems, Regulated Rivers: Research & Management 6, 75-86
- Tockner K, Malard F, Ward JV (2000): An extension of the flood pulse concept, Hydrological Processes 14, 2861-2883
- Thorp JH, Delong MD (1994): The riverine productivity model: A heuristic view of carbon sources and organic processing in large river systems, Oikos 70, 305-308
- Fisher, S. G., Grimm, N. B., Marti, E., Holmes, R. M. & Jones, J. B. (1998): Material spiraling in stream corridors : a telescoping ecosystem model, Ecosystems, 1 : 19-34.
- Bravard et Petit (2000) : "Les Cours d'Eau, Dynamique du Système Fluvial", PUF.
- Amorros et Petts (1994) : "Hydrosystèmes Fluviaux"
 Portail de l’eau
Portail de l’eau
Catégories : Géomorphologie fluviale et lacustre | Écosystème | Cours d'eau




Wikimedia Foundation. 2010.