- Choix du partenaire sexuel
-
Sélection intersexuelle
 La sélection intersexuelle est particulièrement visible au niveau des aires de parade. Chez le Tétras lyre, les mâles se regroupent et paradent dans des tourbières. Les femelles viennent les observer et s'accouplent parfois avec l'un d'entre eux.
La sélection intersexuelle est particulièrement visible au niveau des aires de parade. Chez le Tétras lyre, les mâles se regroupent et paradent dans des tourbières. Les femelles viennent les observer et s'accouplent parfois avec l'un d'entre eux.
La sélection intersexuelle désigne, en biologie de l'évolution, la pression sélective exercée sur les individus d'une espèce sexuée par le choix, fait par les individus d'un des deux sexes (généralement le sexe femelle[1]), du ou des individu(s) de l'autre sexe qui lui donneront des descendants. Cette forme de sélection sexuelle s'oppose à la sélection intrasexuelle, bien que ces deux mécanismes évolutifs interagissent fortement.
Même si le sexe choisissant l'autre est généralement le sexe femelle, il arrive que le mâle choisisse les femelles avec lesquelles il se reproduit, lesquelles sont alors sujettes à la sélection intersexuelle. Chez certaines espèces, les deux sexes sélectionnent leurs partenaires pour la reproduction. De nombreuses théories ont été développées pour expliquer le fait que ce soit l'un ou l'autre des deux sexes qui choisit ses partenaires. Par commodité, dans cet article, le sexe choisissant ses partenaires d'accouplement (et éventuellement ses descendants) sera désigné, par la suite, comme le sexe femelle, bien que ce ne soit pas toujours le cas.
On a longtemps considéré que la sélection intersexuelle se limitait au choix du partenaire (ou mate choice en anglais) pour la reproduction. On sait maintenant que, chez certaines espèces, les femelles s'accouplent plusieurs fois et opèrent un choix sur les gamètes mâles utilisés pour féconder les ovules, ce qui favorise une compétition spermatique. Il arrive aussi que la femelle s'occupe préférentiellement de certains de ses descendants. On distingue donc le choix du partenaire (ou choix pré-copulatoire) du choix post-copulatoire, et il convient de parler de choix de la paternité (ou de la maternité), plutôt que du partenaire sexuel[2].
Historique du concept de sélection intersexuelle
C'est Darwin qui crée, dans son livre L'Origine des espèces publié en 1859, le concept de sélection sexuelle. Il détaille par la suite cette notion dans son livre La Filiation de l'homme et la sélection liée au sexe publié en 1971. C'est alors qu'il fait la distinction entre ce que l'on appellera par la suite sélection intersexuelle et sélection intrasexuelle. Si cette dernière est unanimement reconnue par les contemporains de Darwin, l'existence de la sélection intersexuelle est assez controversée lors de la publication de L'Origine des espèces, et met du temps à être reconnue[3]. L'étude de la sélection intersexuelle est ainsi longtemps négligée, et l'on n'accorde aux femelles qu'un rôle passif, limité à la récompense du gagnant de la compétition intrasexuelle.
Cependant, Darwin n'explique pas l'origine du choix des femelles, et de la préférence pour certains mâles, même s'il rajoute un paragraphe sur l'évolution des préférences dans la seconde édition de The Descent of Man, and Selection in Relation to Sex (1874, II, p495)[réf. nécessaire][4]. Cette absence d'explication pousse Thomas Hunt Morgan à tourner en dérision la théorie de sélection intersexuelle de Darwin[5], proposant sur le ton de l'ironie un mécanisme de l'évolution des préférences, dans lequel le caractère préféré et la préférence de ce caractère, tous deux apparus par hasard, coévolueraient et donneraient lieu à un emballement. Pourtant, si Morgan considère ce mécanisme comme ridicule, c'est très sérieusement que Ronald Aylmer Fisher développe en 1915 la théorie du fils sexy[6] (ou emballement fisherien, mécanisme aujourd'hui largement accepté[réf. nécessaire]), dont le principe est celui proposé sarcastiquement par Morgan. On ne sait cependant pas si Fisher s'est inspiré de la remarque critique faite par Morgan pour développer sa théorie: il connaît certainement le livre de Morgan au moment où il développe sa théorie, puisqu'il le cite dans un de ses articles publié peu avant[7], mais il ne fait pas référence à Morgan dans son article sur l'évolution des préférences sexuelles[4].
L'étude de la sélection naturelle[3], et celle de la sélection intersexuelle en particulier[réf. nécessaire], est cependant relativement délaissée jusqu'aux années 1970. Ainsi, bien que Alfred Russel Wallace[8] et Fisher[6] évoquent déjà, en 1889 et 1915, des idées s'apparentant à la théorie des bons gènes[4], il faut attendre la fin des années 1960 avant que d'autres mécanismes de choix du partenaire sexuel ne soient réellement théorisés et testés.
De même, le rôle relatif des deux sexes lors de la sélection sexuelle n'est presque pas étudié jusqu'aux années 1970. Robert Trivers crée en 1972[9] un cadre conceptuel de base pour cette étude, développant la théorie de l'investissement parental. Celle-ci propose que le sexe pouvant se permettre de choisir l'autre est celui qui investit le plus dans la reproduction. Cette théorie est développée pendant les années suivantes et jusqu'à aujourd'hui, en introduisant de nouveaux facteurs pour expliquer l'existence d'un sexe choisissant, tels qu'un biais de sexe-ratio opérationnel[réf. nécessaire] ou des différences de taux de reproduction potentiel entre mâles et femelles[réf. nécessaire].
Plus récemment, l'idée que la sélection intersexuelle ne se limite pas à un choix du partenaire de la reproduction, mais peut continuer à s'exercer après l'accouplement, voire après la naissance, se développe. La notion de choix de la paternité vient remplacer celle de choix du partenaire sexuel, et des théories de sélection intersexuelle post-copulatoire sont construite par prolongement des théories de sélection pré-copulatoire développées jusqu'alors[2]. Des données empiriques récentes commencent aujourd'hui à appuyer ces théories[réf. nécessaire].
Quel sexe choisit l'autre ?
Diversité au sein du vivant
Chez la plupart des espèces décrites, le choix de la paternité des enfants, s'il existe, est fait par la femelle. Cependant, ce n'est pas nécessairement le cas, et l'on connaît plusieurs espèces pour lesquelles c'est le mâle qui choisit les femelles avec lesquelles il s'accouple. De telles espèces, chez lesquelles la compétition pour la reproduction est plus forte chez les mâles que chez les femelles, sont qualifiées d'espèces à rôle sexuel inversées[10]. On observe souvent chez ces femelles des caractéristiques usuellement mâles, telles que la présence d'ornements, et chez les mâles des caractéristiques usuellement femelles telles que la garde et le soin des enfants. L'exemple le plus connu d'espèces à rôles sexuels inversés est celui des hippocampes, mais il en existe de nombreux autres.
De manière plus générale, on observe une grande diversité des systèmes de choix de paternité au sein des espèces sexuées. Ainsi, il arrive que les deux sexes choisissent (et donc soient sélectionnés), ou inversement que le choix soit très limité chez les deux sexes. Par ailleurs, le fait que ce soit l'un ou l'autre (ou les deux) des sexes qui choisisse peut changer en fonction des conditions environnementales, ainsi que l'exigence de ces choix[11]. Enfin, le sexe choisissant peut même varier en fonction des individus qui composent l'espèce[réf. nécessaire].
Paramètres déterminant le sexe choisissant
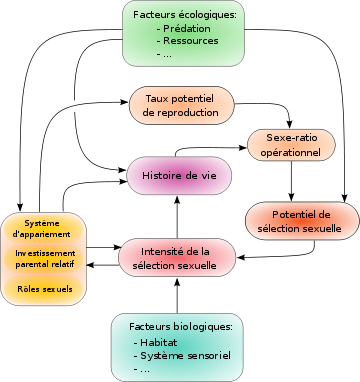 Relations entre la sélection sexuelle et les variables reliées à celle-ci, proposées par Malte Andersson[12]. L'investissement parental relatif influe sur le taux potentiel de reproduction, qui modifie le sexe ratio opérationnel et donc la force et la direction de la sélection sexuelle, modifiant éventuellement les rôles sexuels.
Relations entre la sélection sexuelle et les variables reliées à celle-ci, proposées par Malte Andersson[12]. L'investissement parental relatif influe sur le taux potentiel de reproduction, qui modifie le sexe ratio opérationnel et donc la force et la direction de la sélection sexuelle, modifiant éventuellement les rôles sexuels.L'existence d'une sélection sexuelle vient du fait que l'accès à l'un des deux sexes pour la reproduction est une ressource limitante pour l'autre sexe. Dans ce cas, une partie des individus du sexe ainsi limité n'aura pas accès à la reproduction, ce qui génère une sélection, engendrant une compétition entre les individus du sexe limité, et permettant aux membres du sexe limitant d'effectuer un choix de la paternité de ses descendants. Ainsi l'intensité de la sélection naturelle dépend directement du sexe-ratio opérationnel, lequel dépend lui-mêmes d'autres facteurs tels que le taux potentiel de reproduction et l'investissement parental de chacun des deux sexes.
Le sexe-ratio opérationnel
Le sexe-ratio opérationnel est défini comme le rapport du nombre de femelles fécondables sur le nombre de mâles sexuellement actifs à un moment donné[13]. En outre, il ne prend en compte que les individus fertiles. De plus, seuls les individus qui, à un moment donné, sont disponibles pour se reproduire sont pris en compte. Par exemple, chez les mammifères les individus en gestation à un moment donné ne sont pas comptés dans le sexe-ratio car incapables de participer à la reproduction. Le sexe-ratio opérationnel décrit donc le nombre de mâles par femelle réellement rencontré lors de la reproduction, et permet donc d'évaluer au mieux l'intensité de la sélection sexuelle exercée sur chaque sexe, et ainsi de déterminer quel sexe sera le plus sujet à la compétition.
Ainsi, si le sexe-ratio opérationnel est biaisé en direction des mâles, un plus grand nombre de mâles que de femelles est impliqué dans la reproduction, conduisant à une compétition entre les mâles et un choix des femelles pour certains mâles. En revanche, un sexe-ratio opérationnel biaisé vers les femelles favorise l'évolution de rôles sexuels inversés.
Le sexe-ratio opérationnel est en pratique très difficile à mesurer. Il dépend cependant de certains paramètres que l'on peut plus facilement estimer. Il est en effet directement relié au sexe-ratio adulte, éventuellement biaisé par une mortalité plus importante chez l'un des deux sexes. Il est de plus fortement dépendant des taux de reproduction potentiels des mâles et des femelles.
Le taux potentiel de reproduction
Le taux de reproduction potentiel d'un individu est le nombre maximum de descendants indépendants que celui-ci peut produire par unité de temps[14]. Le sexe produisant plus vite ses descendants sera ainsi sur-représenté lors de la reproduction, et le sex-ratio opérationnel sera biaisé vers celui-ci. Ce taux n'est qu'un potentiel: en effet, chaque descendant ayant obligatoirement un père et une mère (condition de Fisher), le taux de reproduction moyen réel est nécessairement le même pour les mâles et les femelles. Cependant les individus d'un sexe transmettant des descendants ont en moyenne un taux de reproduction réel supérieur à la moyenne de son sexe (certains ne se reproduisant pas), plus proche du taux de reproduction potentiel.
Ainsi, le sexe pour lequel le taux de reproduction potentiel est le plus fort a tendance à être le plus sujet à la compétition pour la reproduction, et la pression sexuelle exercée sur celui-ci est généralement plus forte. Le taux de reproduction potentiel dépend de nombreux facteurs. Les mâles produisant, par définition, des gamètes plus petits, ils peuvent en produire plus que les femelles (à coût égal), ce qui augmente leur taux de reproduction potentiel. De même, le temps passé à s'occuper de chaque portée ne peut parfois pas être employé à la génération de nouveaux descendants, ce qui diminue le taux de reproduction potentiel. Le taux de reproduction potentiel dépend ainsi, entre autres, de l'anisogamie et de l'investissement parental.
Le taux de reproduction potentiel à l'avantage d'être plus facile à mesurer que le sexe-ratio opérationnel, et possède un meilleur pouvoir de prédiction des rôles sexuels que l'investissement parental[14]. Clutton-Brock et Parker montrent que sur 13 espèces dont le le taux de reproduction maximum est plus grand chez les mâles que chez les femelles, toutes montrent une compétition plus grande chez les mâles. Réciproquement, 14 des 16 espèces chez lesquelles les femelles ont un taux de reproduction maximum plus élevé présentent une compétition plus intense chez les mâles.[14]
L'investissement parental
L'investissement parental a été le premier facteur utilisé pour expliquer la répartition des rôles sexuels au sein du vivant. Ce concept, développé par Trivers en 1972[9] considère que le sexe qui investit le plus dans sa descendance devient une ressource limitante pour l'autre sexe, exerçant ainsi une pression de sélection sexuelle sur ce dernier.
Trivers défini lui-même l'investissement parental comme l'ensemble des investissement qu'un parent fait pour un de ses descendants qui augmente la chance de survie de ce descendant et réduit la possibilité du parent d'investir dans d'autres descendants[15]. L'investissement parental comprend ainsi de nombreuses formes d'investissement parental, comme la production des gamètes, ou encore la garde et l'élevage des petits.
La théorie de l'investissement parental prédit ainsi que si la femelle investit plus que le mâle, les rôles sexuels seront conventionnels (les mâles se battent pour l'accouplement et les femelles choisissent), alors qu'ils seront inversés si les mâles investissent moins. Du fait de l'anisogamie, les femelles investissent au départ plus que les mâles, ce qui explique (selon cette théorie) que les rôles sexuels soient plus souvent conventionnels. De plus, Trivers explique qu'il existe des mécanismes favorisant le maintien d'un investissement parental supérieur chez les femelles, et donc le maintien des rôles sexuels conventionnels. Cette théorie permet enfin d'expliquer un certain nombre de cas de réversion des rôles sexuels, pour des espèces chez lesquels les mâles investissent plus que les femelles.
Cependant, la théorie de Trivers ne permet pas d'expliquer la distribution des rôles sexuels chez toutes espèces présentant des soins parentaux. Chez l'Épinoche ou le Nandou d'Amérique, par exemple, les soins sont exclusivement paternels, mais sont aussi les plus sujets à la compétition intrasexuelle. Même si ceci pourrait être dû à un investissement femelle supérieur et moins visible que celui des mâles (comme la production des gamètes), il y a peu d'arguments en faveur de cette explication[14].
L'investissement parental agit en réalité de manière indirecte sur l'intensité de la sélection sexuelle, via le taux potentiel de reproduction. En effet, d'après la définition de Trivers, l'investissement parental chez un sexe diminue le temps disponible pour former de nouveaux descendants, et diminue donc le taux potentiel de reproduction de ce sexe favorisant ainsi un biais de sex-ratio opérationnel vers l'autre sexe et la compétition intrasexuelle chez ce dernier. L'investissement parental d'un sexe favorise donc bien la compétition intrasexuelle chez l'autre sexe, mais d'autres paramètres interviennent dans le déterminisme des rôles sexuels.
Modèles de prédiction du sexe choisissant
Bénéfices et coûts du choix de la paternité
Chez de nombreuses espèces sexuées, les femelles exercent un choix sur les mâles qui porteront la paternité de leur descendants, exerçant ainsi une pression de sélection intersexuelle sur ces mâles. Ces choix, qui détermineront les caractères sélectionnés chez les mâles, peuvent eux-mêmes avoir été sélectionnés parce qu'ils augmentent le fitness des femelles, ou peuvent être apparus par hasard sans apporter d'avantage sélectif aux femelles. Par ailleurs, le choix du ou des mâles qui seront les pères de leurs descendants a souvent un coût pour les femelles, généralement inférieur ou égal à l'avantage sélectif conféré par ce choix.
Avantage sélectif du choix
Le fait pour une femelle de choisir les mâles qui lui donneront des descendants, selon certains critères particuliers, peut avoir été sélectionné au cours de l'évolution, car ce choix confère un avantage sélectif aux femelles, celles choisissant leurs mâles ayant plus de descendants viables et fertiles. Les avantages sélectifs que peuvent conférer un choix sont divers et nombreux. Ainsi, le bénéfice obtenu par la femelle peut être direct, et se présenter sous forme de nourriture apportée à la femelle, ou encore de soins parentaux conférés par le mâle. Les bénéfices peuvent aussi être indirects, sous la forme de gènes capables d'augmenter le fitness de la femelle parce qu'ils produiront des descendants plus viables (parce qu'ils seront plus attrayants pour les autres femelles ou plus aptes à survivre par exemple).
Bénéfices directs
Le fait de choisir les mâles peut conférer un bénéfice direct aux femelles si les critères de ce choix reflètent la capacité du mâle à fournir divers avantages matériels. Ceux-ci peuvent-être des dons nutritifs, comme des proies chassées, des nutriments synthétisés par le mâle, voir le mâle lui-même qui s'offre à être mangé si le bénéfice de ce sacrifice est suffisamment grand pour le mâle. Les femelles peuvent aussi avoir avantage à choisir le mâle selon des critères permettant d'évaluer sa capacité à fournir des soins aux enfants, sous forme de nutrition ou de protection par exemple. Les mâles sont parfois choisis parce qu'ils sont plus aptes à défendre les femelles, ou parce qu'ils gardent des territoires plus avantageux (plus riches en ressources par exemple). Le choix du mâle sur des critères reflétant la fécondité du mâle peut aussi garantir à la femelle que ses ovules seront bien fécondés. Enfin, les signaux mâles peuvent garantir la bonne santé du mâle et éviter à la femelle la contamination par des parasites lors de la fécondation.
Avantages nutritifs
Soins parentaux des descendants
Défense d'un territoire
Fécondité du mâle
Risques de contamination et de prédation
Bénéfices indirects
Le bénéfice conféré par le choix du mâle peut aussi être indirect, s'il n'affecte non pas directement le fitness de la femelle, mais celui de ses descendants. Les avantages sont alors transmis sous forme génique par le mâle à ses descendants, et le fitness de la femelle est augmenté indirectement, par le biais de ses descendants. Les avantages génétiques conférés aux descendants peuvent être de plusieurs types. Si la préférence maternelle est déterminée génétiquement, la préférence pour un trait et le trait préféré vont être transmis ensemble à la génération suivante, donnant lieu à un phénomène d'auto-amplification appelé runaway de Fisher[16]. Choisir un mâle considéré comme plus sexy garantit donc aux descendants de la femelle d'avoir des descendants mâles plus à même d'être choisis par les femelles. La femelle peut aussi choisir le mâle selon des critères qui reflètent la qualité génétique de celui-ci, et donc la capacité de survie des descendants, en évaluant par exemple sa capacité à produire des organes coûteux[17]. Enfin, une femelle peut choisir le mâle susceptible de fournir les descendants les plus viables compte tenu de son propre potentiel génétique. L'hétérozygotie étant généralement considéré comme un avantage[18], la femelle choisit alors un mâle dont le patrimoine génétique est le plus différent du sien[9], par le biais du complexe majeur d'histocompatibilité par exemple. De même, les femelles peuvent exercer un choix post-copulatoire permettant de sélectionner les gamètes qui sont compatibles avec ses ovules, ce choix pouvant donc être différent d'une femelle à l'autre[19]. Une telle sélection de la compatibilité génétique permettrait aux femelles d'éviter la fécondation par du sperme incompatible, et donc l'avortement de l'embryon, à cause de conflits cytoplasmiques par exemple.
Runaway de Fisher
Indicateurs de la qualité génétique
Complémentarité génétique
Article détaillé : CMH et choix du partenaire sexuel.Compatibilité génétique
Caractère non-adaptatif
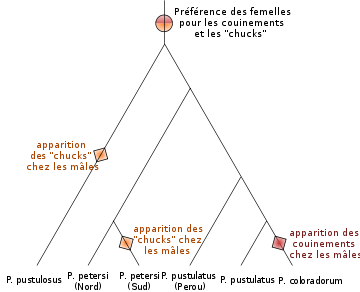 Dans le genre de crapaud Physalaemus, la préférence des femelles pour deux types de cris mâles, des couinements et des sortes de "chucks", existe même chez des espèces dont les mâles ne poussent pas ces cris. Ceci suggère que la préférence des femelles pour les cris est apparue avant les cris au cours de l'évolution, et que les cris sont apparus chez les mâles en exploitant ce biais de préférence des femelles[20].
Dans le genre de crapaud Physalaemus, la préférence des femelles pour deux types de cris mâles, des couinements et des sortes de "chucks", existe même chez des espèces dont les mâles ne poussent pas ces cris. Ceci suggère que la préférence des femelles pour les cris est apparue avant les cris au cours de l'évolution, et que les cris sont apparus chez les mâles en exploitant ce biais de préférence des femelles[20].Il est aussi possible que les femelles exercent un choix sur la paternité de leurs descendants sans que ce choix n'ait été sélectionné, et le choix n'apporte aucun bénéfice ou coût supplémentaire notable pour la femelle. La préférence de la femelle peut-être alors due à un biais dans son système sensoriel, sélectionné pour des raisons autres que la reproduction, comme la recherche de nourriture. Les mâles exploitant ce biais sensoriel sont alors préférés et sélectionnés[21]. Cette hypothèse, au point de départ non adaptative, peut-être renforcée par un phénomène de coévolution antagoniste entre les mâles et les femelles[22]. En effet, si le choix engendré par ce biais sensoriel diminue le fitness de la femelle, ce choix va être contre-sélectionné, et la sensibilité de la femelle pour ce caractère peut baisser. Les mâles dont la valeur de ce caractère dépasse ce seuil de sensibilité vont alors être sélectionnés, favorisant l'évolution d'une baisse de sensibilité encore plus importante chez les femelles.
Exploitation d'un biais sensoriel
Coévolution antagoniste entre les sexes
Coûts du choix de la paternité
 Le poids des femelles du Gobe-mouche noir est d'autant plus faible qu'elles recherchent longtemps un partenaire[23]. Ainsi, le choix du partenaire prend du temps qui ne peut être consacré à se nourrir, ce qui impose un coût énergétique à la femelle.
Le poids des femelles du Gobe-mouche noir est d'autant plus faible qu'elles recherchent longtemps un partenaire[23]. Ainsi, le choix du partenaire prend du temps qui ne peut être consacré à se nourrir, ce qui impose un coût énergétique à la femelle.Le choix des femelles a aussi un coût, qui peut ne pas être négligeable. Ce coût peut prendre différentes formes[24]. Tout d'abord, le choix lui-même est coûteux. Premièrement en termes de temps, la femelle ne pouvant se consacrer à d'autres activités telles que la recherche de nourriture ou le soin des petits pendant ce temps. Il est aussi coûteux en énergie, laquelle est nécessaire à l'échantillonnage des mâles ou des gamètes et à la prise de décision elle-même. De plus, le choix peut engendrer des coûts indirects, par exemple si la femelle est plus vulnérable aux prédateurs pendant la période de choix.
Le choix de la paternité par les femelles
Le choix d'une femelle pour la paternité de ses enfants se déroule en plusieurs étapes, et peut s'exercer soit sur le mâle, soit sur ses gamètes, soit sur les descendants eux-mêmes. Tout d'abord, la femelle procède à un échantillonnage des candidats qui seront évalués. Cet échantillonnage peut être aléatoire ou non, et son ampleur dépend des coûts et bénéfices procurés par le choix. La femelle évalue alors chacun de ces candidats, déterminant lequel peut potentiellement maximiser son fitness. Elle peut ainsi établir une fonction de préférence entre les candidats évalués. Enfin, la femelle décide du ou des candidats qui seront sélectionnés en fonction de cette préférence et d'autres critères environnementaux. Le processus de choix des femelles a cependant été majoritairement étudié pour le choix du partenaire sexuel, et on connaît mal le mécanisme de choix post-copulatoire.
Niveau du choix
Le choix d'une femelle pour le ou les mâles lui donnant des descendants peut se faire à plusieurs niveaux. La femelle choisit tout d'abord ses partenaires sexuels, autorisant les gamètes de ceux-ci à rencontrer les siens. Il peut alors exister un choix entre les gamètes d'un même mâle, ou de différents mâles si la femelle s'accouple plusieurs fois. Enfin, la femelle peut choisir ou privilégier un ou plusieurs de ses descendants par rapport aux autres.
Choix du partenaire sexuel
Le choix du partenaire sexuel, c'est à dire le fait que les femelles ne s'accouplent pas au hasard avec les mâles qu'elles disponibles, est la forme de sélection intersexuelle la plus évidente, et la mieux étudiée. Le choix du partenaire peut s'exercer à toutes les étapes de l'accouplement. La femelle peut d'abord choisir le groupe de mâles parmi lequel elle va choisir son partenaire, notamment lorsque les mâles sont regroupés en lek lors de la parade nuptiale[25]. Elle peut ensuite choisir de ne visiter que certains mâles et examiner plus précisément ceux-ci. Elle opère alors un nouveaux choix entre les mâles étudiés, et décide de celui avec lequel elle s'accouple. Enfin, la femelle peut aussi choisir d'interrompre l'accouplement et empêcher le mâle de délivrer son sperme.
Choix des gamètes mâles
Les femelles peuvent aussi effectuer un choix sur les gamètes mâles qui vont féconder leurs ovules, appelé choix femelle cryptique[26]. Ce choix peut s'effectuer dès l'accouplement, par un rejet du sperme tout juste entré. La femelle peut aussi digérer les gamètes, ou les stocker à l'écart des ovules. Si la fécondation se déroule lors du dépôt des œufs, comme chez certains insectes, le choix cryptique peut s'exercer à travers le motif de dépôt des œufs. Le choix des gamètes est aussi le principal mécanisme de sélection intersexuel connu chez les plantes, exercé sur le pollen lors de son passage dans le style.
Les avantages sélectifs du choix des gamètes, pour la femelle, sont semblables à ceux procurés par le choix du partenaire sexuel. Ainsi, l'hypothèse du beau sperme, équivalente au modèle du beau fils de Fisher, postule que la préférence pour un caractère du gamète et le caractère lui-même sont génétiquement liés, pouvant engendrer un emballement entre le caractère et la préférence. De même, l'hypothèse du bon sperme est équivalente à celle des bon gènes, et suppose que la femelle choisit les gamètes suivant des critères indiquant la qualité des gènes du gamète, garantissant une meilleure survie à l'œuf ainsi produit.
Choix des descendants
Le choix de la paternité peut enfin s'exercer après la fécondation des ovules, directement sur les œufs. Le choix peut être radical, et consister en un avortement de certains descendants. Ceci peut avoir un avantage pour la femelle. Si la femelle sait que son enfant sera tué par un autre mâle à la naissance, par exemple, il peut être avantageux pour elle de s'épargner le coût de la gestation et de l'accouchement. Le choix des descendants peut aussi s'exercer par un investissement différent dans les différents œufs produits. Pour cela, la femelle peut par exemple produire plus d'œufs après certains accouplements, nourrir plus tôt certains jeunes, ou encore accorder plus ou moins d'investissement parental aux différents enfants[27].
Échantillonnage
Les femelles ne peuvent généralement évaluer tous les candidats, et doivent en sélectionner un certain nombre qui seront évalués. Plusieurs stratégies d'échantillonnage peuvent ainsi être utilisées. Michael Jennions et Marion Petrie distinguent 6 stratégies différentes d'échantillonnage du partenaire sexuel[28]. La tactique d'accouplement aléatoire consiste à accepter le premier partenaire potentiel rencontré, ce qui revient à ne pas exercer de choix du partenaire. Lors d'une stratégie de seuil fixe, les mâles sont évalués successivement, et si la qualité de l'un deux est supérieure à un seuil fixé, il est choisi et l'échantillonnage s'arrête. Les femelles suivant une tactique de comparaison séquentielle[29] évaluent successivement les mâles et s'arrêtent dès lors que la qualité du dernier mâle est inférieure à celle du précédent et retournent s'accoupler avec ce dernier. La stratégie de décision en une étape[30] consiste à évaluer successivement les mâles et à s'arrêter et s'accoupler avec le dernier mâle lorsque celui-ci est plus avantageux que ce que la femelle peut espérer trouver par la suite, compte tenu de l'échantillonnage qu'elle a déjà effectué et du coût de l'évaluation. Elle peut ainsi être considérée comme un équivalent de la stratégie à seuil fixe, mais dont le seuil varie en fonction du coût et des bénéfices présumés issus de la poursuite de l'échantillonnage. De même, lors d'une stratégie utilisant une règle d'interruption optimale[31], les femelles s'arrêtent en fonction des qualités probables des futures mâles rencontrés, mais ne connaissent pas a priori comment est distribuée la qualité de ces mâles. Les femelles peuvent aussi effectuer une comparaison groupée[30], évaluant un nombre N défini de mâle et sélectionnant le meilleur d'entre eux.
Évaluation
L'évaluation des candidats se fonde probablement toujours sur plusieurs critères[28]. Pour chacun de ceux-ci on peut déterminer la valeur préféré par la femelle. Ces critères sont très variés, et peuvent avoir été sélectionnés de manière directe, ou indirecte en tant qu'indicateurs. Cependant ces différents critères n'ont pas tous la même importance, et il existe différentes manières d'utiliser ces critères pour classer l'ensemble des candidats selon une fonction de préférence.
Critères d'évaluation
Tout stimulus sensoriel associé au candidat peut être utilisé comme critère d'évaluation de celui-ci. On observe ainsi une grande variété de critères de choix utilisés par les femelles au sein du monde vivant. Ces critères sont souvent issus d'information visuelles, mais peuvent aussi bien être chimiques (goût, odeur) que sonores ou tactiles. Par ailleurs, ces critères peuvent correspondre à des caractéristiques physiques du mâle, comme la coloration ou la fréquence du chant, mais aussi comportementales ou écologiques (territoire occupé par le mâle, par exemple).
Un critère d'évaluation peut avoir été sélectionné directement chez la femelle, parce le choix de celui-ci lui procure un avantage sélectif. Ainsi, certaines femelles poissons s'accouplent préférentiellement avec des mâles dont elles peuvent évaluer directement l'investissement parental (lorsque les mâles gardent les œufs issus d'un autre accouplement par exemple), et donc une meilleure survie de ses descendants. Cependant, de nombreux critères sont sélectionnés par les femelles en tant qu'indicateurs. Ceux-ci peuvent aussi bien indiquer des bénéfices indirects, comme la qualité génétique du mâle, que directs, comme la santé du mâle et sa propension à transmettre des parasites à la femelle.
Utilisation de plusieurs critères
 Chez la bouvière, le succès d'un mâle auprès des femelles dépend à la fois de son comportement de parade, de sa couleur rouge, et du territoire de ponte qu'il défend. Cependant, ces critères de sélection n'interviennent pas au même moment de la phase de décision: les deux premiers influent sur le choix de la femelle d'inspecter ou non le mâle, alors que le dernier intervient dans sa décision de se reproduire avec lui, alors qu'elle l'inspecte[32].
Chez la bouvière, le succès d'un mâle auprès des femelles dépend à la fois de son comportement de parade, de sa couleur rouge, et du territoire de ponte qu'il défend. Cependant, ces critères de sélection n'interviennent pas au même moment de la phase de décision: les deux premiers influent sur le choix de la femelle d'inspecter ou non le mâle, alors que le dernier intervient dans sa décision de se reproduire avec lui, alors qu'elle l'inspecte[32].Le choix du partenaire est généralement basé sur plusieurs critères. L'usage de différents critères pour évaluer la qualité du mâle peut s'expliquer de plusieurs manières[33]. Ces différents signaux peuvent être porteurs d'informations différentes[34]. Les signaux peuvent cependant être redondants et porter le même type d'information, mais chaque signal pris séparément n'apporte pas une information assez pertinente, et il est nécessaire de combiner les signaux[34]. Les différents signaux peuvent aussi porter la même information, mais dans différentes conditions[33], par exemple à des distances différentes[28]. Par ailleurs, il est possible que, du fait de sa psychologie, le destinataire du signal soit plus sensible à des signaux à plusieurs composantes qu'à des signaux simples[35],[33]. Enfin, un certain nombre de ces critères de choix peuvent être maintenus par une légère préférence femelle peu coûteuse, sans être reliés à la qualité du mâle[34]. Ce processus peut même être amplifié par une coévolution antagoniste entre les deux sexes, poussant les mâles à exploiter les biais sensoriels des femelles et les femelles à résister à cette exploitation, multipliant le nombre de signaux utilisés[33].
Importance relative des différents critères
Le mâle préféré peut-être différent selon chaque critère considéré, et les femelles synthétisent l'information apportée par les différents critères utilisés en une fonction de préférence entre les mâles. Pour cela les femelles peuvent établir (sans que cela soit nécessairement conscient) une sorte d'index global de la qualité du mâle à partir des différents critères, en pondérant chaque critère d'une importance différente par exemple[29]. La synthèse des informations apportée par les différents signaux peut aussi se faire par des interactions entre les différents signaux, lesquelles peuvent être multiples[33]. Ainsi, l'attention portée à un signal chez un mâle peut dépendre de la valeur des autres signaux chez ce même mâle. Par ailleurs, certains signaux peuvent amplifier l'information donnée par d'autres[36]. Enfin, un signal peut modifier le coût d'un autre signal[37].
Décision
La décision correspond au choix effectif du candidat, qui se déroule effectivement. La décision est fortement influencée par la fonction de décision établie par la femelle entre les candidats. Cependant, le candidat choisi peut être différent de celui préféré[28],[38]. En effet, le choix final dépend d'autres facteurs. Le coût que peut avoir le choix d'un candidat peut être trop important, par exemple si la femelle s'est éloignée du mâle préféré au cours du processus d'échantillonnage, et qu'il lui faut le retrouver avant de pouvoir s'accoupler. Ainsi la stringence du choix, c'est à dire le prix que la femelle est prête à payer pour s'accoupler avec son mâle préféré, est un paramètre important lors de la prise de décision. Le choix effectif dépend aussi de la sélection intrasexuelle. Ainsi la compétition entre les mâles peut conduire à l'élimination du mâle préféré par une femelle, et la compétition entre femelles peut empêcher une femelle de se reproduire avec le mâle qu'elle préfère, si celui-ci est monopolisé par une autre femelle. Enfin, le choix dépend de facteur environnementaux, tels que la structure physique du milieu, qui peuvent empêcher la femelle de choisir le mâle préféré. Le milieu peut ainsi modifier la transmission du signal, fausser les informations reçues par la femelle, et donc son choix, ou encore modifier le coût associé au choix du mâle préféré.
Conséquences indirectes de la sélection intersexuelle
Dimorphisme sexuel
Compétition intrasexuelle
Conflit avec la sélection utilitaire
Spéciation
Maintien de la reproduction sexuée
Cas particuliers de sélection intersexuelle
Le paradoxe des leks
La sélection intersexuelle chez les plantes
Notes et références
- ↑ Charles Darwin (trad. Edmond Barbier, préface de Carl Vogt), La descendance de l'homme et la sélection sexuelle [« The descent of man, and selection in relation to sex »], C. Reinwald & Cie, 707 p., partie II, chap. XXI (« Conclusion principale »), p. 673.
"la lutte a également lieu entre individus de même sexe, pour séduire et attirer les femelles, généralement les femelles ne restent point passives et choisissent les mâles qui ont pour elles le plus d'attrait."
- ↑ a et b (en) Malte Andersson et Leigh W. Simmons, « Sexual selection and mate choice », dans Trends in Ecology and Evolution, vol. 21, no 6, June 2006, p. 877-904 (ISSN 0169-5347) [lien DOI]
- ↑ a et b (en) Malte Andersson, Sexual selection, Princeton University Press, 1994, 624 p. (ISBN 9780691000572, ISBN 0691000573 et ISBN 9780691033440) (OCLC 28891551), « Préface »
- ↑ a , b et c (en) Malte Andersson, Sexual selection, Princeton University Press, 1994, 624 p. (ISBN 9780691000572, ISBN 0691000573 et ISBN 9780691033440) (OCLC 28891551), chap. 1.5 (« Evolution of Female Preferences for Male Traits »)
- ↑ (en) Thomas Hunt Morgan, Evolution and Adaptation, Macmillan, New York, 1903, 470 p..
"Shall we assume that still another process of selection is going on, [...] that those females whose taste has soared a little higher than that of the average (a variation of this sort having appeared) select males to correspond, and thus the two continue heaping up the ornaments on one side and the appreciation of these ornaments on the other? No doubt an interesting fiction could be built up along these lines, but would anyone believe it, and, if he did, could he prove it?"
- ↑ a et b (en) Ronald Aylmer Fisher, « The evolution of sexual preference », dans Eugenics Review, vol. 7, 1915, p. 184-192
- ↑ (en) Ronald Aylmer Fisher et C.S. Stock, « Cuénot on preadaptation », dans Eugenics Review, vol. 7, 1915, p. 46-61
- ↑ (en) Alfred Russel Wallace, Darwinism, Macmillan, London, 1889, p. 295
- ↑ a , b et c (en) Bernard Campbell (dir.) et Robert Trivers, Sexual selection and the descent of man, 1871-1971, Chicago, Aldine, 1972 (ISBN 0202020053 et ISBN 9780202020051) (OCLC 525995), « Parental investment and sexual selection », p. 136-179
- ↑ (en) Hanna Kokko et Michael D. Jennions, « Parental investment, sexual selection and sex ratios », dans Journal of evolutionary biology, vol. 21, no 4, Juillet 2008, p. 919-948 (ISSN 1010-061X) [lien DOI]
- ↑ (en) Andreas Paul, « Sexual selection and mate choice », dans International Journal of Primatology, vol. 23, no 4, Août 2002, p. 877-904 (ISSN 0164-0291) [lien DOI]
- ↑ (en) Malte Andersson, Sexual selection, Princeton University Press, 1994, 624 p. (ISBN 9780691000572, ISBN 0691000573 et ISBN 9780691033440) (OCLC 28891551), chap. 7.2 (« Sexual Selection, Parental Investment, Sex Ratio, and Reproductive Rate »)
- ↑ (en) Stephen T. Emlen et Lewis W. Oring, « Ecology, sexual selection, and the evolution of mating systems », dans Science, vol. 197, no 4300, 15 Juillet 1977, p. 215-233 [lien DOI]
- ↑ a , b , c et d (en) Tim H. Clutton-Brock et A.C.J. Vincent, « Sexual selection and the potential reproductive rates of males and females », dans Nature, vol. 351, 1991, p. 58-60 (ISSN 0028-0836) [lien DOI]
- ↑ (en) Bernard Campbell (dir.) et Robert Trivers, Sexual selection and the descent of man, 1871-1971, Chicago, Aldine, 1972 (ISBN 0202020053 et ISBN 9780202020051) (OCLC 525995), « Parental investment and sexual selection », p. 139.
"I first define parental investment as any investment by the parent in an individual offspring that increases the offspring's chance of surviving (and hence reproductive success) at the cost of the parent's ability to invest in other offspring"
- ↑ (en) Ronald Aylmer Fisher, The Genetical Theory of Natural Selection, 1930
- ↑ (en) William Donald Hamilton et M. Zuk, « Heritable true fitness and bright birds: A rôle for parasites? », dans Science, vol. 218, 1982, p. 384-387 (ISSN 0036-8075)
- ↑ (en) J.L. Brown, « A theory of mate choice based on heterozygosity », dans Behavioral ecology, vol. 8, 1997, p. 60-65 (ISSN 10452249) [lien DOI]
- ↑ (en) J. A. Zeh et D. W. Zeh, « The evolution of polyandry I: Intragenomic conflict and genetic incompatibility », dans Proceedings of the Royal Society B, vol. 263, no 1377, 1996, p. 1711-1717 (ISSN 0962-8452)
- ↑ (en) Kerry Shaw, « Phylogenetic tests of the sensory exploitation model of sexual selection », dans TREE, vol. 10, no 3, Mars 1995, p. 117-120 (ISSN 0169-5347) [lien DOI]
- ↑ (en) M.J. Ryan, « Sexual selection, sensory systems and sensory exploitation », dans Oxford Surveys in Evolutionary Biology, vol. 7, 1990, p. 157–195 (ISSN 0265-072X)
- ↑ (en) B. Holland et W.R Rice, « Chase-away selection: Antagonistic seduction versus resistance », dans Evolution, vol. 52, no 1, 1998, p. 1-7 (ISSN 00143820)
- ↑ (en) Tore Slagsvold, Jan T. Lifjeld, Geir Stenmark et Torgrim Breiehagen, « On the cost of searching for a mate in female pied flycatchers Ficedula hypoleuca », dans Animal Behavior, vol. 36, Avril 1988, p. 433-442 (ISSN 0003-3472) [lien DOI]
- ↑ (en) John D. Reynolds et Mart R. Gross, « Costs and benefits of female mate choice: is there a lek paradox? », dans The American Naturalist, vol. 136, no 2, Août 1990, p. 230-243 (ISSN 0003-0147)
- ↑ (en) Jacob Höglund et Rauno V. Alatalo, Leks, 1995, 224 p. (ISBN 978-0-691-03727-1) (OCLC 31374226)
- ↑ « », dans
- ↑ (en) E.J.A. Cunningham et T.R. Birkhead, « Sex roles and sexual selection », dans Animal Behaviour, vol. 56, Décembre 1998, p. 1311-1321 (ISSN 0003-3472)
- ↑ a , b , c et d (en) Michael D. Jennions et Marion Petrie, « Variation in mate choice and mating preferences: a review of causes and consequences », dans Biological Reviews of the Cambridge Philosophical Society, vol. 72, no 2, 1997, p. 283-327 (ISSN 0006-3231) [lien DOI]
- ↑ a et b (en) Patrick Bateson (dir.) et James F. Wittenberger, Mate choice, Cambridge University Press, 1983 (ISBN 9780521272070, ISBN 0521272076 et ISBN 9780521251129) (OCLC 8709418), « Tactics of mate choice », p. 435-447
- ↑ a et b (en) A.C. Janetos, « Strategies of female mate choice: a theoretical analysis », dans Behavioral Ecology and Sociobiology, vol. 7, no 2, 1980, p. 107-112 (ISSN 0340-5443)
- ↑ (en) Y. Dombrovsky et N. Perrin, « On adaptive search and optimal stopping in sequential mate choice », dans American Naturalist, vol. 144, no 2, Août 1994, p. 355-361 (ISSN 0003-0147) [lien DOI]
- ↑ (en) Ulrika Candolin et J.D. Reynolds, « Sexual signaling in the European bitterling: females learn the truth by direct inspection of the resource », dans Behavioral Ecology, vol. 12, no 4, 2001, p. 407-411 (ISSN 1045-2249) [texte intégral]
- ↑ a , b , c , d et e (en) Ulrika Candolin, « The use of multiple cues in mate choice », dans Biological Reviews, vol. 78, no 4, 2003, p. 575-595 (ISSN 1464-7931) [texte intégral lien DOI]
- ↑ a , b et c (en) A.P. Moller et A. Pomiankowsky, « Why have birds got multiple sexual ornaments ? », dans Behavioral Ecology and Sociobiology, vol. 32, no 3, 1993, p. 167-176 (ISSN 0340-5443) [lien DOI]
- ↑ (en) T. Guilford et Dawkins, « Receiver psychology and the evolution of animal signals », dans Animal Behavior, vol. 42, Juillet 1991, p. 1-14 (ISSN 0003-3472) [lien DOI]
- ↑ (en) Oren Hasson, « Amplifiers and the handicap principle in sexual selection: a different emphasis », dans Proceedings of the Royal Society B, vol. 235, no 1281, 23 Janvier 1989, p. 383-406 (ISSN 0962-8452)
- ↑ (en) Rufus A. Johnstone, « Multiple displays in animal communication: 'Backup signals' and 'multiple messages' », dans Philosophical Transactions of the Royal Society B, vol. 351, no 1337, 29 Mars 1996, p. 329-338 (ISSN 0962-8436) [lien DOI]
- ↑ (en) Samuel Cotton, Jennifer Small et Andrew Pomiankowski, « Sexual selection and condition-dependent mate preferences », dans [[Current Biology|Current Biology]], vol. 16, no 17, 5 Septembre 2006, p. R755-R765 (ISSN 0960-9822) [lien DOI]
- ↑ (en) Lisa Horth, « Sensory genes and mate choice: Evidence that duplications, mutations, and adaptive evolution alter variation in mating cue genes and their receptors », dans Genomics, vol. 90, no 2, 2007, p. 159-175 (ISSN 0888-7543) [lien DOI]
- ↑ (en) A.F. Agrawal, « Sexual selection and the maintenance of sexual reproduction », dans Nature, vol. 411, 2001, p. 692-695 (ISSN 0028-0836) [lien DOI]
Voir aussi
Articles connexes
- Évolution
- Sélection sexuelle
- Sélection intrasexuelle
- Course aux armements
- Théorie de la reine rouge
- Runaway de Fisher
Liens externes
Bibliographie
- (en) Malte Andersson, Sexual selection, Princeton University Press, 1994, 624 p. (ISBN 9780691000572, ISBN 0691000573 et ISBN 9780691033440) (OCLC 28891551)
- (en) Patrick Bateson (dir.), T. R. Halliday, Wolfgang Wickler, Uta Seibt, Peter O’Donald, Stevan J. Arnold, Jack W. Bradbury, Robert M. Gibson, G. A. Parker et al., Mate choice, Cambridge University Press, 1983, 480 p. (ISBN 9780521272070, ISBN 0521272076 et ISBN 9780521251129) (OCLC 8709418)
- (en) Andreas Paul, « Sexual selection and mate choice », dans International Journal of Primatology, vol. 23, no 4, Août 2002, p. 877-904 (ISSN 0164-0291)
- (en) Jacob Höglund et Rauno V. Alatalo, Leks, 1995, 224 p. (ISBN 978-0-691-03727-1) (OCLC 31374226)
Catégorie : Sélection sexuelle - ↑ Charles Darwin (trad. Edmond Barbier, préface de Carl Vogt), La descendance de l'homme et la sélection sexuelle [« The descent of man, and selection in relation to sex »], C. Reinwald & Cie, 707 p., partie II, chap. XXI (« Conclusion principale »), p. 673.





Wikimedia Foundation. 2010.