- Vol de l'insecte
-
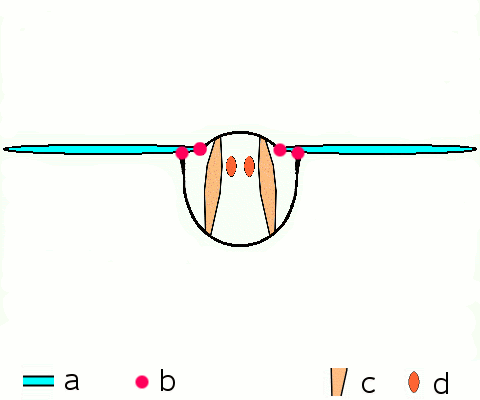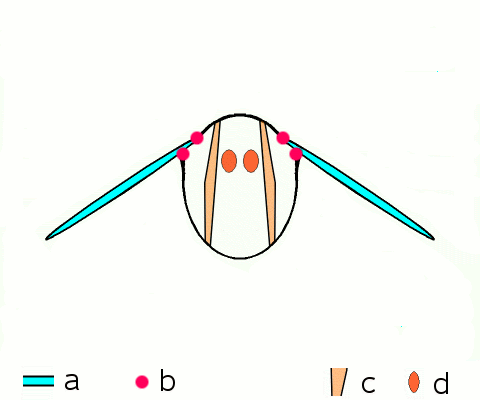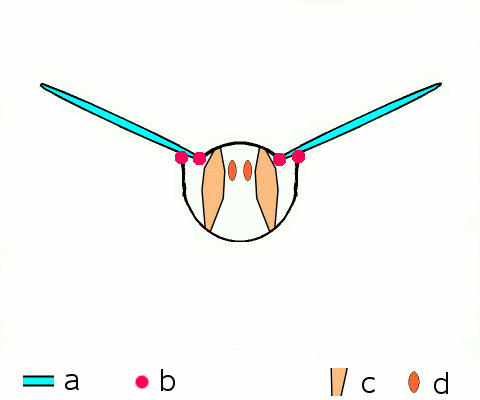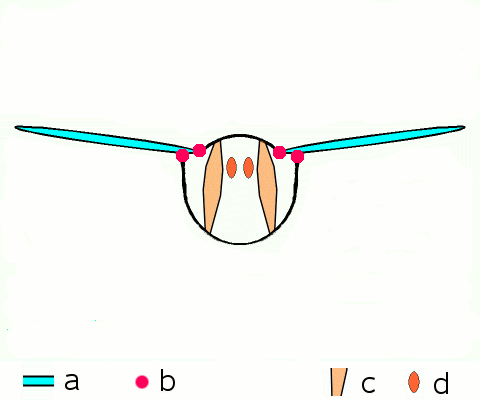
 Vol d'approche d'un argus bleu (Polyommatus icarus)
Vol d'approche d'un argus bleu (Polyommatus icarus)
 Vol d'un syrphe
Vol d'un syrpheLes insectes sont les seuls parmi les invertébrés à pouvoir réellement voler. Les araignées, ainsi que de nombreux autres petits organismes, peuvent se laisser emporter par le vent, mais ils n'ont pas d'ailes et ne peuvent pas diriger leur mouvement. La possibilité de voler a été importante pour la dispersion des insectes. Cette faculté leur permet d'échapper à leurs prédateurs, de s'accoupler plus facilement, d'atteindre de nouveaux biotopes et de nouvelles réserves alimentaires où il pourront déposer leur progéniture.
Seuls les insectes au stade final d'imago (ou accessoirement de subimago chez les éphémères) sont capables de voler. Aucune larve d'insecte ne possède cette faculté.De même, tous les insectes, même s'ils sont ailés ne volent pas forcément. Certains répugnent simplement à prendre l'envol alors que d'autres en sont incapables.
Au cours des 350 derniers millions d'années, les insectes ont développé des aptitudes au vol remarquables et techniquement originales, supérieures en bien des points à celles inventées par les hommes. Même les connaissances les plus récentes en aérodynamique, sur le vol battu ou sur les ailes flexibles, ne sont pas encore en mesure d'expliquer toutes les performances des insectes.L'étude du vol des insectes a des applications technologiques, comme celle des micro-drones.
Sommaire
Évolution et adaptation
Il y a 350 millions d'années, à l'époque du Carbonifère, alors que la Terre ne comptait que deux supercontinents, les insectes se sont à un moment mis à voler. Les tout premiers “aviateurs” de l'histoire de la Terre ressemblaient aux libellules, possédaient deux paires d'ailes auxquelles les muscles du vol étaient directement rattachés, et étaient incapables de replier leur ailes sur leur abdomen.
La plupart des insectes actuels, qui descendent de ces premiers insectes volants, ont simplifié leur anatomie en réduisant à une le nombre de paires d'ailes ou en faisant fonctionner leur deux paires comme une seule, et en utilisant un système d'attachement indirect des muscles du vol. La sélection naturelle a affiné les ailes, leur système de contrôle et leur système sensoriel, et tout ce qui affecte l'aérodynamique ou la cinématique.
Un trait particulier de l'aile de l'insecte est sa torsion (son "calage"). La plupart des ailes d'insectes présentent une torsion, comme les pales des hélices, avec un angle d'attaque plus grand à la base qu'à l'extrémité. Cette torsion est de l'ordre de 10 à 20 degrés. De plus, les surfaces alaires ne sont pas nécessairement plates ou lisses : la plupart des plus gros insectes ont la membrane des ailes distordue avec des angles entre les nervures disposés de telle sorte que la section en coupe de l'aile est approximativement profilée comme celle d'un avion. Ainsi, la forme de base de l'aile est déjà capable de générer une certaine portance même à l'état stationnaire.
La plupart des insectes contrôlent leur ailes en ajustant l'angle d'attaque, la rigidité, et la fréquence de battement à l'aide de petits muscles situés dans le thorax.
Certains insectes ont développé des caractéristiques alaires désavantageuses pour le vol mais pouvant servir à d'autre fonctions, par exemple pour l'accouplement ou pour la protection.
D'autres insectes, en occupant des niches écologiques spécialisées, ont eu besoin d'acquérir des capacités incroyables à manœuvrer, pour accéder à la nourriture dans des espaces très exigus, pour échapper à des prédateurs plus grands qu'eux, ou au contraire, en tant que prédateurs, pour capturer leurs proies. La manœuvrabilité, d'un point de vue aérodynamique, est permise par d'importantes forces de portance et de poussée : les insectes volants produisent généralement des forces de portance atteignant jusqu'à trois fois leur poids et des forces de poussée horizontales atteignant jusqu'à cinq fois leur poids.
Il existe deux grands types différents de mécanismes de vol des insectes. Chaque type a ses propres avantages et désavantages. Le fait que les odonates aient un mécanisme de vol plus primitif ne fait pas d'eux de mauvais voiliers ; ils sont même, dans une certaine mesure, plus agiles que n'importe quoi qui ait évolué par la suite.
Mécanisme de vol direct
 Aeschne des joncs (Aeshna juncea) en vol
Aeschne des joncs (Aeshna juncea) en volChez les paléoptères, ordre de classification traditionnelle qui comprend les éphémères et les odonates, les muscles alaires, au contraire de la plupart des autres insectes, s'insèrent directement à la base des ailes. L'articulation est faite de telle façon qu'un petit mouvement vers le bas de la base de l'aile soulève le reste de l'aile vers le haut, à la manière d'une rame que l'on remonte en l'air.
Les ailes postérieures des éphémères sont réduites ou même quelquefois absentes et jouent un rôle mineur dans le vol, malhabile et disgracieux.
En revanche, chez les odonates (les demoiselles et les libellules), la paire d'ailes antérieures et la paire d'ailes postérieures sont semblables par la forme et par la taille. Elles opèrent indépendamment, ce qui permet un très fin degré de contrôle de la mobilité et une capacité, inégalée par les autres insectes volants, à pouvoir changer soudainement de direction et de vitesse, notamment pour chasser. La supériorité technique de ces prédateurs sur leurs proies pourrait expliquer la longévité de ce groupe biologique qui existait déjà bien avant l'apparition des dinosaures.
De plus les demoiselles peuvent tenir un vol stationnaire. Les libellules, avec des mécanismes de vol plus primitifs, n'y parviennent qu'imparfaitement et à condition de s'aider des courants d'air.
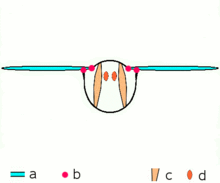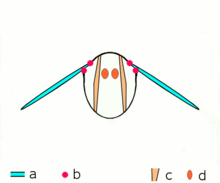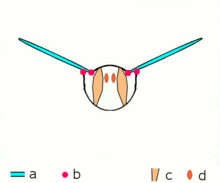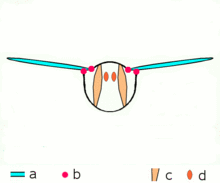 Mécanisme du vol
Mécanisme du volMécanisme de vol indirect
 Moro sphinx (Macroglossum stellatarum) en vol stationnaire
Moro sphinx (Macroglossum stellatarum) en vol stationnaireLe mécanisme de vol indirect n'est manifestement apparu qu'une seule fois au cours de l'évolution, chez l'ancêtre de tous les autres insectes volants. C'est le caractère commun, ou synapomorphique, qui permet de définir l'infraclasse des néoptères : les muscles responsables du vol ne sont pas directement attachés aux ailes.
Ce caractère est associé au mécanisme de repliement des ailes sur le dessus de l'abdomen lors du repos, mais l'apparition des deux mécanismes n'a peut-être pas été simultanée. Actuellement, certains groupes, en particulier tous les papillons, ont perdu secondairement cette capacité à se couvrir la surface du dos avec leurs ailes.
Chez les néoptères les plus primitifs, les deux paires d'ailes peuvent encore être indépendantes, mais chez les néoptères les plus évolués et possédant deux paires d'ailes fonctionnelles, celles-ci sont liées mécaniquement de diverses façons possibles, et fonctionnent de manière synchrone comme une seule paire.
En revanche tous les néoptères partagent le même principe de transmission du mouvement aux ailes. Les muscles moteurs du vol, sont situés dans le thorax, y sont attachés et le déforment, et ce sont les déformations du thorax qui provoquent le mouvement des extensions de l'exosquelette thoracique que sont les ailes.
Une série de muscles dorsaux longitudinaux compriment le thorax de l'avant vers l'arrière, provoquant une courbure vers le haut de la surface dorsale du thorax, le notum), et faisant s'abaisser les ailes. Une série de muscles tergosternaux tirent le notum vers le bas de nouveau, provoquant le mouvement vers le haut des ailes. Dans quelques groupes, le battement vers le bas est accompli seulement par le recul élastique du thorax quand les muscles tergosternaux sont relâchés.
Plusieurs petites sclérites situées à la base de l'aile et attachées à d'autres séries de muscles, permettent d'ajuster finement la courbure et l'amplitude des battements d'ailes.
Un des ultimes raffinements apparu chez certains des néoptères supérieurs, tels les coléoptères, les diptères, et les hyménoptères, est un système de contrôle musculo-nerveux où une seule impulsion nerveuse entraîne la contraction répétée d'une fibre musculaire. Ceci permet à la fréquence des battements d'ailes d'excéder le rythme auquel le système nerveux peut envoyer des impulsions. Ce type de muscle est appelée « muscle de vol asynchrone ».
Le mécanisme de vol indirect, et ses diverses évolutions, a doté de nombreux néoptères supérieurs de capacités aériennes sophistiquées : vol stationnaire, à reculons, etc. avec une finesse et un degré de contrôle auquel les insectes ayant des muscles directement attachés aux ailes ne pouvaient prétendre.
Chez les diptères comme chez les coléoptères, le vol s'effectue à l'aide d'une seule paire d'aile. Chez les coléoptères, il s'agit de la seconde paire, la première étant transformée en élytres. Chez les diptères, c'est la première paire qui sert au vol, la seconde s'étant transformée en « haltères », qui sont de minuscules petites massues dont le mouvement sert à la mouche de gyroscope pour stabiliser son vol.
Bases de l'aérodynamique
 Abeille (Apis mellifera) dans les airs
Abeille (Apis mellifera) dans les airsIl existe deux modèles aérodynamiques de base du vol des insectes.
La plupart des insectes utilisent une méthode qui crée un vortex qui les entraîne. Ces ailes battantes se déplacent par deux demi-battements basiques. Le battement vers le bas commence en haut et vers l'arrière puis plonge vers le bas et l'avant. Ensuite l'aile est rapidement renversée (supination), afin que le bord d'appel soit pointé vers le bas. Le mouvement de battement vers le haut pousse alors l'aile vers le haut et l'arrière. Puis l'aile est retournée de nouveau (pronation), et un autre battement vers le bas peut se produire. L'échelle de la fréquence de battement des insectes ayant des muscles de vol synchrones est typiquement de l'ordre de 5 à 200 Hz. Chez les insectes pourvus de muscles de vol asynchrones, cette fréquence peut excéder 1 000 Hz. Quand l'insecte vole sur place, les deux battements sont de même durée. Un battement vers le bas plus lent, cependant, fournit de la poussée.
L'identification des forces majeures est critique pour comprendre le vol des insectes. Les premières tentatives pour comprendre le vol battu assumaient un état quasi-stable. Ceci signifie que le flux d'air au-dessus de l'aile à un moment donné était assumé être le même comme il le serait au-dessus d'une aile non battante stable au même angle d'attaque. En divisant l'aile battante en un nombre important de positions immobiles et en analysant alors chaque position, il serait possible de créer un aperçu des forces instantanées sur l'aile à chaque point de temps. La portance calculée était trop petite par un facteur de trois, et les chercheurs se dirent qu'il devait y avoir des phénomènes instables fournissant des forces aérodynamiques. Il y eut plusieurs modèles analytiques développés tentant d'approximer le flux à proximité d'une aile battante. Certains chercheurs prédirent des pics de force à la supination. Avec un modèle dynamiquement mis à l'échelle d'une mouche du vinaigre, ces forces prédites ont été confirmées plus tard. D'autres ont protesté que le pic de force durant la supination et la pronation étaient dus à un effet rotationel inconnu fondamentalement différent du phénomène de translation. Il y a des désaccords avec cet argument. Par le biais de la modélisation informatique de la dynamique des fluides, certains chercheurs affirment qu'il n'y a aucun effet rotationnel. Ils affirment que les grandes forces sont causées par une interaction avec le tourbillon dû au battement précédent.
De même que pour l'effet rotationel précédemment mentionné, les phénomènes associés aux ailes battantes ne sont pas complètement compris ou n'aboutissent pas sur un accord. Du fait que tout modèle est une approximation, différents modèles négligent des effets considérés comme négligeables. Par exemple, l'effet Wagner affirme que la circulation s'élève lentement jusqu'à son état stable à cause de la viscosité quand une aile inclinée est accélérée depuis un état de repos. Ce phénomène expliquerait une valeur de portance plus petite que prévue. Typiquement, le cas a été de trouver des sources pour la portance ajoutée. Il a été dit que cet effet est négligeable pour un flux avec un nombre de Reynolds de l’ordre de grandeur de celui du vol des insectes.
Un des plus importants phénomènes qui se produit durant le vol de l'insecte est l'effet de succion du bord d'attaque. Cette force est significative pour le calcul de l'efficacité. Le concept de succion du bord d'attaque a d'abord été amené pour décrire la portance vorticielle des ailes delta à angles aigu.
La deuxième méthode de vol, le “lancé et claqué”, fonctionne différemment. Les ailes sont lancées et claquées l'une contre l'autre au-dessus du corps de l'insecte puis rejetées chacune de son côté.Recherche actuelle
Les scientifiques étudient le vol des insectes pour diverses raisons : compréhension du développement biologique des animaux, un intérêt purement scientifique pour l'aérodynamique instable, ou l'intérêt pour la conception de micro-véhicules aériens ou appareils similaires (drones).
Voir aussi
Références
- Chapman, R.F., “The Insects : Structure and Function, 4th Edition, Cambridge University Press, 1998.
- Dickinson, M.H., Lehmann, F.-O., and Sane, S.P., “Wing Rotation and the Aerodynamic Basis of Insect Flight,” Science Magazine Vol. 284, June 1999.
- Ellington, C.P., van der Berg, C., Willmott, A.P. and Thomas, A.L.R., “Leading Edge Vortices in Insect Flight,” Nature, Vol. 384, p. 626-630, 1996.
- Ellington, C.P., “The aerodynamics of hovering insect flight,” Philosophical Transactions Royal Society of London B305, 1984.
- Haj-Hariri, H., “Unsteady Aerodynamics of Flapping Wings,” University of Virginia, 2001.
- Lewin, G.C., Haj-Hariri, H., “Modeling thrust generation of a two-dimensional heaving airfoil in a viscous flow,” Journal of Fluid Mechanics, Vol. 492, 2003.
- Lighthill, M.J., “On the Weis-Fogh mechanism of lift generation,” Journal of Fluid Mechanics, Vol. 60, p 1-17, 1973.
- Nachtigall, W., “Insects in Flight: A Glimpse Behind the Scenes in Biophysical Research.,” McGraw-Hill Book Co., NY, June 1974.
- Platzer, Max F., Department Chairman, “Aerodynamics and Aeroelasticity: Flapping-Wing Propulsion.” Last revised 2005, Retrieved 1 November 2005, from http://www.aa.nps.navy.mil/programs/aero/propulsion/
- Polhamus, E.C., “A Concept of the Vortex Lift of Sharp-Edge Delta Wings Based on a Leading-Edge-Suction Analogy,” Langley Research Center, 1966.
- Pringle, J.W.S., “Insect flight,” Oxford Biology Readers, Vol. 52, 1975.
- Sane, S.P., “The aerodynamics of insect flight,” The Journal of Experimental Biology, Vol. 206, p. 4191-4208, August 2003.
- Savage, S.B., Newman, B.G. and Wong, D.T.M., “The role of vortices and unsteady effects during the hovering flight of dragon flies,” The Journal of Experimental Biology, Vol. 83, p. 59-77, 1979.
- Smyth, T., Jr. 1985. Muscle systems, pp. 227-286. In M.S. Blum [ed.], Fundamentals of insect physiology. John Wiley and Sons, New York.
- Sotavalta, O. 1953. Recordings of high wing-stroke and thoracic vibration frequency in some midges. Biol. Bull. Woods Hole 104: 439-444.
- Van den Berg, C., Ellington, C.P., “The vortex wake of a hovering model hawk moth,” Philosophical Transactions Royal Society of London, Vol. 352, p. 317-328, 1997.
- Walker, J.A., “Rotational lift: something difference or more of the same?,” The Journal of Experimental Biology, Vol. 205, p. 3783-3792, September 2002.
- Walker, P.B., “Growth of circulation about a wing and an apparatus for measuring fluid motion,” Reports and Mem., Aeronaut. Res. Com. no. 1402, 1931.
Lien externe
Catégories :- Vol (aérodynamique)
- Biophysique de l'insecte





Wikimedia Foundation. 2010.